デジタル大辞泉 「セルロース」の意味・読み・例文・類語
セルロース(cellulose)
Sponserd by ![]()
翻訳|cellulose
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
繊維素ともいう。セルロースは自然界で最大量の有機化合物であり,おそらく世界中の全植物質の1/3を占めるだろう。セルロースの命名は,1838年フランスのパヤンAnselme Payen(1795-1871)によって,高等植物の細胞(セル)壁を構造する糖の意味でなされた。セルロースは古来衣類に用いられてきたが,工業的に利用しようという試みは近代工業とともに興った。44年,マーセルJohn Mercer(1791-1866)はセルロースとアルカリの反応(マーセリゼーション)を研究し,工業的利用への道を開いた。銅アンモニア溶液にセルロースが溶解することを,シュワイツァーE.Schweizerは57年に見いだした。クロスCharles Frederick Cross(1855-1935)とベバンEdward John Bevan(1856-1921)は93年キサントゲン酸セルロース法によるビスコースレーヨンを発明し,今日も大量生産されている。
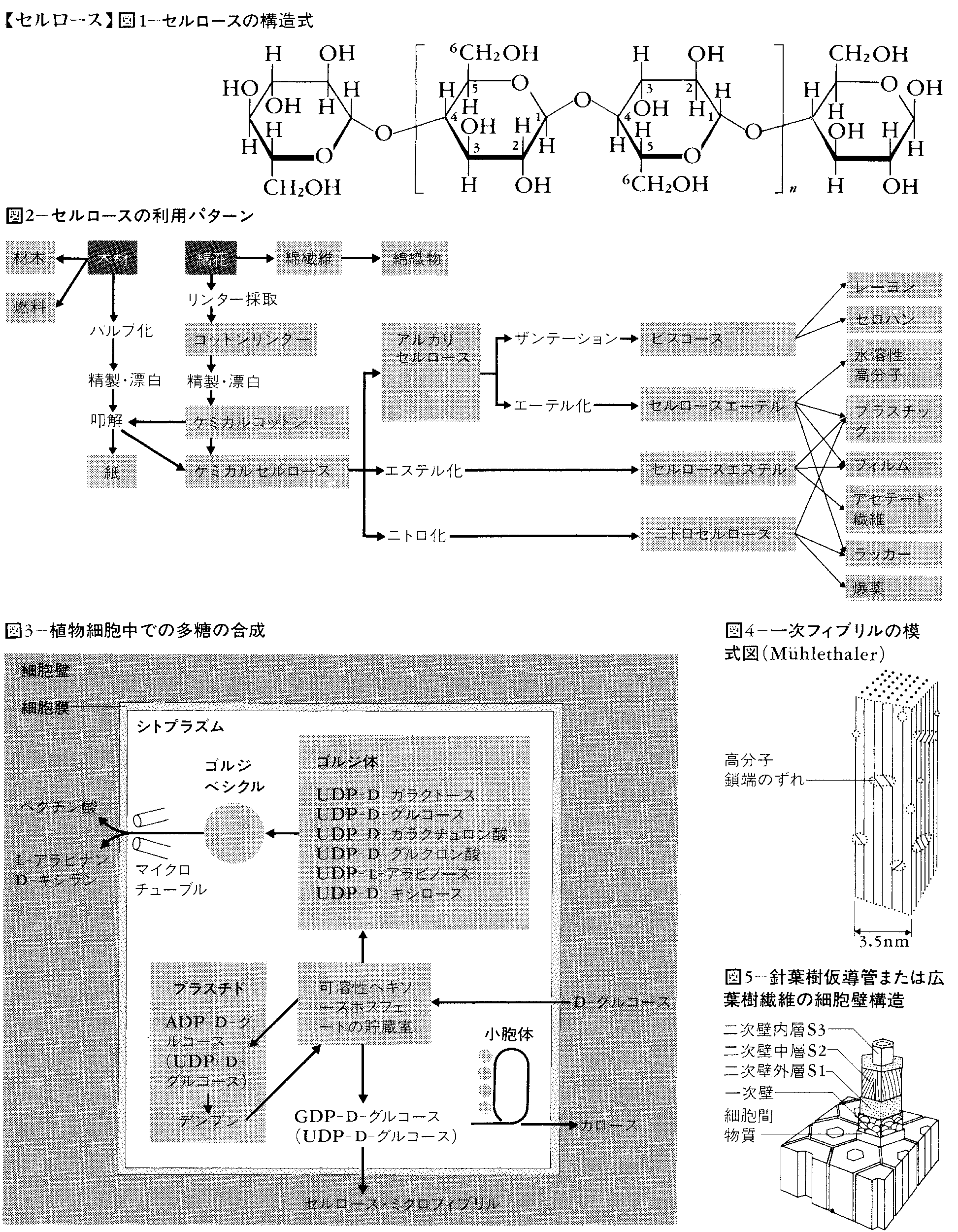
化学的意味では,セルロースは水や酸,アルカリに不溶で,図1に示すように,1位の炭素と4位の炭素によってβ-結合された無水グルコース単位から成り,特有のX線回折図を与える十分な分子量の多糖(糖のポリマー)である。純粋なセルロースはα-セルロースと呼ばれ,β-セルロースとγ-セルロースとは区別される。α-セルロースは17.5%水酸化ナトリウムに溶解後希釈して8%水酸化ナトリウムにすると不溶化する。このときは溶解しており,少し酸を加えると沈殿するのがβ-セルロースである。したがって,β-セルロースはα-セルロースと同成分だが,重合度(n)の小さいものである。γ-セルロースは,β-セルロースの沈殿後も溶解している多糖であり,キシラン(キシロースの重合体),マンナン,アラビナン,ウロン酸(カルボキシル基をもつ多糖)などからできている。β-セルロースとγ-セルロースを合わせてヘミセルロースと呼ぶ。ホロセルロースは,木材やわらなどセルロースを含む物質から,炭水化物以外の物質を除いた残りをいい,セルロースとヘミセルロースを合わせたものである。再生セルロースは,セルロースを一度溶解後,元のポリアンヒドログルコース構造へ戻したものをいう。
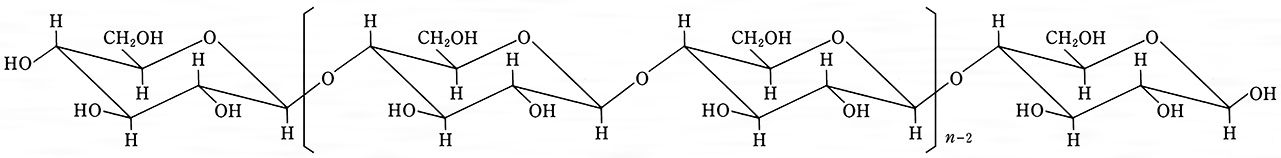
セルロースを含む原料とその含有割合を表に示す。(1)種子の毛 ワタの種子の長い繊維である綿花は94%という高い割合のセルロースを含む。また,綿花を採った後の短い繊維であるコットンリンターも同じ高い割合でセルロースを含むので,銅アンモニアレーヨン,アセチルセルロースなどセルロース誘導体の原料となる。パンヤ繊維(カポック)は詰物に広く使用される。(2)靱皮(じんぴ)繊維 木の皮を利用する麻のセルロース含量も65~75%と高い。(3)木材 すべてのセルロース原料のなかでいちばん大量にまた広く使われている。木材の40~50%はセルロースで,20~30%はリグニン,そして10~30%がセルロース以外のヘミセルロースと多糖類からできているので,パルプ化工程によってセルロースを取り出す。(4)葉繊維 マニラアサの葉茎から得られるマニラ麻およびサイザルアサの葉からのサイザル麻は,堅い繊維であるので紐に使われる。木材および綿花を出発原料とするセルロースの利用パターンを図2に示す。セルロースからその誘導体へ導かれるとき,精製されたケミカルセルロースがまず作られる。これと化学薬品を用いてザンテーション(キサントゲン酸化),エーテル化,エステル化,およびニトロ化を行い,それぞれ誘導体へ導く。ビスコース,セルロースエステル,セルロースエーテル,およびニトロセルロースを成形または塗布して,繊維,フィルム,成形品,塗料および爆薬を製造する。
セルロースは単糖のグルコースが多数結合してできた生体高分子である。セルロースの生合成は,ウリジン5′-(α-D-グルコピラノシルピロホスフェート)(UDP-D-グルコース)とグアニン5′-(α-D-グルコピラノシルピロホスフェート)(GDP-α-グルコース)という二つのヌクレオチドに,特異的トランスフェラーゼが作用して,つぎつぎに高分子鎖末端にD-グルコースを結合させて分子鎖を長くする。図3に模式的に細胞内の多糖合成の仕組みを示す。細胞は膜中にD-グルコース(ブドウ糖)を取り込み,セルロースを合成し,排出する。同時にヘミセルロースも合成されるので,セルロースと混在することになる。酢酸菌の1種Acetobacter xylinumはセルロースを生合成するが,その生成状態の観察はセルロース生合成に関する多くの知見を与える。セルロース分子の集合体が,バクテリアの細胞壁を取り囲むゲル状物質の中から30秒の誘導期間後に現れる。34℃では,ミクロフィブリル(微細繊維)と呼ばれる紐は毎分約0.2μmの速度で生長していく。このとき,2種類の酵素が働いて,生長しているセルロースの両末端にグルコース単位を続々と結合させ,さらに,このようなセルロース分子が集合して一次フィブリルが形成される。微小繊維の最小単位である一次フィブリルは,図4に示されるような3.5nmの幅をもつことが,X線回折や電子顕微鏡で調べた結果わかった(1969)。この中に36個のセルロース鎖が配列して含まれる。一次フィブリルがさらに集まって,ミクロフィブリルを作っているとする説が有力である。
木材の細胞壁は,図5のように,セルロース・ミクロフィブリルがリグニンとヘミセルロースから成る組織の中に埋め込まれた層が4層重なってできている(1959)。すなわち一次壁,二次壁外層S1(厚さ0.1~0.3μm),二次壁中層S2(1~5μm),二次壁内層S3(0.1μm)である。ミクロフィブリル内において,セルロース分子は完全に配列した結晶でなく,結晶と非結晶の混じった状態からできているという総(ふさ)状ミセル説によって,セルロース分子の配列構造が説明されている。ポリエチレンのような合成高分子の場合に認められている折畳み構造で,ミクロフィブリル中のセルロース配列を説明する説もあるが,両者ともミクロフィブリルの観測された性質を完全には説明していない。
セルロースの水酸基-OHは水やジメチルスルホキシドのような極性の大きい溶媒に強い親和性をもっている。乾燥セルロースは空気中の湿度に応じて湿気を吸収し膨潤する。吸湿してもセルロースのX線回折図は変化しないので,水分の吸収は結晶領域でなく非結晶領域で起こる。すなわち,結晶間膨潤を引き起こすわけである。吸収された水はセルロース-セルロースの水素結合を切ってセルロース-水の水素結合を作る。セルロースを溶かす溶媒があまりないのは,セルロース間の強い水素結合を切るのが難しいからである。しかし,セルロースを溶解させてレーヨンを作ろうとする研究は19世紀終りころから20世紀初頭活発になされ,セルロースの溶媒が見つかった。鉄,コバルト,ニッケル,銅,カドミウムや亜鉛のような重金属のアンモニウムやアミン錯体はセルロースを溶解する。テトラアミノ銅ジヒドロキシドCu(NH3)4(OH)2(キュプラアンモニウム)やビス(エチレンジアミン)銅ジヒドロキシド(キュエン)がそれで,前者は発見者にちなんでシュワイツァー試薬と呼ばれ,銅アンモニアレーヨンを工業的に作る溶媒である。また,これらの溶媒を用いてセルロースの粘度が測定され,分子量が求められた。1970年代に,セルロースの新しい溶媒として,ジメチルスルホキシド-パラホルムアルデヒド混合系が見つけられている。
セルロース製品の製造業者や使用者にとって,セルロースの分解は望ましくない。しかし,次のような条件下では分解は起こる。(1)微生物的分解,(2)酸加水分解,(3)酸化的分解,(4)アルカリ加水分解,および(5)機械的分解である。微生物的分解は,生物によって作られたものは生物的に分解される。セルラーゼは有名なセルロース分解酵素である(後述)。酸加水分解は,ポリマー主鎖を構成する1位の炭素と酸素間のグリコシド結合は酸によって容易に切れて分解する。セルロースを濃硫酸または濃塩酸で最初に処理し,熱い希酸で加水分解してグルコースを工業的に製造するプロセスがあるが,あまり使われていない。セルロースは酸化を受けやすく,なかでも還元末端と水酸基は最も酸化されやすい位置であり,酸化によってカルボニル基やカルボキシル基が導入される。次亜ヨウ素酸塩のような温和な酸化剤では,還元末端が酸化され,カルボキシル基になる。過ヨウ素酸や酸性の四酢酸鉛は,2位の炭素と3位の炭素間の炭素-炭素結合を切断し,ともにアルデヒド基に変え,ジアルデヒドセルロースを生ずる。アルカリ分解には空気との接触による酸化的分解があり,これは比較的高温で起こる。また,セルロースはアルカリには比較的抵抗性をもつが,160~180℃という高温ではアルカリによってグリコシル結合が切れて,分子鎖が短くなる。セルロースの熱分解には,低温で加熱によって徐々に起こる分解と高温でレボグルコサン(1,6-無水グルコース)の生成を伴って起こる分解とがある。
セルロースは分子内に多くのアルコール性水酸基をもつので,エステル化やエーテル化反応を起こし,多くの誘導体を生ずる。最も重要なエステル化は,セルロース(cellulose-OHと書く)のザンテーションである。キサントゲン酸セルロースは酸によって元のセルロースに戻り,レーヨンが得られる。
セルロースの無機エステルであるニトロセルロースは硝酸HNO3によるニトロ化で作られ,ラッカー,爆薬,推進薬などに使われる。 酢酸セルロースはアセテート繊維,プラスチック,フィルム,塗装に多く使われるセルロースのエステルである。セルロースエーテルはその水溶性とゲル生成性を生かして,粘稠剤,分散剤,膨張剤,接着剤などに使われる。よく使われるカルボキシメチルセルロース(CMC)は,アルカリセルロースにクロロ酢酸ナトリウムを反応させて作られる。
酢酸セルロースはアセテート繊維,プラスチック,フィルム,塗装に多く使われるセルロースのエステルである。セルロースエーテルはその水溶性とゲル生成性を生かして,粘稠剤,分散剤,膨張剤,接着剤などに使われる。よく使われるカルボキシメチルセルロース(CMC)は,アルカリセルロースにクロロ酢酸ナトリウムを反応させて作られる。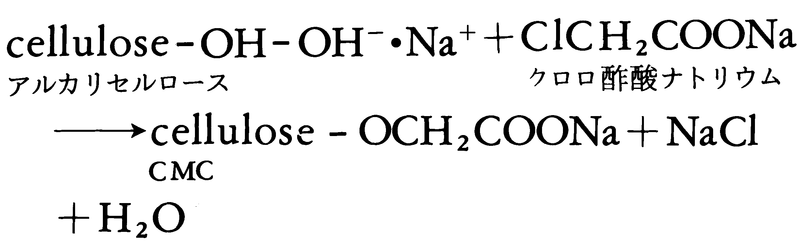
このほか,繊維の寸法安定性や防しわ性向上のため,セルロース間を結合させる架橋や合成高分子の枝を生やすグラフト化が行われている。
執筆者:瓜生 敏之
セルロースのβ,1-4結合を加水分解する酵素であるセルラーゼは,原生動物やカタツムリ,フナクイムシの消化液中といったまれな例外を除いて,一般に動物には見いだされない。また,α,1-4結合に作用するアミラーゼも,セルロースにはまったく働かない。したがって,通常の動物はセルロースを栄養素とすることができない。草食性の動物のうち,ヒツジ,ヤギ,ウシ,ラクダ,キリンなどの反芻(はんすう)動物の胃ではセルロースが分解されて利用されるが,この場合でも,その動物そのものがセルラーゼを生産するのではなく,間接的な方法がとられている。たとえば,ウシの胃(反芻胃)は四つの部分から成るが,その最初の二つに微生物が寄生していてセルラーゼを生産し,飼料中のセルロースを分解してグルコースとする。この微生物がグルコースを発酵して生ずる脂肪酸が,ウシの胃から吸収されて利用されるのである。またウシの後の二つの胃は,前の胃で増殖した微生物を消化して,アミノ酸,糖などとして吸収する。シロアリもセルロースを消化するが,これもその消化管に寄生しているトリコニンファTrichonymphaと呼ばれる原生動物がセルラーゼを分泌するためである。
執筆者:村松 喬
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
植物細胞壁の主要構成成分。自然界にもっとも多く存在する有機化合物であり、植物中で二酸化炭素と水とから光合成によってつくられている。繊維素ともいう。セルロース鎖(α-1,4グルカン)が40本くらい集まった単位をミセルとよび、ミセルが多数集合したものをミクロフィブリル(微繊維)という。ミクロフィブリルは太さ10~30ナノメートル、長さ数マイクロメートルの大きさである。細胞壁の中で、セルロースはミクロフィブリルを形成する(セルロースミクロフィブリル)。これは、他の非セルロース性多糖類(ヘミセルロースおよびペクチン質)からなるマトリックス(細胞質基質)の中に埋没されたような状態で存在している。一次細胞壁(最初に形成される細胞壁)中のセルロース含量は、植物の種類や組織、年齢によって異なるが、普通、細胞壁乾燥重量の25~40%を占める。一般に若い細胞の細胞壁では、セルロース鎖を構成するグルコースの重合度は小さく、セルロース含量も少ない。さらに、ミクロフィブリルは細胞の長軸(茎軸)方向に対してほぼ直角に配列しているのみならず、ミクロフィブリルどうしの間隔も広い。このことは、若い細胞では細胞壁構造が長軸方向に対して機械的に柔軟であり、したがって、長軸方向へ伸長成長しやすくなっているということと関係がある。加齢した細胞では、セルロース分子のグルコース重合度も大きくなり、含量も増加して、60%に達することもある。また、若い分裂組織の細胞に比べて、成熟した細胞では、細胞当りのセルロース含量は約10倍にもなる。この際、ミクロフィブリルどうしの間隔も密になり、配列も長軸と平行のものが多くなり、全体としては網目構造を形成するようになる。この状態になった細胞壁は、機械的に強い構造となり、伸展性は少なくなる。したがって、細胞の成長も停止することになる。
植物細胞壁におけるセルロースミクロフィブリルの配列方向は、微小管の配列方向によって決められる。
[勝見允行]
植物体においてセルロースの占める割合はふつう1/2~1/3であるが、ジュートでは65%、アマでは80%、木綿では98%に及ぶ。木綿をアルコールとエーテルで洗うことにより、ろう、脂肪を除き、約1%の希アルカリ溶液で煮沸することによりペクチン様物質を除くと、純粋なセルロースが得られる。木材など普通の植物繊維はリグニン、ヘミセルロースなどを含むため、硫酸塩法、亜硫酸塩法によりこれを取り除くと、パルプとよばれるセルロースが得られるが、パルプには非セルロース分がまだ残っている。
[谷利陸平]
セルロースはD-(+)-グルコースのみより構成される多糖類(C6H10O5)nであり、グルコース単位は六員環いす形構造をもつヘミアセタールであるグルコピラノース環となり、そのC1位にあるヒドロキシ基-OHと、隣接するグルコース単位のC4位ヒドロキシ基との間で水1分子を失った形のβ-1,4-グルコシド結合により直鎖状につながった構造をとっている。結合をつくる二つのヒドロキシ基はいずれも赤道方向ともいうべきエクアトリアル位(赤道という意味)にあるため、グルコース単位は直鎖状に並ぶ。セルロースとデンプンは、結合をつくる位置と方向が多少違うことから、立体配置が異なっている。セルロースの分子量は数十万またはそれ以上のこともあり、1分子につき存在するグルコース単位の数(分子式でのnの数)は3000~6000個、セルロース分子の長さは2万~3万オングストローム(Å。10-10メートル)といわれている。これら鎖状セルロース分子において、数多くのヒドロキシ基が残っており、いずれもエクアトリアル位にあるため分子間の水素結合によって結び合い、平行に規則正しく配列し束をつくっている。束の両端は房状となり、他の房と網状につながりセルロース繊維となっていると考えられている。実際に天然セルロースにおいては微結晶部分と無定形部分とが認められ、このような構造が繊維の強さ、弾力性、染色性、吸湿性を生んでいる。セルロースを溶解したのち再生すると結晶構造は変わる。木材ではセルロース分子の束がリグニンの中に埋め込まれ、さらに強化された構造となっている。セルロースのなかには、D-マンノース、D-キシロースを含むもの、またD-グルコースが多少枝分れして結合しているものも例外的にある。
地球上のグルコースがエナンチオマー(光学異性体、鏡像異性体)の一方D-(+)-グルコースのみであるため、セルロースもエナンチオマーの一方のみである。
[谷利陸平]
セルロースは吸湿性の強い無味・無臭の白色固体で、普通のセルロース試料は10%程度の水を含んでいる。水、エタノール(エチルアルコール)、エーテルなど通常の溶媒には溶けず、シュワイツァー試薬(銅アンモニア溶液)、硫酸、塩酸、リン酸に溶ける。
セルロースを希酸と煮沸すると加水分解されD-グルコースが得られるが、デンプンなど他の多糖類と比べると加水分解されにくく、アルカリでは加水分解されない。セルロースのグルコース単位はそれぞれ3個のヒドロキシ基をもち、反応はこのヒドロキシ基でおこる。すなわち、セルロースはアルコールと同様にエステル、エーテルを生成する。硝酸、硫酸の混合酸を作用させると硝酸エステルであるニトロセルロースが、クロロスルホン酸を作用させると硫酸セルロースが得られる。また無水酢酸と少量の硫酸で処理するとトリアセテートとなり、これを部分加水分解すると、鎖の開裂、アセチル基の部分脱離がおこり、200~300単位のジアセテートすなわちアセチルセルロースが生成される。アルカリで処理して得たアルカリセルロースに二硫化炭素、ハロゲン化アルキル、クロロ酢酸塩を作用するとセルロースキサントゲン酸塩やメチルセルロース、CMC(カルボキシメチルセルロース)などのセルロースエーテルが得られる。これらセルロースの反応は、安価で入手しやすい既製の高分子の性質を改良するため行われ、工業的に重要な意味をもっている。
[谷利陸平]
セルロースは繊維、紙として大量に用いられるほか、セルロース誘導体は火薬、プラスチック、フィルム、ビスコースレーヨン、消泡剤、乳化分散剤、増粘剤、光学分割剤、保護コロイドなどとして用いられ、イオン交換セルロースは生化学分野での用途が多い。かつて多く製造されたセルロイド、セロファンもセルロース誘導体である。
[谷利陸平]
人間の消化管には繊維素分解酵素(セルラーゼ)は分泌されないから、セルロースは消化吸収されないで排泄(はいせつ)される。しかし大腸に生育する微生物のあるものはセルロースを分解する。セルロースは食品分析における「粗繊維」の主成分である。また腸の蠕動(ぜんどう)運動を促進し、便秘を防ぐ。
[不破英次]
『右田伸彦・米沢保正・近藤民雄編『木材化学』上(1968・共立出版)』▽『木材工業ハンドブック編集委員会編『木材工業ハンドブック』改訂3版(1982・丸善)』▽『増田芳雄著『植物の細胞壁』(1986・東京大学出版会)』▽『村尾沢夫ほか著『セルラーゼ』(1987・講談社)』▽『桜井直樹ほか著『植物細胞壁と多糖類』(1991・培風館)』▽『越島哲夫編『セルロース資源――高度利用のための技術開発とその基礎』(1991・学会出版センター)』▽『井手文雄著『界面制御と複合材料の設計』(1995・シグマ出版)』▽『松永是・本宮達也編著『おもしろいバイオ新素材のはなし』(1996・日刊工業新聞社)』▽『宮本武明ほか編『21世紀の天然・生体高分子材料』(1998・シーエムシー)』▽『化学工学会編『先端材料制御工学』(1999・槇書店)』▽『セルロース学会編『セルロースの事典』(2000・朝倉書店)』▽『磯貝明著『セルロースの材料科学』(2001・東京大学出版会)』▽『越島哲夫ほか著『機能性セルロース』(2003・シーエムシー出版)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
繊維素ともいう.植物の細胞壁を構成する主成分であり,天然にもっとも豊富に産出する有機物質の一つ.セルロースは,グルコースがβ-1,4-グルコシド結合した多糖類で,化学式は (C6H10O5)n で示される.
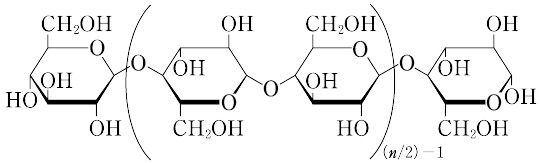
セルロースは,一般に,植物体中のリグニン,ヘミセルロースなどの非セルロース成分を除去,精製して得られるが,多くの場合,非セルロース成分の除去・精製にいろいろの工夫がされている.その方法は,亜塩素酸塩,過酢酸などによってリグニンを選択的に分解除去し,おもにセルロースとヘミセルロースからなるホロセルロースを得たのち,アルカリ抽出によってヘミセルロースを中心とした非セルロース成分を除去して調製される.また,クラフト法,サルファイト法で得られるクラフトパルプ,サルファイトパルプなどの化学パルプを漂白し,残存するリグニンおよびヘミセルロースを除いて得ることもできる.天然セルロースは分子が集合し,部分的に微結晶を形成している(セルロース-Ⅰ).また,結晶構造の異なる水和セルロース(セルロース-Ⅱ),セルロースの銅アンモニア化合物を分解するときに得られるセルロース-Ⅲ,セルロースの高温再生によって得られるセルロース-Ⅳなど,各種の結晶変態が存在する.セルロース-Ⅰの結晶形には,三斜晶系の Iα と単斜晶系の Iβ があり,天然セルロース中では両者がセルロースの起源によって異なる割合で混在していることが知られている.微結晶以外の部分は非晶領域とよばれ,分子は秩序ある配列をしていない.さらにこれらの繊維が集合して,フィブリルを形成している.このフィブリルの存在は,セルロースの膨潤,こう解などでみとめられる.生体内では,グアノシン三リン酸とα-D-グルコピラノシルリン酸から生成するグアノシン二リン酸-D-グルコースが,アクセプターと順次反応してグルコース単位を増して合成される.セルロースの分子量は数万~数十万に及び,ヒドロキシ基をもっているにもかかわらず,水には溶けず,アルカリで膨潤する.セルロースは各グルコース単位ごとに3個のヒドロキシ基があるため,セルロースエステル,セルロースエーテルなどのセルロース誘導体をつくることができる.その形態をそのまま利用した紡織繊維,紙などの製造がおもな用途であるが,各種セルロース誘導体についても,多様な用途がひらかれている.
出典 森北出版「化学辞典(第2版)」化学辞典 第2版について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
Sponserd by ![]()
…動物の骨格構造を構築している主要な構成成分であり,また生体を保護する役割を果たしているタンパク質でもある。植物界では硬タンパク質の代りにセルロース類が同じ役割をしているものと考えられている。 例としては骨,皮,腱などに含まれているコラーゲン,靱帯や動脈などの成分であるエラスチン,毛髪,羽毛などのケラチン,絹のフィブロイン,カイメンのスポンジ(海綿質)などが知られている。…
…しかし互いに類似の化学構造の分子からなる物質でも,分子量が1000程度のものと1万以上のものとではとくに物理的性質に大きな差異があり,一般には分子量が数千以上のものを高分子と呼ぶ。天然に存在する高分子にはセルロース,タンパク質,核酸などがあり,それぞれ生物にとって必須の役割を担っている。そのなかでもセルロースは古くから人類によって被服の材料などとして利用されてきた。…
…いずれについても生物種によって多糖の種類が大きく異なっている。高等植物の細胞壁多糖はセルロースを主体としている。エビ,カニ,昆虫の殻の主体をなすのはN‐アセチルグルコサミンからなるキチンである。…
…単糖の例としてはブドウ糖(グルコース),ガラクトース,また少糖の例としてはグルコースが2分子結合した麦芽糖(マルトース),グルコースと果糖(フルクトース)が結合したショ糖(砂糖),グルコースとガラクトースが結合した乳糖(ラクトース)をあげることができる。多糖にはデンプン,グリコーゲン,セルロースなどがある。
[単糖と多糖の関係]
グルコースとデンプンを例にとって説明する。…
※「セルロース」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
1 花の咲くのを知らせる風。初春から初夏にかけて吹く風をいう。2 ⇒二十四番花信風...