目次 原核細胞と真核細胞 細胞の構造と機能 細胞内膜系 細胞骨格 細胞の表面構造 細胞間のコミュニケーション 人為的細胞系 生命の基本的単位。細胞は細胞膜によって包まれた原形質 で,原則として自己増殖ができ,生体の構造単位でもある。細胞についての最初の記載は,R.フックによってなされた(1665)とされているが,それはコルク組織 の死んだ細胞のあとの細胞壁で囲まれた小さな部屋cellを観察したものであった。19世紀中ごろの細胞説の確立によって,生体機能を営むのは仕切られた小部屋を満たしていた原形質であり,すべての生体は原形質を含む細胞からでき上がっていることがわかってきた。20世紀中ごろから電子顕微鏡 による観察技術が急速に進歩して,細胞内部の微細構造が詳細に捕らえられるようになり,また,同じころに超遠心分離機 を利用する細胞分画法 が確立されたこととあいまって,生細胞から分離された各種細胞小器官の微細構造と,その代謝機能の局在性とが直接関連づけられるようになった。
現在,いろいろな細胞の微細構造ならびにその代謝機能が明らかにされるに及んで,生物の違いや細胞の違いを超えた共通普遍性を基盤に,細胞の特異性を理解し,また,研究する細胞生物学cell biologyが大きな発展を遂げている。
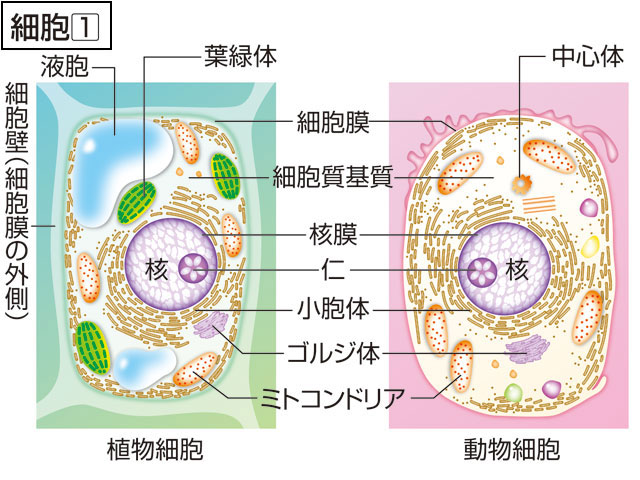
原核細胞と真核細胞 細胞には,原則的に1個の核様体nucleoid,あるいは核 nucleusがあって,その生物種に固有の遺伝子(DNA)のすべてがそこに局在している。すべての細胞は,核様体をもつ〈原核細胞prokaryotic cell〉と核をもつ〈真核細胞eukaryotic cell〉の二つのグループに分けられる。この両者の違いは,核をもつもたないにとどまらず,多くの著しい相違があって,両者の中間にあるような細胞は見あたらない。明らかに原始的とされる原核細胞は,細胞の大きさが1μm程度で小さく,通常細胞壁の形成によって決まる単純な形態をとっており,一部を除いて原形質にはとくに発達した構造が見あたらない。原核細胞と名付けられるとおり,核膜によって囲まれる原形質の区分(核質と呼ぶ)がなく,遺伝子を配列する染色体 は,ほとんどDNAの1分子と考えられ,核様体と呼ばれる構造をとって原形質中に広がっている。原始的な細胞生物である細菌類は,すべて原核細胞に属しているが,なかでも最小の細胞といわれる球形のマイコプラズマ は直径0.1~0.25μmの範囲にあり,細胞壁はなく,細胞膜のみによって囲まれる原形質には微細構造がきわめて乏しく,最小の染色体(長さ500μmにも達しない環状DNA)と最少数のリボソーム が含まれるにすぎない。
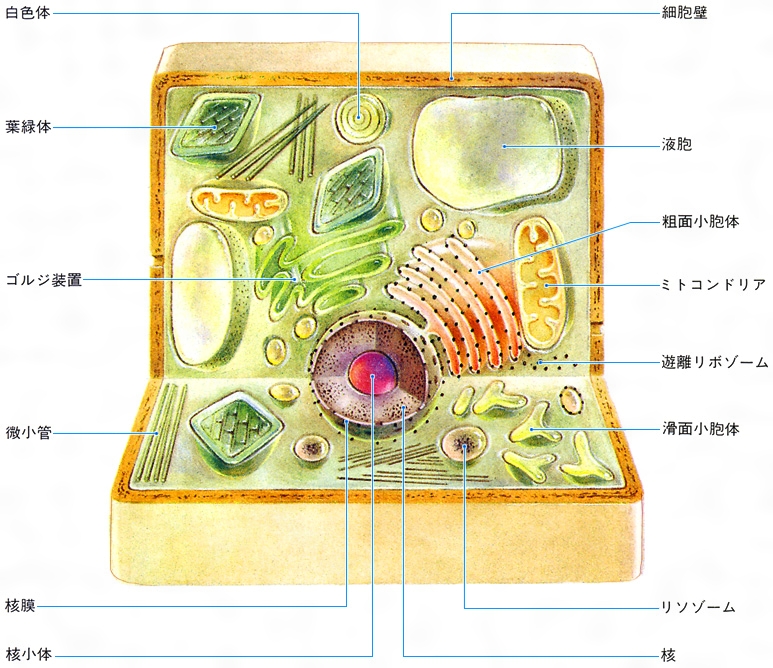
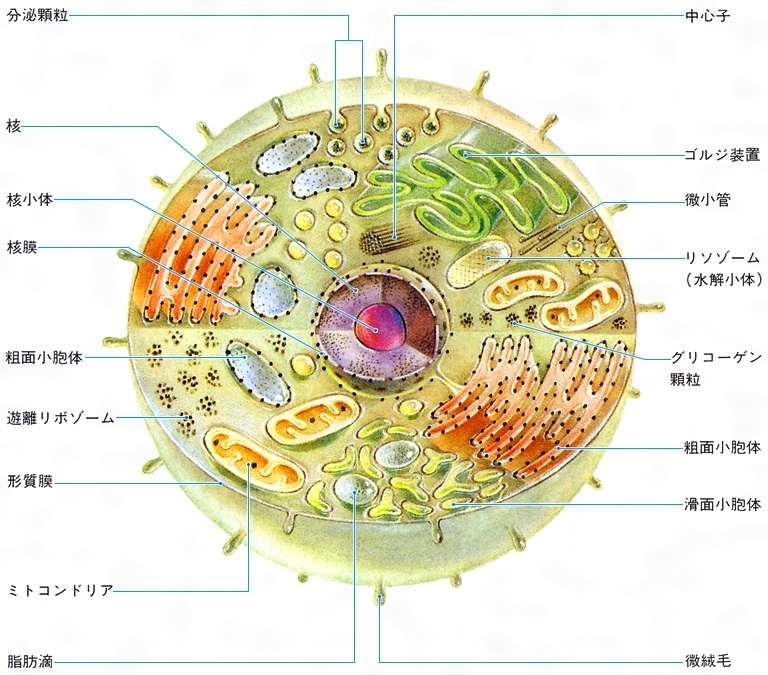
真核細胞は,生物種や細胞種によっても異なるが,通常原核細胞に比べて体積で3桁以上は大きく,ヒトの細胞の体積は200~1万5000μm3 の範囲にある。そもそも真核細胞の特徴は,原形質が二重の核膜によって囲まれる核質とその外側にある細胞質とに区分されていることである。核は,原核細胞に比べて100~1000倍も大きなゲノムDNAが結合する多数の染色体を保護し,遺伝情報の発現につごうのよい環境を形づくっている。核質のうち,染色体DNAを結合する染色質chromatinは,とくに遺伝情報発現とその調節を行っている重要な部分で,原核細胞の核様体とは違って,遺伝子DNAがヒストンというタンパク質8分子でつくる〈ヌクレオソームコアnucleosome core〉に巻きとられたビーズ状の構造を基本構造にして,スーパーコイルなどの高次な折りたたみ構造をとる染色糸がそこに位置している。また,核質には,RNAに富み,容易にそれとわかるボール状の核小体が通常1ないし数個分布しており,細胞質のリボソームに含まれる3種類のRNA分子(18S,5.8Sおよび28 Sr RNA)はつながった前駆体RNAとしてここで転写されている。真核細胞の細胞分裂 は,原核細胞が最も簡単な2分裂によるのとは違って,核分裂と細胞質分裂とが区別される。核分裂の際には,複製したDNAをもつ染色分体chromatidの分離のために,光学顕微鏡で容易に観察されるコンパクトな染色体構造と紡錘糸がつくる分裂装置によって,遺伝子セットは両娘細胞(じようさいぼう)核に正確に分配される。続いて細胞質を二分する細胞質分裂が起こるのである。なお,真核細胞の分裂像は,染色体の挙動,動原体と紡錘糸の結合,紡錘体の極と中心体,核膜の状態など,複雑で多様なものであるが,真核細胞の系統進化を知る重要な手がかりになっている。核が傷つくことは細胞の死につながるが,細胞質は細胞の生活活性の場であり,核があれば細胞質は損なわれても残りの細胞質の物質代謝によって再生や生長が行われる。また,真核細胞の著しい特徴は,細胞質の各種代謝機能が細胞質の部分構造と結びついて細胞小器官 となり,分業化によって効率的に行われていることである。細胞小器官としてミトコンドリア ,小胞体膜系と各種小胞ゴルジ体 などがあり,植物細胞には葉緑体 や色素体 ,また,しばしば大きな液胞 が発達していることなどは,原核細胞との大きな相違である。この相違は,真核細胞の起源を問題にするとき,説明されなければならない。例えばミトコンドリアは好気的細菌を先祖とし,葉緑体はラン藻を先祖として,始原真核細胞の細胞質へ共生した結果であると説明しようとする内共生説endosymbiotic theory はその仮説の一つである。真核細胞の細胞小器官のすべてが,膜分化によって直接に始原細胞の細胞質中に分化してきたとする膜分化説の立場もある。
原核細胞は原核生物として単独で生育しているが,特殊に分化した真核細胞である原生生物も単細胞生物 である。藻類の遊走子・配偶子,動物の卵細胞・精子のように細胞が遊離細胞として独立に機能している細胞もある。また,多核体や変形体のように通常の真核細胞とは著しく違う多細胞的生物やボルボックス のように多数の個体が集まって群体を形づくるものもある。多細胞生物の場合は,1個の卵細胞の受精によって発生を開始し,細胞分裂によって細胞数が増加して,それぞれの細胞が構造と機能の分化した組織を構成する組織細胞になる。組織細胞は種固有の構築にしたがって決められた位置関係をとっている。しかし,どの組織細胞もその生物種の完全なゲノムDNAをもっている等価な細胞と考えられる。したがって分化した細胞の特異な構造と機能は,特定の構造と機能を支配する遺伝子群の情報発現が特定の細胞で行われるように,細胞分化 の機構が働くことによって実現される。通常,細胞の分化は不可逆的で決まった方向に起こるように見えるが,ゲノムDNAのすべてを細胞分化ののちにももち続けているので特定の条件が与えられれば,別の細胞へ分化しなおすことも可能なはずである。実際に,ニンジンの1組織細胞を適当な条件下で培養し続けると,ニンジンの植物体ができ上がることがこの典型的な証明とされている。なお,組織細胞のように分化した細胞の増殖は無限には続けられることがなく,したがって組織の大きさに限度がある。癌細胞は,分化した細胞とは違い,組織細胞のような行動をとらず,ほとんどかってに分裂し続ける。また,組織細胞は,細胞分裂によって絶えず再生されてはいるが,細胞分裂を繰り返しているうちに,この周期からそれて分裂が止まり,ついには細胞死に至る細胞の老化 の過程が見られる。一方,原核細胞は,胞子形成のような特殊な場合を除いて,細胞分化にあたるものも,老化にあたるものも見られない細胞である。
細胞の構造と機能 真核細胞は生物種を超えて共通な細胞小器官としてミトコンドリア,小胞体,ゴルジ体などを含むが,分化した細胞には,このほかにその分化した機能を発揮するための細胞表層や内部形質に特異的な微細構造が発達する場合が多い。筋細胞に発達する筋原繊維と筋原繊維に興奮を伝達する筋小胞体とトリアドtriad,神経細胞の軸索突起や樹状突起および多数のシナプス,腸上皮細胞の微細絨毛(じゆうもう),精子の先体acrosome構造と鞭毛構造,色素細胞のメラノソームmelanosomeなどはその例である。
細胞の物質交代は,酵素反応によって行われ,化学エネルギーを効率的に利用するために,きわめて巧妙に制御されている。その鍵を握る多くのアロステリック酵素 は基質以外の作用物質によって代謝系ごとに,また,代謝系の間で,細胞環境に適応したエネルギー代謝の調節を行っている。糖の代謝やアミノ酸の代謝は細胞質の可溶性酵素系で行い,タンパク質合成はリボソーム上で,脂質の代謝は主として小胞体膜系とミトコンドリアで行う。生体物質の合成に必要なエネルギーはATPによって与えられるが,大部分のATPはミトコンドリアの酸化的リン酸化と,植物では光合成 における葉緑体の光化学反応によって合成される。このATP合成は,いずれも化学浸透chemiosmosisの機構を利用したもので,ミトコンドリアや葉緑体の内膜の内外に電子伝達系と共役してプロトンの濃度こう配をつくり,そのポテンシャルエネルギー が,共役因子(ATPアーゼ )の働きによってATPの合成に使われている。生物はこの機構によって解糖系の酵素によるATP合成よりも多量なATPを合成している。
真核細胞は,原核細胞と同様に原形質の可溶性分画によって細胞の基本的な代謝機能を果たしているが,そのほかにさらに高次な構造をもち,高次な制御系をもっている。とくに,細胞質に発達した異質の構造系として細胞内膜系と細胞骨格構造とがあり,高次な構造をつくり上げ,高次な機能を発揮できるのはこれらの構造のためである。
細胞内膜系 網目状に発達した複雑な膜構造で,分化した細胞の機能と密接な関係があり,その発達のようすは細胞の種類によってひじょうに違っている。細胞内膜系は全体として緊密な連絡を保ってはいるが,部分によってある程度の独自性をもつコンパートメント を形づくる。代謝系の酵素群は一様な分布をせず,効率のよい分業と流れ作業の工程ができ上がっている。このような膜系の構造と機能は,エクソサイトーシスexocytosis(細胞外への物質の放出)やエンドサイトーシス endocytosis(細胞内への物質の取り込み)の場合によく調べられており,GERL系(ゴルジ体Golgi apparatus→小胞体endoplasmic reticulum→リソソームlysosome)によってよく表されている。GERL系の各部分,粗面小胞体膜と滑面小胞体膜,ゴルジ体の膜系と小胞,食作用にともなうリソソームと食胞,分泌過程における分泌顆粒(かりゆう)と細胞膜の融合など一連のダイナミックな膜流動を行い,絶えず膜系の新生と再生をともなっている。GERL系は,原核細胞に比べて大きな真核細胞の細胞質が細胞外液との間の物質交換の機能を高めるために,細胞質中に網目状に発達させた運河系と考えられる。とくに,真核細胞の核は,内外二重の核膜によって取り囲まれており,その内腔は小胞体系を経て細胞外へ通じているから,細胞外の環境からの直接の影響を受け入れることができるようになっている。また,膜系自体は,親油性,疎水性物質の代謝系を結合しており,脂質,とくにリン脂質,コレステロール ,ステロイドホルモン ,プロスタグランジン ,ロイコトリエン などの合成の場所になっている。
細胞骨格 動物細胞の細胞内繊維構造は,細胞内膜構造とともに細胞の構造と機能を支える2本の主柱といえる。植物細胞では厚い細胞壁によって,外側から細胞形態のほとんどが決められてしまうので,細胞の骨格構造の発達を必要としない。これに対して,動物細胞は,広い自由表面を包む細胞膜自体に大きな細胞の形態を保つ働きがない。細胞質に網目状に発達した繊維構造は骨格として働き,また,細胞形態を変えたり,細胞運動に必要な張力圧力の伝達を行っている。多くの細胞で,核が細胞の中心に位置するように,細胞の繊維構造が核を係留し,細胞小器官やその他の微細構造の配分とその変化に対しても重要な役割を果たしている。さらに繊維構造は,デスモソーム desmosomeを介して細胞間の結合にも関係しており,細胞集団の形態形成は,各細胞内の繊維構造の再編成とその繊維構造による力の発生および伝達で実現される。
このように多方面にわたる細胞機能に関係している細胞骨格cytoskeltonは,複雑で細胞種によっても微妙に違うが,それぞれ特色ある3グループの繊維(微小管,中間繊維,マイクロフィラメント )によってつくられる構造に大別される。これらの繊維構造は,それぞれの構成タンパク質を抗原として調製した蛍光抗体で,細胞を染めれば,蛍光顕微鏡によってこれらの細胞骨格構造を識別することができる。
(1)微小管microtubule 細胞骨格の中で最も太い250nmの中空の繊維構造で,おもな微小管タンパク質としてチューブリン α,βが重合してできたものである。そのほかに,微小管の重合や他の繊維構造との間の相互作用に必要なτタンパク質や微小管結合タンパク質(MAP)など少量のタンパク質がこの構造に加わっている。真核細胞の分裂には,微小管が紡錘体の両極に位置する中心粒から発する紡錘糸として,染色体の動原体kinetochoreに結合し,染色体分離の役割を果たす。分裂間期に入ると,微小管は核に近い細胞質に位置している微小管形成中心(microtubule-organizing centre=MOC)から発して,放射状に細胞周縁に向かって走る。また,同時に細胞核を網目の竹籠のように包み込み,それを宙づりにするような骨格構造に発達する。微小管構造はこのように細胞周期に連動した細胞形態に関係している。また,広く真核細胞に見られる鞭毛や繊毛は微小管が構成繊維となっており,その根もとにある基粒体basal bodyは中心粒と同じ構造をもつ自己増殖系である。内共生説の立場からすると,基粒体をもつ鞭毛や繊毛と中心粒はもともとスピロヘータ が始原真核細胞へ共生したものであるという仮説に発展する。ある種のスピロヘータには,チューブリン と同種の抗原タンパク質が含まれ,細胞質を貫く繊維構造はこれが構成タンパク質になっているので,このような仮説を支持する証拠にあげられている。
(2)マイクロフィラメントmicrofilament 直径50nmほどの細い繊維構造で,Fアクチンが構成タンパク質である。筋細胞では筋原繊維の細繊維がα-アクチンの重合によってできており,これと平行に走る太いミオシン 繊維との間の相互作用によって力を発生し,筋収縮が起こる。広く非筋細胞にも同族タンパク質のβ-アクチンおよびγ-アクチンが分布していて,マイクロフィラメントを形づくって細胞のダイナミックな形態変化を引き起こしている。マイクロフィラメントのあるものは,ミオシンと結合してストレス・ファイバーstress fiberと呼ばれる細胞質を直線的に走る太い繊維として分布するが,多数のマイクロフィラメントは細胞の伸長方向に平行に走る細繊維として,また,細胞表層に沿って平行に走る繊維束として分布する。細胞の形態変化や運動にとって重要なことは,細胞膜にも結合点をもつことである。アクチン繊維 の特色は,分化した細胞形態や機能にかかわって,それぞれ異なった繊維構造の成分タンパク質と相互作用することであり,その状況によって機能の仕方が違ってくることである。
細胞表層の構造について見ると,赤血球では,スペクトリンspectrinが細胞表層にあってアクチン繊維と相互作用し,赤血球以外では,ホドリンfodrin,α-アクチニンα-actinin,ビンキュリンvinculinがあってアクチン繊維と相互作用する。また,これらの相互作用には細胞膜と細胞表層のアクチン繊維の結合が関係する。一般に,細胞表層が変化して形づくられる微細絨毛,マイクロスパイク,皺状(しゆうじよう)突起,細胞分裂に見られる収縮環などいずれもアクチン繊維の配列の変化によって起こる現象である。癌化した細胞が丸くなるような形質転換には,細胞膜と細胞骨格の繊維タンパク質の間で相互作用が変化しているらしい。
(3)中間繊維intermediate fiber 微小管とマイクロフィラメントの中間の太さに当たる100nmほどの繊維構造を総称したもの。おもに細胞の形態保持,細胞形態のゆがみにかかわり,張力,弾力が細胞間接着構造(デスモソーム)の中間繊維を介して細胞内および細胞間に伝播(でんぱ)すると考えられている。例えば,表皮組織に加えられる各方向の張力,弾力の伝播はその例である。
中間繊維の構成タンパク質は,細胞種によって異なり,それぞれの細胞構造や機能の特異性に関係していると思われる。上皮細胞にはサイトケラチンcytokeratin,間充織細胞その他の細胞にはビメンチンvimentin,筋細胞にはデスミンdesmin,神経細胞にはニューロフィラメントタンパク質neurofilament protein,グリア細胞にはグリア細胞特異的フィラメントタンパク質があって,それぞれの細胞種における中間繊維を形づくっている。中間繊維の網目状構造は,まず核を取り囲む籠状の構造を形づくり,一方では,細胞膜上の付着点に発し,離れた細胞膜上の付着点に至るかすがい状に走る網目状の構造を形づくっている。細胞膜上の付着点は隣接細胞間の接着斑にあり,中間繊維はトノフィラメントtonofilamentと呼ばれているものである。
なお,細胞骨格をつくる3種の繊維構造の間にも相互作用があり,細胞全体の構造ならびに機能に対して調節し合っていると考えられる。
細胞の表面構造 細胞外液に面している細胞膜表面には,糖鎖が突き出してできる親水性の層glycocalyxがある。糖鎖は各種の単糖の配列の違いによって,生物種や細胞種の違いをよく表している。動物細胞は,細胞が合成し分泌したタンパク質であるコラーゲンcollagen,フィブロネクチン fibronectin,ラミンlaminなどによって覆われ,また,これらのタンパク質は細胞膜と相互作用することによって細胞の形態や機能に影響を与えている。ラミンはコネクチンconnectinを介して細胞表層のアクチン繊維と結合でき,フィブロネクチンは細胞膜を貫く結合タンパク質を介してアクチン繊維と結合でき,結果として,細胞骨格と連動しうる構造になっている。癌化に際して,細胞形態の変化と同時に,細胞の外側を包み込む形で発達しているフィブロネクチンの網目構造が著しく減少することが知られているが,この現象は細胞膜からアクチン繊維が離れ,ストレスファイバーが見えなくなるなど,アクチン繊維の分布の変化と関係がある。
細胞間のコミュニケーション 多細胞生物では,まとまりのある個体としては欠かせない細胞間のコミュニケーションがいろいろなレベルで行われている。隣接細胞の間には,ギャップ結合 と呼ばれる隣り合う細胞膜部分を貫いた多数の筒形タンパク質集合体がセットされている。筒形の中心は,隣り合う細胞の細胞質に通ずるトンネルになっており,分子量1000以下の分子は自由に通過できるので,このような低分子物質の往来が情報となりうる。また,この結合は電気的結合とよばれるように,低分子イオンによる通電性を高くしている。同一組織内の細胞間にはこのような結合が密に発達していて,細胞機能のグループ化に働いていると考えられる。
遠く離れた細胞間のコミュニケーションには,体液の循環によって運ばれるホルモンや栄養物質が,標的器官の細胞にキャッチされるしくみがあって,それぞれの物質に特異的な受容体receptorが細胞膜の外側に分布している。受容体が分布する細胞だけがその物質を情報として受け入れられるので,少量で多数の情報物質があっても個々の情報は整理され,的確にとどけられているのである。インシュリン ,β-メラノトロピン ,絨毛性性腺刺激ホルモン,神経成長因子(NGF),上皮細胞増殖促進因子(EGF),ソマトメジンCなどのペプチドホルモン ,また,栄養物質の輸送タンパク質として,低比重リポタンパク質 (LDL),卵黄タンパク質(ホスビチン,リポビテリン),ハプトグロビン ,トランスフェリン などは細胞膜上の特異的受容体に結合して,レセプトソームreceptosomeと呼ばれる細胞膜の陥入によってできた小胞とともに,細胞質中に取り込まれていく(この過程がエンドサイトーシスである)。この機構によれば10⁻8 ~10⁻1 0 molという低濃度の情報物質をも濃縮して標的細胞に伝えられる。
多細胞生物の場合は,直接に隣接する細胞間の物理的結合や情報の伝達だけではなく,拡散するが,化学的に特異性の高い情報物質と結合する受容体によって遠くの細胞間にコミュニケーションを成立させ,個体の統合を果たす機構を著しく発達させているのである。
人為的細胞系 遺伝子操作の技術が容易になったため,細胞に外来の遺伝子を導入して,その遺伝子が他の細胞の中でどのような働きをするかを調べる研究が,いろいろな細胞の組合せで試みられている。遺伝子DNAの化学的な性質は,原核細胞から真核細胞に至るまで,まったく同じであるから,情報としての機能も基本的に同一と考えられる。しかし,情報発現の調節機構については種によってかなり違うので,遺伝子の導入に際しては調節機構を支配するDNA部分を含んだDNAが用いられる。この種の研究の一つとして,癌遺伝子を導入して癌化の機構を解明するという実験が盛んになされつつあり,近い将来,その全容が明らかにされることが期待されている。また,細胞融合 法を用いて,違う種の間で自然界に存在しないような雑種細胞をつくり継代培養するという実験も行われている。ニンジンなどでは,単一培養細胞から完全な植物体をつくることが成功しているから,こうして生じた雑種細胞を培養することによって,全く新しい雑種植物を人為的につくりだすことが可能である。
しかし,遺伝子の組換え操作は,人類にとって有用な生物を人為的につくりだす一方で,自然の生物界を混乱させることになる可能性があり,慎重に行われるべきものである。腰原 英利