目次 呼吸研究の歴史 呼吸の生化学 動物の呼吸 ヒトの呼吸 呼吸運動 肺気量と肺容量 肺胞におけるガス交換 血液によるガス輸送と組織におけるガス交換 呼吸中枢と呼吸調節 呼吸の異常 呼吸器官respiratory organ 無脊椎動物の呼吸器官 脊椎動物の呼吸器官 呼吸法の文化史 呼吸研究の歴史 いき(息)もの,英語のアニマル(ラテン語のanima=息,生命に由来)などの語からもわかるとおり,呼吸と生命は古来密接に結びつけられてきた。空気中のプネウマpneuma(精気)が体内に取り込まれて生体を活気づけるという考えはギリシアにひろく見られ,アリストテレス は3種の精気を区別した。2世紀のローマのガレノスは自然精気が消化,栄養,排出などのいわゆる植物性機能をになうと考えた。こうして呼吸を具体的な生命現象と関連させる努力はなされたが,思弁にとどまった。
W.ハーベーが血液循環を確証(1628)して以来,研究は新しい段階に入り,血流にのって全身に分配される空気の重要成分は何かが問題になった。16世紀の錬金術的化学者・医者パラケルススは,〈硫黄精〉と〈硝石精〉によって燃焼は起こり,かつ呼吸と燃焼は同じ現象であると考えていた。この解釈は,17世紀に呼吸について実験したR.ボイル,メーヨーJohn Mayow(1640-79)などに影響を与えた。18世紀にはフロギストン(燃素)説 の誤りを経て,ラボアジエが燃焼での酸素の役割を確定する。呼吸も体内での酸化として位置づけられたが,熱をだす燃焼と同じことが体内でも起こると考えられたので,J.L.ラグランジュ は,肺のみで燃焼が起これば肺は高熱になりすぎると論じた。ここからかえって,酸素は全身末梢組織に分配されるはずだとの正しい見通しが生まれた。
20世紀初めのエネルギー代謝の研究は,酸素を用いない解糖,発酵を中心にしていたが,有機酸から水素を奪う脱水素酵素の作用も,1920年代から明らかになる。呼吸とは酸素が直接に基質を〈燃やす〉のではなく,基質から奪われた水素が酸素と出会うことであるとの理解が,こうして整ってきた。細胞内で水素を授受する機構については,セント・ジェルジ などもモデルを提出したが,H.A.クレブスのクエン酸回路 (1937)が正しい回答をあたえた。第2次大戦後の研究の発展で,呼吸の反応成分はすべてミトコンドリア に局在することがわかった。こうして,プネウマ説にはじまり,肺での空気の出入りを意味していた〈呼吸〉は,現代生物学の用語では,細胞での基質からの水素原子の取出しと,その水素を酸素と化合させる際のエネルギーを用いてのATP生成(酸化的リン酸化)を意味するに至っている。長野 敬
呼吸の生化学 以上のような歴史的経過から,呼吸は古くは動物の呼吸運動をさす言葉であったものが,動物の外界とのガス交換(これを外呼吸という),さらに体液と細胞のガス交換(これを内呼吸という)をも含める言葉となった。そして,現在,生化学的には呼吸は次のように定義されている。(1)狭義には生体が分子状酸素O2 を最終電子受容体として有機化合物 を二酸化炭素CO2 ,水H2 O,アンモニアNH3 などの無機化合物にまで酸化的に分解し,生体に利用可能な形で(ATPとして)エネルギーを蓄えることを呼吸という。(2)この定義を拡張し,無酸素条件下で分子状酸素の代りに硝酸塩,硫酸塩を最終電子受容体として上記と同様の過程が行われる場合をも含める。これはある種の嫌気性細菌が行う呼吸で,それぞれ硝酸呼吸,硫酸呼吸という。(3)さらに広義には,アルコール発酵 や解糖のように,外から最終電子受容体の供給がなくても,基質から生じた化合物の間で酸化還元が行われ,この過程でATPが生成される場合をも含む。この場合を無酸素呼吸,無気呼吸,嫌気呼吸,または分子間呼吸といい,これに対し(1)の場合を酸素呼吸または好気呼吸という。
呼吸の強さを知るには,酸素消費量または二酸化炭素放出量を測定すればよい。前者の測定には酸素電極法,検圧法などが用いられ,後者の測定には赤外線ガス分析法,電気伝導度測定法などが用いられている。二酸化炭素放出量(モル)と酸素消費量(モル)の比,[CO2 ]/[O2 ]を呼吸商respiratory quotient(略称RQ)という。実際には容積比として求めることが多い。グルコース(糖類)が完全酸化するときはRQ=1(C6 H1 2 O6 +6O2 =6CO2 +6H2 O)。脂肪やタンパク質のように分子中の酸素原子の割合が糖より少ない物質ではRQ<1(ステアリン酸C1 8 H3 6 O2 +26O2 =18CO2 +18H2 O,RQ=0.69)。逆にリンゴ酸のような有機酸の場合はRQ>1(C4 H6 O5 +3O2 =4CO2 +3H2 O,RQ=1.33)。ただし,基質が同じでも呼吸経路によりRQは異なる。たとえば,アルコール発酵のような二酸化炭素放出を伴う無気呼吸が併行して行われるとRQは上昇する。
グルコース酸化の最終的な反応式だけを見ると,生体外での燃焼も呼吸もまったく同じであるが,両者は次の点で異なる。(1)生体外での燃焼を引き起こすには発火点まで熱しなければならないが,生体内での酸化は酵素により低い温度のもとで段階的に進行する。(2)生体外での燃焼時に遊離されるエネルギーは,ほとんどすべて熱の形で放出されてしまうが,呼吸では遊離されるエネルギーは高い効率でATPに蓄えられる。
呼吸の反応系は,多くの場合次の二つの過程に大別される。(1)基質分子が脱水素され(これと共役して水素受容体が還元され),基質分子の炭素鎖が切られる。(2)還元された水素受容体から最終電子受容体へ電子が渡される。呼吸の代謝経路の典型的なものは,解糖 系(図1)で生じたピルビン酸 が,クエン酸回路 (図2)で完全酸化を受ける場合である。解糖系の諸酵素は細胞質の可溶性画分にあると考えられているが,細胞質中の微細構造に結合していると主張する研究者もある。真核生物では,クエン酸回路,β酸化,および電子伝達系の諸酵素はミトコンドリア に含まれている。赤血球や肝臓および多くの植物の組織では,グルコースの一部は解糖系とは異なるペントースリン酸回路 (細胞質の可溶性画分にある)によって完全酸化を受ける(図3)。解糖系で働く補酵素がNADであるのに対し,この回路の場合はNADPであることと,回路上に五炭糖が出現することが特徴である。脂肪酸が分解されるときは,脱水素されたのちカルボキシル末端から炭素原子C2個きざみで炭素鎖が切られアセチルCoA を生じる(β酸化。図4)。これがクエン酸回路へ入り完全酸化を受ける場合と,グリオキシル酸回路 (マイクロボディ にある。図5)へ入りもう一度脱水素されたのちコハク酸となる場合がある。緑色組織で光のもとで行われるグリコール酸 の酸化的分解は光呼吸photorespirationと呼ばれるが,ATP生成がないから本来の呼吸とは生理学的な意味が異なる。
呼吸過程の最後の部分で,NADHやコハク酸から電子を分子状酸素に渡す経路を電子伝達系 または呼吸鎖という(図6)。この系は酸化還元電位 の異なる多数の成分から構成されており,電子は酸化還元電位の低いものから高いものへと流れてゆく。電子伝達にはフラビン,ユビキノン,ヘムのほかに非ヘム鉄,銅イオンも関与している。この系の成分は4種の複合体の形でミトコンドリア内膜に存在する(図7)。この電子伝達と共役して行われるATP生成を酸化的リン酸化反応という(図8)。ADPとオルトリン酸 の濃度が下がると,呼吸鎖の反応速度(電子の流れの速さ)が低下する現象を呼吸調節respiratory controlといい,基質と酸素の不必要な消費を避け,ATPの消費に見合った供給を行うしくみと考えられている。細菌においては細胞膜にミトコンドリアのものと類似の呼吸鎖が存在する。
グルコース1molが解糖とクエン酸回路を経て酸化されるとき生成するATPは,前核生物では38mol,真核生物では36molである(表1)。グルコース1molが完全酸化されるときの自由エネルギーの生成は686kcalである。ADPからATPを生成するのに必要な自由エネルギーを7.3kcalとすれば,ATPが38mol生じる場合を考えると,呼吸によって277.4kcal,すなわち約40%の効率で,ATPの中にエネルギーが蓄えられたことになる。アルコール発酵ではグルコース1mol当りATPが2molしか生成しない。大気中に分子状酸素が大量に存在するようになったのは光合成生物の出現以後であるから,酸素を用いる効率のよい呼吸系は,無気呼吸を基礎として代謝系の進化の結果,あとからでき上がったものと考えられる。
呼吸の第1の生理学的意義はATPの生成であるが,細胞構成成分の前駆体(表2)およびこれら成分の合成に必要な還元力(NADPH)を供給するというもう一つの重要な役割をもっている。辻英夫
動物の呼吸 18世紀の末になって,呼吸の実体は,動物が外界からO2 を吸収し,外界にCO2 を排出することであることがわかった。さらに呼吸器官から取り込まれたO2 は,血液によって体中の細胞にまで運ばれ,CO2 は逆に細胞から放出されて,血液によって呼吸器官に運ばれるということも明らかにされた。そこで動物と外界とのガス交換を外呼吸,体液と細胞とのガス交換や細胞内での酸化的分解過程を内呼吸(細胞呼吸,組織呼吸)といって区別するようになった。また,血液中のヘモグロビン のように,O2 の運搬の働きをしている色素を呼吸色素 と呼んでいる。細胞の中では炭水化物,脂肪,タンパク質などの有機化合物が分解され,その中に含まれている化学エネルギーが取り出されて,細胞のあらゆる生命活動のエネルギーとなるのであるが,この分解にO2 が利用され,分解の最終産物としてCO2 が生じるのである。
外呼吸の際,O2 とCO2 の移動は,体表や呼吸器官の膜を介しての拡散によって行われる。小型の水生動物,たとえばゾウリムシ,アメーバ,プラナリアなどは体表における拡散だけで体中に十分なO2 を補い,CO2 を放出できる。しかし,体の厚みが1mm以上になると,拡散だけでは体の深部まで十分にO2 をいきわたらせることはできない。とくに活動性が低く酸素要求の少ないものを除くと,ほとんどの動物は体表から深部へのO2 ,あるいは逆方向のCO2 の運搬のために,体液の循環を利用している。また昆虫では細かく分岐して体中にいきわたっている気管系が,空気を直接組織に供給している。水生動物の呼吸器官であるえらは,体表の一部の表面近くに血管の分布がとくに密になっているところであるが,ガス交換の面積を増大するためにひだとなっているのが普通である。これに対して陸生動物の肺は空気呼吸に適した器官であって,脊椎動物では食道の一部のふくらみとして形成され,空気をその中に吸い入れることによってガス交換面の乾燥を防ぐことができる。えら呼吸でも肺呼吸でも,たいていの動物では呼吸器表面をたえず新鮮な水や空気にふれさせておくために,呼吸運動によって換水や換気が行われている。魚類では,呼吸運動によって起こるえらの表面の水流の方向と,えらの毛細血管内の血流の方向とが逆になっていて,ガス交換がきわめて効率よく行われることが知られている。また,カツオやマグロのように,たえず泳ぎまわっている魚は運動によって換水をしており,泳がないと呼吸困難になるといわれる。
空気中には容積比にして約20%のO2 と0.03%のCO2 が存在する。1気圧で空気と平衡状態にある水1l 中には5℃で9.22ml ,20℃で6.51ml ,海水にはそれぞれ6.89ml ,5.05ml のO2 が含まれている。空気の重さは水のほぼ1/1000であり,同じ量のO2 を含む水と空気の重さを比較すると,空気は水の約1/30000に過ぎない計算になる。さらに空気は,水に比べて粘度もずっと少ないこともあって,空気中で呼吸運動に費やす労力は,水中よりずっと少なくてすむはずである。魚が呼吸のために使うエネルギーは,哺乳類の20倍以上にも達するという報告もある。
えらや肺をもつ動物でも,ある程度は皮膚でガス交換が行われている。たとえば,カエルで調べたところでは,冬眠中は必要なO2 の2/3以上を皮膚からとっており,CO2 の排出は年間を通じて肺よりも皮膚からの方が多い。皮膚呼吸のために,カエルの皮膚はつねに湿っている必要があり,これがカエルが水辺から離れられない理由の一つである。鳥類や哺乳類では皮膚呼吸の割合はきわめて少ない。
呼吸量や代謝量は体が大きい動物ほど大きいが,体重に比例するのではなく,体重の3/4乗に比例して増加するとされている。この関係は同種の個体間にも,また一つの分類群に属する異種間にもあてはまる。たとえば,哺乳類について体重の対数を横軸に,個体当りのO2 消費量の対数を縦軸にしてプロットすると,小さなネズミから大きなゾウまでの種々の動物が,3/4のこう配をもつ直線上にならぶ。個体の呼吸量が体重の3/4乗に比例するならば,単位体重当りの呼吸量(呼吸度)は体重の1/4乗に反比例することになる。すなわち,小型の動物ほど呼吸度は大きい。たとえば体重1kg当り1時間のO2 消費量は,体重25gのネズミでは1.65l であるのに対し,体重が4tに近いゾウでは0.07l である。一般に活発な動物は不活発なものより呼吸度は大きい。また同じ個体でも,活動の程度によって呼吸量は大きく変化する。ヒトが走るときは,呼吸量は速さに比例して増加し,静止状態の呼吸量の10倍以上にも達する。昆虫類では飛翔(ひしよう)中に呼吸量が100倍に増加することがある。呼吸量はまた温度,酸素濃度などの環境要因によっても変化することが知られている。佃 弘子
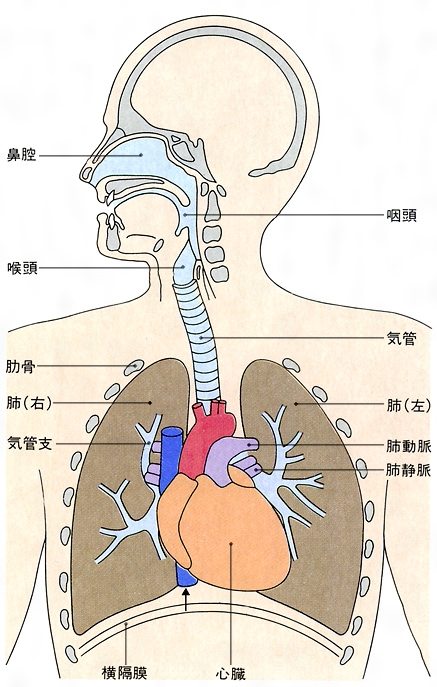
ヒトの呼吸 アメーバのような単細胞生物では,生体と外界との間に行われるO2 とCO2 の拡散のための移動距離が短いので,細胞表面で直接,周囲の環境とガス交換が行われる。しかし,高等な多細胞動物になると,ガス交換のための特別なしくみが必要となる。ヒトや哺乳類では肺の中に空気を吸い込んで,全身分のO2 とCO2 の交換をまとめて行い,肺と体内の組織との間のガスの運搬は血液循環のシステムによっている。循環系の分岐によって,毛細血管とその周辺の組織との間の距離は短縮され,ガスの拡散によるガス交換が可能となる。十分にO2 を含み,CO2 の少ない動脈血が毛細血管に到達すると,ガスの濃度こう配に従ってO2 は組織へ移り,CO2 が組織から血液へ移る。ここで血液は静脈血となって心臓の右心房,右心室を通って肺へ送られ,肺の中の気道の末端分岐部に位置する肺胞に接する毛細血管に達する。ここで拡散によって血液から肺胞へCO2 ,肺胞から血液へO2 のガス交換が起こり,血液は動脈血化し,左心房,左心室を経由して体内の組織へ送られる。
吸気は気道(口腔,鼻腔,咽頭,喉頭,気管,気管支,細気管支)を通過するうちに水蒸気で飽和され,呼気も完全飽和の状態にあり,このことは体内の水の収支に一つの役割を演じている。
測定によって成人は1分間に約250ml のO2 を消費し,約200ml のCO2 を呼出する。一定時間内のCO2 の排出量とO2 の摂取量の比,呼吸商(RQ)は,呼吸によって燃焼される栄養素の種類に応じて,0.75~1.0の値をとる。大気と血液のガス交換は肺胞で行われる。気管支系の最も細い部分である細気管支が肺胞管に移行し,肺胞管は肺胞に分かれる。一つの細気管支に属する肺胞の集まりが直径1~1.5cmの肺小葉を形成する。肺小葉の内部に細気管支と肺動脈の小さい枝が入り込む。一つの肺胞は直径およそ0.1mmであるが,両側の肺について総計すると,肺胞の数は3億から7億5000万個の間にあり,ガス交換が行われる全肺胞面積は約70m2 と推定される。肺胞には弾性繊維と格子繊維からなる支持基質があり,肺胞の入口は平滑筋繊維に囲まれている。血管系は両側の肺で分岐して約300m2 の面積をもつ毛細血管網となって肺胞の表面にからみつき,ここで分圧の高低に応じてO2 とCO2 の交換が起こる。
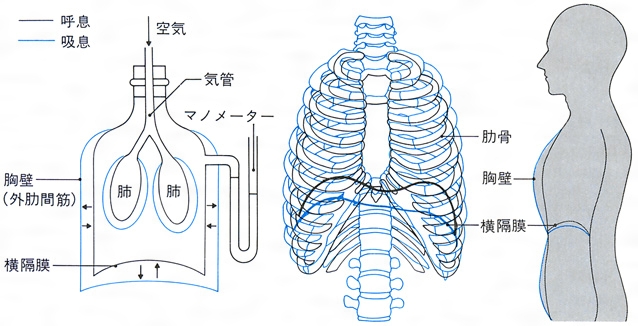
呼吸運動 肺胞と外界の間のガス交換,つまり換気は,呼吸運動によって生ずる気道の各部の圧力の差が原動力となって起こる。外呼吸のための運動を呼吸運動,それに働く骨格筋を呼吸筋という。呼吸運動は規則正しい吸息と呼息の交代である。吸息によって胸腔が拡張し,外気は気道から肺の中に吸いこまれ,呼息により胸腔は縮小して,肺内の混合ガスが押し出される。両側の肺はこの胸腔の運動に受動的に従う。吸息のときには肺容量が増加して肺胞内の圧力(肺内圧)は外界の圧力(大気圧)より低くなり,呼息のときには肺容量が減少して圧力差は逆となる。吸息は,横隔膜 が収縮(平坦化)して胸腔が下方に広がるとともに,外肋間筋が収縮して肋骨がひきあげられ,その他の補助呼吸筋の活動も加わって,結果として胸郭はひき上げられ,胸腔が前方と左右へ広がることにより起こり,受動的に肺へ空気が流入する。呼息は一般に受動的に起こり,胸郭自身の重さと弾性による胸郭の沈下(胸郭の収縮)が主体であるが,強制的に呼息が行われるときには,腹壁の筋肉群の活動(腹圧の上昇)によって,横隔膜が上方へ押し上げられ,それに内肋間筋の収縮が加わって胸腔の容積は減少し,肺は圧縮されて気道内のガス体は外へ向かって流れる。
肺は気管の部位だけで固定されていて,胸壁には付着していない。肺の表面と胸壁の内面は胸膜 でおおわれているが,両方の間の狭い空所を胸膜腔といい,安静呼息時における胸膜腔の内圧(胸腔内圧)は外気よりも約-4mmHg低い。ここにはムコ多糖類を含む胸膜液がある。安静時の呼吸運動による肺内圧(肺胞内圧)の変化は,吸息時には大気圧より0.7~2.2mmHg低く,呼息時には0.7~2.2mmHg高い。このほかに,呼吸運動に伴う肺の拡張,収縮には,肺の弾力繊維,肺胞の表面張力による収縮する性質,肺胞の表面張力を減少させる表面活性物質(レシチンを含むリポタンパク質 )の量,肺,胸郭の受動的伸展性を示すコンプライアンス ,気道抵抗,非弾力性組織抵抗などの種々の因子が関係し,これらはいろいろな呼吸器疾患の病態との関連からも重要視されている。
吸息は直ちに呼息へ移行するが,通常,呼息と次の吸息の間には休止期がある。呼吸数は新生児では1分間に29回(未熟児では34回),健康な成人では安静時1分間に平均11.7(10.1~13.1)回,軽労働時に平均17.1(15.7~18.2)回,重労働時に平均21.2(18.6~23.3)回で,24回をこえることはない。呼吸筋として横隔膜がおもに働く場合を腹式呼吸,肋間筋が主体となる場合を胸式呼吸というが,自然の呼吸運動では両方が共存している。おおよそ,肺活量の2/3が横隔膜の運動に,1/3が胸郭の運動によっている。男子では女子に比べて腹式呼吸の割合が大きく,とくに高齢者ではこの割合がさらに大きくなるが,これは加齢による胸郭の伸展性の減少のためと考えられている。
呼吸運動は,脳幹部の呼吸中枢で形成される自動的周期性興奮が,呼吸筋支配運動ニューロン を経由して呼吸筋へ送られることによって起こる。たとえば横隔膜は,第3・4頸髄の運動ニューロン群から発する横隔神経に支配されているので,それ以下のレベルでの脊髄損傷で肋間筋の運動麻痺が起こっても,横隔膜による呼吸運動は続けることができる。
肺気量と肺容量 安静時呼吸での呼息後,胸郭は中間状態,安静時呼息位(基準位)にある。安静時呼吸において出入する空気量,すなわち1回換気量は成人男子で約500ml である。さらに努力吸入により最大約2500ml (予備吸気量)を吸入できる。安静時呼息位からさらに強制呼息によって呼出できる量(予備呼気量)は最大1500ml である。このときなお肺内に残るガス量(残気量)が約1200ml ある。これらの基本的な肺気量はlung volume として表され,それらの組合せによる肺容量はlung capacity として表される。capacityとして表されるものには,たとえば全肺容量や機能的残気量がある。前者は前記の肺気量すべての和(約6000ml )であり,後者は予備呼気量と残気量の和(約2700ml )である。肺活量 もcapacityの一つであって,最大吸息位から最大呼息位,すなわち,1回換気量,予備吸気量,予備呼気量の和(約4500ml )をいう。したがって全肺容量は肺活量に残気量を加えたものでもある。残気量と全肺容量以外の各量はスパイロメーターspirometer(スピロメーター ともいう。肺気量計)によって測定され,肺気量測定はコンプライアンス,動作時の呼吸機能検査や肺機能検査に応用される。なお1分間に呼吸気道に入った外気量を分時換気量という。上にあげた種々の気量や容量は年齢,性別,体質,訓練によって個人個人で著しく異なる。したがって肺気量測定結果の絶対値よりも肺気量の変化のほうが重要であって,たとえば肺疾患の経過を監視するのはこのことによる。1回の安静呼吸によって肺胞に入る新鮮な外気量は350ml で,これが機能的残気量約2700ml と混合するのであるから,1回の安静呼吸の換気率は約1/8である。このことは肺胞気の組成が呼吸周期によって大きく変動せず,ほぼ一定の値を保つことができ,呼息相においても血液とのガス交換を可能にしている。
肺胞におけるガス交換 肺でのガス交換には肺胞換気が重要である。しかし呼吸系のうち肺胞までの空気の通過経路となる部分,すなわち口腔,鼻腔,咽喉頭腔,気管,気管支などはガス交換に関与しないので,これらの諸部分の空間容積(約150ml )を死腔または解剖学的死腔という。ところが種々の原因で一部の肺胞が機能的にガス交換に関与しないような場合,これも一種の死腔と考えられるので,この死腔を前記の解剖学的死腔に加えたものを機能的死腔(または生理学的死腔)といっている。健康人ではこの二つの死腔はほぼ等しいが,病的状態では機能的死腔が解剖学的死腔よりも大きくなる。解剖学的死腔は吸気が肺胞へ到達するために必要な経路となること,死腔で吸気の清浄化,加湿,加温が行われること,死腔の一部が発声器官の一部を構成していることなど,重要な機能を果たしている。
1回換気量のうちの肺胞換気量と呼ばれる部分だけが肺胞に接する。したがって肺胞換気の程度は,肺胞ガスの組成に直接影響を与えるので重要である。健康人の肺胞換気量は1分間当り約4l ,これに対する肺毛細血管血流量 は約5l /分で,その割合,換気血流比は約0.8である。外気が呼吸気道を通って肺胞に達すると水蒸気で飽和され,37℃に加温される。水蒸気の圧力は47mmHgで,肺胞気の圧力は大気圧に等しいから,肺胞気のO2 分圧は104mmHg,CO2 分圧は40mmHgである。呼気は死腔のガスと肺胞気との混合物であり,安静時の呼吸では呼気の1/3が死腔気,2/3が肺胞気で,そのガス組成は水蒸気飽和空気と肺胞気の中間値となる。
肺胞の平均直径は約0.1mm,ガス交換が起こる全肺胞面積は約70m2 と推定されている。それに接する毛細血管には75~100ml の血液が含まれ,ミクロン単位の厚さの膜をへだてて約2000ml の肺胞気との間でO2 とCO2 の交換が行われる。ガスの拡散は圧力のこう配によって起こる。肺胞気のO2 分圧は100mmHg,静脈血では40mmHgで,その差は60mmHgである。CO2 分圧はそれぞれ肺胞気で40mmHg,静脈血では46mmHg,その差は6mmHgであるが,CO2 の膜の拡散速度はO2 のそれの20倍もあるので,十分な交換が行われる。拡散に有効な膜の面積は拡散の過程に影響し,正常面積の1/3~1/4になると安静時でも換気障害が起こる。激しい運動時には膜の表面積のわずかな減少でも影響が大きい。肺胞気と血液の間には肺胞上皮,基底膜,毛細血管内皮があり,これらを一まとめにして呼吸膜(肺膜)といい,厚さは0.1~1μで,ガスはこれらを通過せねばならない。毛細血管の直径は7μ前後で,ほぼ赤血球の大きさである。O2 もCO2 も呼吸膜を容易に通過するが,肺浮腫などで間質液が貯留すると拡散距離が長くなって,換気不全が起こる。
拡散の距離は短く,血漿(けつしよう)までが約1μ,赤血球内で約1μと見積もられ,肺胞と赤血球が互いに接触している時間(1秒以下)内に分圧の平衡がなりたつと考えられる(血液から肺胞気へのCO2 の拡散はきわめて速く,0.4秒以内にほぼガス平衡に達する)。動脈血化した毛細血管内の血液のO2 分圧,CO2 分圧は肺胞のそれらとほぼ等しいとみなされる。ガスの呼吸膜での拡散のしやすさを拡散能といい,1mmHgの圧力差で1分間に肺胞気と血液との間を移行するガス量で表す。成人の安静時のO2 の拡散能は21ml /分・mmHg O2 分圧で,激しい運動時には肺胞が伸展されて表面積が増し,呼吸膜の厚さが減少すること,毛細血管が拡張して換気面積が大きくなることなどによって,値が3倍に増加する。CO2 の呼吸膜の通過があまりに速やかなので,肺胞気と毛細血管血液とのCO2 の圧力差は1mmHg以下で,現在の技術では測定できないが,拡散能はガスの拡散係数にほぼ比例するから,O2 のそれの20倍として約400ml /分・mmHg CO2 分圧と推定される。
血液によるガス輸送と組織におけるガス交換 血液路によるO2 およびCO2 の輸送のあらましは次のようである。血液中のO2 はおもに赤血球のヘモグロビンと可逆的に結合し,体循環によって組織に運ばれる。動脈血はおよそ21容量%のO2 を結合しうる。静脈血も,なお12~14容量%のO2 を含んでいる。細胞からは炭酸が血漿の中に出て,一部はそのまま溶けて,一部は重炭酸塩に,一部はタンパク質と結合して血漿中を運ばれ,肺胞の中で呼気に移る。100ml の血液中には50ml の炭酸が含まれているが,2ml が血漿中に溶けていて,およそ40ml は重炭酸塩類の形で化学的に結合し,一部は重炭酸ナトリウム として血漿中に,一部は赤血球内に重炭酸カリウムとして存在する。およそ8ml は血色素と化学的に結合してカルバミノヘモグロビンとなっている。ガス交換に際して解離されうる炭酸のごく一部分だけが放出されている。
血液によって運ばれたO2 が細胞内に取り込まれて,細胞内の呼吸酵素の働きによって,栄養素などの有機化合物の酸化が起こる物質代謝の過程は多様であって,多数の酵素がそれぞれ定まった反応段階で作用して進められる酸化の過程には,O2 の添加によってスタートする好気性反応と水素を放出する嫌気性反応とがある。酸素消費によるエネルギーの産生は糸粒体(ミトコンドリア)の中で起こり,エネルギー貯蔵物質としてアデノシン三リン酸 (ATP)がつくられ,呼吸の反応によって遊離されるエネルギーはATPの化学結合エネルギーの形でプールされ,必要に応じて細胞の生理作用の維持など種々の仕事に用いられることになる。
組織の毛細血管部で,周囲の組織との間にガス交換(組織呼吸)が起こる。組織液のO2 分圧は細胞の代謝活動によって変化するが,平均40mmHg(20~45mmHg)で,動脈血O2 分圧との間に55mmHgの分圧差があって拡散によって平衡に達し,毛細血管を流れていく血液のO2 分圧は40mmHgとなっている。一般に細胞内は細胞外液よりもO2 分圧が低く(平均6mmHg),拡散によってO2 は組織液から細胞内へと移っていく。細胞内代謝が正常に起こるために必要なO2 分圧は1~5mmHgである。細胞や組織のCO2 分圧は平均45mmHg,動脈血CO2 分圧は40mmHgで分圧差は小さいが,CO2 は拡散性が大であるから速やかに平衡に達する。
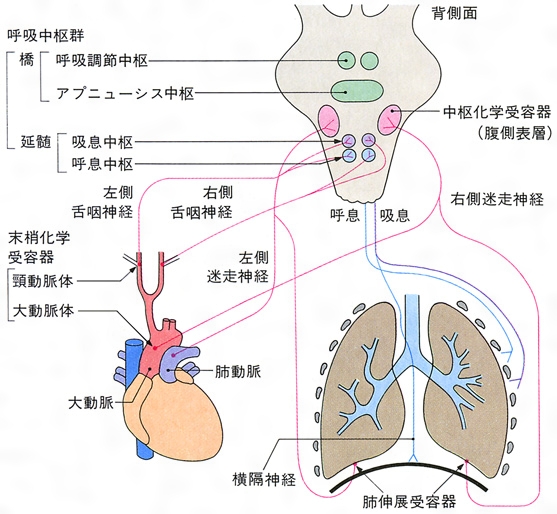
呼吸中枢と呼吸調節 生体の酸素需要と炭酸ガスの産生は時々刻々変化していて,これに応じて呼吸運動の頻度と深さが調節され,肺胞換気が増減して体のO2 分圧,CO2 分圧が一定に保たれる。このような呼吸調節は,脳幹の呼吸中枢による横隔膜と肋間筋の周期的収縮を主体とする呼吸運動の制御,呼吸反射による神経性調節,それと体内のO2 分圧,CO2 分圧,水素イオン濃度 (pH)などの化学的呼吸調節因子による化学調節のメカニズムによって,絶えず促進的,抑制的な影響を受けて行われている。
呼吸運動は,胸腹部の広い範囲に存在する横隔膜や肋間筋など,多数の呼吸筋の秩序正しい協調活動によって,円滑な吸息運動と呼息運動が交代することで維持されている。呼吸筋の活動を支配している多数の運動ニューロンは脳と脊髄の広い領域に分布しているが,これらは脳幹部の特定の領域にある呼吸中枢によって統合的に制御されている。以下に述べる多数の呼吸調節のメカニズムの大多数は,フィードバック型のループをつくっていて,神経性および化学調節の調節因子による中枢神経系への求心性入力のほとんどすべてがこの呼吸中枢に収束し,呼吸調節の効果は呼吸中枢からの神経性出力の変化として現れてくる。
呼吸反射による神経性調節には,気道にある肺伸張受容器から起こるヘーリング=ブロイエル反射 ,肺刺激受容器,肺胞壁のJ受容器から起こる反射,肺血管反射,肺化学反射などをふくむ肺迷走神経反射 がある。また,気道とくに咽頭,喉頭,気管,気管支内面にある咳の受容器の機械的および化学的刺激によって発生する求心性インパルスが,肺迷走神経内を上行して延髄に達してひき起こされる咳 の反射,鼻腔内の受容器から三叉神経内を上行する求心性インパルスによって起こるくしゃみ の反射のように,防御反射としての意味をもつ反射がある。このほか,皮膚,粘膜の化学刺激,温熱,機械的刺激などに応ずる非特異的受容器,骨格筋と腱の固有受容器からの反射も知られている。呼吸運動は精神感動,睡眠,身体運動によっても変化し,また,ある範囲内では随意的に促進(深呼吸),抑制(息こらえ)することができることから考えて,脳の高位中枢(前頭葉,辺縁系,視床下部,その他上位の脳,脳幹部,脊髄に存在する呼吸関連領域,体温調節,発語,発声,嘔吐,嚥下,あくび に関係している中枢内の部位)との機能的結合があると推定されている。循環反射のメカニズムでもある頸動脈洞,大動脈弓の圧受容器も呼吸運動に影響を与える。化学的調節因子のうち動脈血のO2 分圧,CO2 分圧,pHは頸動脈小体,大動脈小体の化学受容器を介して,髄液のpHの変化は延髄腹面の中枢性化学受容性領野の関与によって呼吸運動に影響し,血中の性ホルモンも影響する。しゃっくり は横隔膜の痙攣(けいれん)性収縮によって起こる発作的呼吸運動であり,声門が閉じているので奇妙な発声と特有な感覚が生ずるが,呼吸調節系の異常によって起こると推測されてはいるものの,発現のメカニズムは不明である。
身体の運動を行うとき,運動開始とともに換気量は著しく増加する。このときの肺胞気のCO2 分圧の上昇は軽く,身体運動による呼吸運動の促進と換気量増加を説明することはできない。O2 分圧,CO2 分圧以外の原因として種々の因子が考えられているが,呼吸促進がきわめて速やかに現れること,受動的に運動させた場合にも起こることから,神経性因子の働きが有力視されている。すなわち大脳皮質の運動野から脊髄へ送られる遠心性インパルスが,脳幹部の呼吸中枢に促進的に影響し,また屈伸によって四肢の関節の受容器から求心性インパルスが呼吸中枢に伝えられて,呼吸運動の促進,換気量の増加が起こると考えられている。血液 →呼吸中枢 →肺 福原 武彦
呼吸の異常 すでに述べたように,健康人の呼吸数は安静時1分間に平均11.7回,1回の呼吸量は約500ml で,リズム,深さともほぼ一様に保たれ,通常は自覚されないが,このような正常な呼吸運動は,上記の呼吸調節機構の異常をはじめいろいろの原因によって,さまざまの異常をきたすことがある。呼吸の深さ,リズムなどの変化は,呼吸中枢の障害のほか,大脳・脳幹の障害,呼吸筋の萎縮・麻痺,胸郭・肺の弾性・粘性の変化,肺のガス交換機能障害,動脈血組成に影響を与える糖尿病性アシドーシス,腎機能障害など多くの原因によって起こる。一般に,呼吸リズムの乱れは中枢神経の障害,浅く速い呼吸は肺・胸郭の硬くなる変化,遅い呼吸は気道の閉塞性障害,大きく速い呼吸は動脈血組成の異常による場合が多い。
(1)回数,深さの異常 頻呼吸は呼吸数の増加した場合を指し,一般に呼吸の深さは浅くなる。正常人での運動後に見られるほか,肺繊維症,僧帽弁狭窄症 による肺鬱血(はいうつけつ)などの拘束性換気障害において認められる。これは,肺が硬くて伸びにくくなるために,ヘーリング=ブロイエル反射効果の変化や呼吸筋内受容器の刺激で速く浅い呼吸になるものと思われる。毎回の呼吸量が異常に増加した状態が呼吸亢進であって,神経症患者に多く認められる。徐呼吸は呼吸数が病的に減少した場合を指す。クスマウル大呼吸は持続的に著しい呼吸亢進と徐呼吸が共存する場合で,糖尿病,尿毒症,コレラの昏睡時などに多く認められ,生体の代謝性アシドーシス に対する反応と解される。呼吸低下は毎回の呼吸量が減少した場合で,睡眠時にしばしば認められるほか,呼吸筋麻痺で頻呼吸とともに認められることが多い。
(2)リズムの異常 呼吸の深さ,回数が一定でなく,周期的に変化するので,一般に周期呼吸と呼ぶ。チェーン=ストークス呼吸は,浅い呼吸からしだいに深い呼吸となり,再び浅くなって,15~60秒の無呼吸期に移行するという周期を,比較的規則的に繰り返す呼吸である。大脳半球から間脳にかけての障害による意識障害,心臓や肺の病気の重症末期に多く認められるが,最近では老年者の睡眠中にも比較的しばしば出現することがわかってきた。原因に関してはまだ定説はないが,呼吸中枢の血中二酸化炭素レベルに対する感受性の低下あるいは感度の不安定化,血中二酸化炭素レベルを決定するうえで主役となる肺と,それを感知する呼吸中枢間の血液循環時間の遅れなどが関係しているといわれる。ビオー呼吸は失調性呼吸とも呼ばれ,チェーン=ストークス呼吸のように深さの周期的変化はなく,急激な短い呼吸の間に,持続時間の一定しない停止期のある呼吸で,主として髄膜炎において見られ,呼吸中枢の延髄レベルでの脳障害に関連するといわれる。ため息呼吸は,正常な呼吸が,ときどき深い吸気とこれに続く長い呼気で中断されるもので,神経症や神経循環無力症 に多く認められる。
(3)外観の異常 口すぼめ呼吸は,呼気に際して口をすぼめて気道内圧をあげ,気道閉塞を軽減しようと無意識に行っている呼吸で,進行した慢性肺気腫患者に見られる。鼻翼呼吸は呼吸困難のある場合に見られるもので,鼻翼が呼吸に伴って動く場合をいう。細気管支炎,肺炎,鬱血性心不全,自然気胸などに多く見られる。ただし,子どもでは興奮すると呼吸時に鼻翼が運動するので,これだけでは呼吸障害の症状とはいえない。起座呼吸は,寝ていると呼吸困難が強いため起き上がってしまっている状態で,急激に起こった肺鬱血の症状とされてきたが,気管支喘息(ぜんそく)や慢性肺気腫でも見られることがある。下顎呼吸は,吸息時に下あごが上方に上がり,呼息とともにゆるやかに下がるもので,呼吸はしばしば不規則となる。瀕死の重症患者に見られる。喉性呼吸は同様に瀕死または昏睡の患者に見られ,かたわらにいる人にも聞こえるほど,ごろごろまたはぜいぜいいう音を伴う呼吸である。咳反射が不十分なため,気管や気管支に痰が貯留し,その中を空気が通るために起こる。息ぎれ →呼吸機能 →呼吸機能検査 →呼吸困難 白石 透
呼吸器官respiratory organ 外呼吸を主要な機能とする器官。植物では呼吸根のような特殊化したものがあるが,一般には通気組織と気孔 が空気の通路となっており,とくに呼吸器官と呼べるものはない。
無脊椎動物の呼吸器官 無脊椎動物で分化した呼吸器官にはえら,気管,肺があるが,特別の呼吸器官をもたないものも多く(腔腸動物,扁形動物,袋形動物,触手動物,棘皮(きよくひ)動物など),これらでは体外,体内の表皮を通して呼吸が行われ,とくに触手などの薄くなった表皮に集中する。軟体動物のえらは,基本的には体側壁が外套腔(がいとうこう)内に突出したくしえら(櫛鰓)で,入鰓(にゆうさい)血管と出鰓血管が通る扁平な軸部に薄いえら板(鰓板(さいばん))がくしの歯状に列生して呼吸上皮を形成している。えら板には繊毛があって,水流を起こして呼吸のための換水をするが,頭足類では外套筋の運動で換水する。多板類,単板類ではくしえらは多対あるが,腹足類では体が巻くため片側では消失し,あるいは全部消失して二次的に外套膜の一部が突起してえらとなり,陸生のものでは外套腔壁が肺となる。二枚貝類のえら板は,糸状になって折れ曲がり結合したりして,複雑な構造となっている。多毛類では,いぼ足にある背触鬚(はいしよくしゆ)あるいは腹触鬚が葉状,糸状,樹枝状などに変形して,えらを形成している。羽状触手は呼吸上皮としての役割を果たすが,えらではない。節足動物では,付属肢の付属突起が変形してえらを形成している。剣尾類のえらは書鰓といわれ,腹肢の後面に列生している100~150枚の葉状突起で構成されている。甲殻類のえらには簡単な板状のものもあるが,十脚類の分化したものでは,葉状,糸状,樹枝状の鰓条が列生した構造となり,胸肢の基部に形成されるが,その部位,数は種類により多様である。一般に頭胸甲の側甲で覆われた鰓室内に保護され,付属肢あるいはその特化した部分を動かして換水する。口脚類や等脚類では腹肢の一部分がえらの役割をするよう変形している。橈脚類(じようきやくるい)などの小型の群では特別の呼吸器官はない。昆虫類の水生幼虫には,主として腹部から突出した羽状,総状のえらがあり,血管のかわりに気管小枝が入り込んでいるのが特徴的である。トンボ類幼虫の直腸鰓は,直腸壁に形成された微細なひだで,内部には気管小枝が密に分布しており,肛門を通して換水される。半索動物のえらは咽頭にあいた1~多対の鰓裂で,消化管内と体外とを連絡しており,基本的には脊椎動物の鰓裂と同じ構造のものである。ホヤ類では鰓裂は細分して多数の小孔となり,咽頭は繊毛を密生した網目状の鰓囊となって囲鰓腔の中に包み込まれ,出水口で体外に通じる。気管は,体表皮の一部が体内におち込んで呼吸上皮を体組織中に入り込ませている構造物で,主として陸生の昆虫類,多足類,蛛形類(ちゆけいるい),有爪類(ゆうそうるい)に発達している。軟体動物有肺類にある肺といわれるものは,外套腔体壁に毛細血管が分岐,結合しながら密集して呼吸上皮を形成しているものである。外套腔は呼吸孔をのこして薄い隔壁で閉じられて呼吸気室となり,水分の蒸発を防ぐとともにその下面の筋肉を上下させて換気する。ナマコ類には,直腸から体腔内に膨出した樹状の薄膜管で作られた呼吸樹(水肺)と呼ばれる器官があり,呼吸に用いられる。肛門と直腸の運動によってその管内を換水する。原田 英司
脊椎動物の呼吸器官 脊椎動物では,外呼吸に関与する器官には口,鼻,咽頭,喉頭,鰓裂,皮膚,腸管壁などもあるが,これらは呼吸以外に重要な機能も備えているので,普通は呼吸器官には数えない。水呼吸を行う水生脊椎動物の呼吸器官はえらであり,空気呼吸をする動物のそれは肺とその付属器官である。えらはほぼ樹状またはくし状の器官で,その外表面が呼吸作用をもつのに対し,肺は袋状の構造でその内表面が呼吸機能をもつ。また発生学的には,えらは咽頭の側壁に現れる内臓弓(鰓弓)の表面から発生するのに対して,肺は内臓弓の後方の消化管の膨出から生ずるもので,構造的にも発生的にもまったく別個の器官である。高等硬骨魚類だった古生代の総鰭類(そうきるい)はえらで水呼吸をしていたが,それと同時に肺を備え,口腔へ貫通した鼻孔によって空気呼吸をも行うようになっていた(現存の総鰭類であるシーラカンス類は空気呼吸はせず,もっぱらえらによる水呼吸をする)。その直接の子孫である両生類では,幼生(オタマジャクシ)はえら(カエルは内鰓,イモリとサンショウウオは外鰓)で水呼吸をするが,変態して両生化するにつれてえらは退化していき,それに代わって肺が発生し,やがて肺呼吸に頼るようになる。もっとも,えらが消失しても肺は発生せず,外呼吸は皮膚だけに頼るもの(ハコネサンショウウオ,アメリカサンショウウオ)や,天然状態では生殖可能になりながらもえらを終生維持し(この現象をネオテニーという),水呼吸を続けるもの(エゾサンショウウオ,メキシコサンショウウオ/別名アホロートル)が知られている。現生の肺魚類は水呼吸のためのえらと空気呼吸のための肺をともに備えている。比較発生学的に知られるところでは,硬骨魚類のうきぶくろは肺と相同の器官である。また古生物学的な証拠によれば,空気呼吸をする水生動物にはじめに肺が現れ,次いで水呼吸をする魚類でこれが浮力調節器官であるうきぶくろへ変形したと考えられている。したがって肺とうきぶくろを兼備する動物はいない。
えらと肺とに共通する点が二つあり,その一つはガス交換という生理的機能である。他の一つはえらの外面,肺の内面は総面積がひじょうに大きく,その表面はきわめて薄い上皮に覆われ,そこに毛細血管が豊富に分布しているという形態的共通点である。血液を通じてのガス交換という共通の機能はこの共通の構造によって達成される。鰾 (うきぶくろ) →鰓 (えら) →肺 田隅 本生
呼吸法の文化史 呼吸は本来,呼息と吸息にもとづく生理現象であるが,古くギリシアやインド,中国では,宇宙の大気と身体内部の生命の息(生命力)とを媒介する機能をもつとされた。この考え方はその後,とくにインドや中国で大きな発展をみせた。インドの古代哲学(ベーダーンタ学派)では呼吸(プラーナprāṇa)にかかわる機能を呼気,吸気,等気,上気,媒気の5種に分類したが(五風説または五気説),ヨーガ学派では,呼気→呼吸の停止→吸気という3種の機能からなる呼吸法(調息,プラーナーヤーマprāṇāyāma)の実修が説かれた。これは中間における呼吸の停止期間を長くすることに意を用い,独自の坐法と並行して宇宙との合一を目ざすヨーガ瞑想(めいそう)の基本とされた。このヨーガ の呼吸法はその後仏教にも取り入れられ,そのテキストの一つが後漢のころ安世高によって《大安般守意経》として漢訳されたが,〈安般〉とはサンスクリットのāna-apānaを音写した安那般那(阿那波那)を縮めたもので呼吸の意である。これよりさき,中国では戦国時代に身体運動を伴う呼吸術である導引 が行われていた。また道教が成立すると胎息 という独自の呼吸法が説かれたが,これは胎児が母親の腹中で行う呼吸の方法とされ,不死の生命を得るための最終的な手段とされた。また道教では,吸気を下丹田に集めて精と結びつける還精という呼吸法が説かれているが,これは仏教の禅定で重視される丹田呼吸とも関連している。このように中国では,呼吸法をめぐってヨーガ,仏教,道教の相互交流の跡がうかがえるが,その一例として天台智顗(ちぎ)の《摩訶止観(まかしかん)》をあげることができる。この著作の〈病患境〉という章には,座禅瞑想中に病気になったときそれを治すための対症療法的な呼吸法が細説されている。《摩訶止観》は日本の仏教に大きな影響を与えた論書であるが,そこに説かれている呼吸法は無視され,日本ではインドや中国の場合におけるように,呼気や吸気が宇宙と身体を循環するという観念は発達しなかった。《日本書紀》には,伊弉諾尊(いざなきのみこと)が息を吹くとそれが級長戸辺命(しなとべのみこと)という神になったとする伝承が語られているが,これはあくまでも風神の生成論であって,呼吸を媒介とする宇宙論がそこから生みだされることはなかった。息 山折 哲雄






 鑒伝〕今此の
鑒伝〕今此の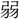 力を以て、彼の強寇に
力を以て、彼の強寇に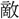 す。
す。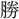
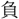 を一
を一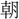 に決し、
に決し、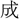 敗を呼
敗を呼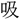 に定む。
に定む。