デジタル大辞泉 「遺伝子」の意味・読み・例文・類語
いでん‐し〔ヰデン‐〕【遺伝子】
2 「DNA2」に同じ。「名人と呼ばれた先代の
[類語]細胞・細胞膜・細胞壁・細胞質・原形質・単細胞・胚・核・核酸・リボ核酸・デオキシリボ核酸・染色体・性染色体・ミトコンドリア・組織・胚珠・胚乳・胚芽
Sponserd by ![]()
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
遺伝形質を規定する因子。遺伝子は、細胞内に存在する核酸分子、デオキシリボ核酸(DNA)の一部であり、自己複製により子孫に伝えられる。また、転写と翻訳によりタンパク質の構造を決定し、その働きにより遺伝形質を発現する。
[黒岩麻里 2023年10月18日]
チェコのブルノの町で修道院の司祭をしていたメンデルは、1865年にエンドウの交雑実験の結果をまとめて「植物雑種についての実験」という講演を行い、翌年にこの講演の内容が論文として学会紀要に掲載された。この論文では、エンドウのそれぞれの形質に対応する細胞内の物質的なエレメント(要素)の存在が記載されていた。そして1900年に、複数の研究者がそれぞれ独立した研究によりメンデルの研究を検証することで普遍的な遺伝的法則をみいだし、遺伝学誕生の基礎となった。なお、メンデルが遺伝の仕組みを明らかにしたが長らく注目を浴びることなく、のちに彼の法則が再発見されたというのが通説である。しかし、実際にメンデルはこの論文で遺伝の仕組みについては考察しておらず、論文中のエレメント(要素)は、現在明らかになっている遺伝子という分子の存在(微小粒子の存在)を想定したものではない、という説もある。ただし、メンデルの遺伝学への貢献が否定されるものではない。
そして、遺伝を担う物質が細胞内の微小粒子であるという考えは、C・R・ダーウィンをはじめとする複数の研究者により提唱された。デンマークの遺伝学者のヨハンセンは、微小粒子の正体は不明であるとしながらも、その粒子を「ゲン(Gen)」とよぶことを提唱した。これを語源として、のちに英語では 「Gene」、日本語では「遺伝子」とよばれるようになった。
また、アメリカの生物学者であるサットンWalter Stanborough Sutton(1877―1916)が、バッタの生殖細胞を用いて染色体を観察し、遺伝の様式を染色体の性質や挙動によって説明する「染色体説」を1902年に提唱した。こうして、遺伝子と染色体の関係が明らかにされてくるなか、アメリカの遺伝学者T・H・モーガンは主としてキイロショウジョウバエを用いた研究を行い、遺伝子は染色体上に線状に配列する粒子(物質)であることを明らかにした。
1940年代には、遺伝生化学や分子遺伝学研究が発展し、遺伝子は染色体をつくる核酸の一種であり、酵素分子の働きを支配して遺伝形質を決定することが明らかにされた。1960年代には、遺伝子のもつ遺伝暗号がすべて解読され、遺伝情報の発現機構が解明された。さらに1970年代には、遺伝子の人工合成が可能になり、また細胞から取り出した遺伝子を異種の細胞に入れて増殖させ利用する遺伝子工学技術も発展してきた。
[黒岩麻里 2023年10月18日]
遺伝子の本体は核酸の一種、デオキシリボ核酸(DNA)である。例外的にある種のウイルスではリボ核酸(RNA)が遺伝子として働く。
遺伝子がDNAであることが証明された科学的歴史は、1928年、イギリスの細菌学者グリフィスFrederick Griffith(1879―1941)による肺炎球菌を用いた実験に始まる。グリフィスは、肺炎球菌の細胞に含まれるなんらかの物質が、子孫に伝達される遺伝的な変化をもたらしていることを明らかにし、この現象を形質転換とよんだ。しかし、この時点ではその物質は何であるかはわかっておらず、当時、遺伝情報を運ぶのはタンパク質であると広く信じられていた。
そして1944年、アメリカの分子生物学者・細菌学者エーブリー(アベリー)、マクラウドColin Munro MacLeod(1909―1972)、マッカーティーMaclyn McCarty(1911―2005)の3人の実験により、タンパク質を分解した肺炎球菌は形質転換を起こすが、DNAを分解した肺炎球菌は形質転換を起こさないという結果が得られ、DNAが形質転換物質であると結論づけられた。
その後、1952年にアメリカの微生物学者・分子生物学者ハーシェイ(ハーシー)とチェイスがバクテリオファージT2の生活史の研究を行い、T2ファージの増殖に必要な遺伝情報をもつのはDNAであることを証明し、遺伝子がDNAの本体であることが確認された。
[黒岩麻里 2023年10月18日]
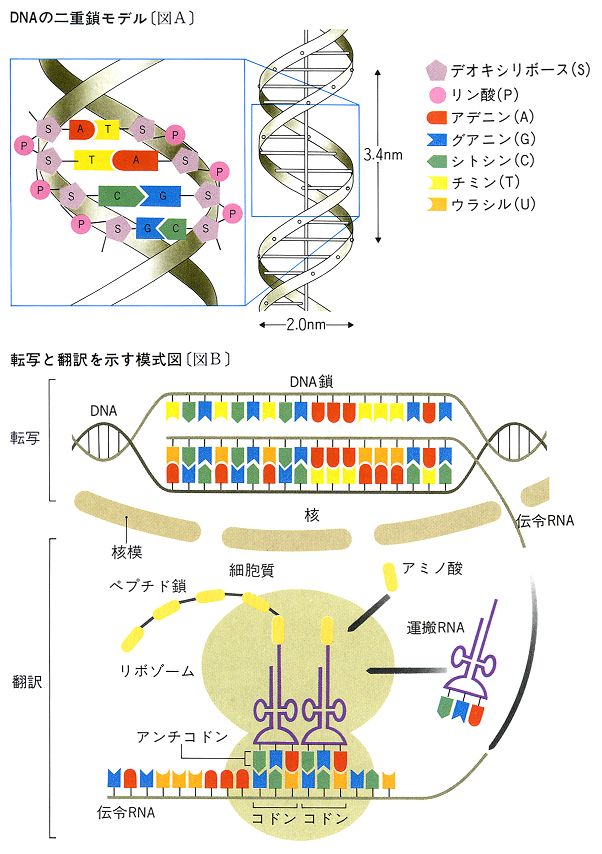
DNAは多数のデオキシリボヌクレオチドが結合してできた高分子物質である。デオキシリボヌクレオチドはリン酸、五炭糖のデオキシリボース、プリンまたはピリミジン塩基が結合したものである。DNAをつくる塩基は、アデニン(A)、グアニン(G)、シトシン(C)、チミン(T)の4種である。1950年、アメリカの生化学者シャルガフはDNA中のアデニンとチミンの数が等しく、シトシンとグアニンの数が等しいことを発見した。これらの結果は、アメリカの分子生物学者J・D・ワトソンとイギリスの物理学者・分子生物学者クリックに伝えられ、1953年に彼らがDNAの構造を発見する成果へとつながった。ワトソンとクリックが提唱したモデルは、DNA分子はデオキシリボヌクレオチドが結合してできた2本の長い鎖が螺旋(らせん)状に巻いた構造からなるというものである()。2本の鎖はAとT、GとCが塩基対をつくるように結び付き、二重螺旋構造をとっている。二重螺旋の直径は2.0ナノメートル、螺旋の1回転の距離は3.4ナノメートルで、その間に塩基対が10個並んでいる。このようなDNA鎖をつくる塩基の配列順序は遺伝暗号として働き、遺伝情報を決定するものである。DNAの量は種によって異なるが、遺伝学研究によく用いられる大腸菌の全DNA塩基配列は464万塩基対であり、約4000の遺伝子をもつと推定されている。
遺伝子は細胞分裂の際に正しく同じものに複製され、子孫細胞に伝えられる。DNA分子の複製の際には、二重螺旋構造は部分的に1本鎖となり、それぞれの鎖を鋳型として、DNA合成酵素の働きにより対になる新しい鎖が合成される。このようにDNA分子の複製は自己複製であり、また二重鎖の1本から新しい分子がつくられるので半保存的複製とよばれる。
[黒岩麻里 2023年10月18日]
高等動植物など真核生物の細胞では、DNAはヒストンタンパク質に巻きつけられたヌクレオソームnucleosomeという構造をとっている。多数のヌクレオソームが連なった構造をクロマチンchromatinとよび、染色体はこのクロマチンが折り畳まれて高次構造をとったものである。狭義には、細胞の分裂時にクロマチンが凝縮して棒状の構造をとったものを染色体とよぶが、広義には、その形や細胞の分裂周期にかかわらず、クロマチンの構造体を染色体とよぶ。細菌類など原核生物の細胞では、細胞分裂の際に真核細胞でみられるような核や染色体をつくらず、DNA分子はクロマチン構造をとることなく細胞質中に分布している。
遺伝子は染色体上に線状に配列している。二つの遺伝子が異なる染色体上にあるときには、次世代への継承はメンデルの独立の法則に従って分離するが、同じ染色体上にあるときにはこの法則に従わず、行動をともにし、連鎖の現象を示す。1本の染色体上の遺伝子は一つの連鎖群を形成するが、交雑の結果、連鎖している遺伝子の組合せが親と異なる組合せに変わることがあり、この現象は遺伝的組換えとよばれる。組換えは減数第1分裂の過程で対合した相同染色体の間で交差と乗換えが起こり、新しい遺伝子の組合せとなる組換え型が生じる現象である。組換え型の出現頻度をパーセント(%)で示した値を組換え価とよぶ。組換え価を遺伝子間の距離とし、これを線上に目盛ると、遺伝子が染色体上にどのように並んでいるかを示す図ができる。この図は染色体地図、遺伝地図、あるいは連鎖地図とよばれる。
組換えはDNA分子間の交差切断と、相同な相手分子への乗換えによって起こると考えられている。組換え過程では、DNA鎖の切断酵素、修復酵素、連結酵素などが働いている。DNA鎖の特定塩基配列部位を切る酵素は制限酵素とよばれる。同じ制限酵素で切った2種のDNA鎖は切り口の構造が相補的であり、それらを結合してできた分子は組換えDNAとよばれる。宿主(しゅくしゅ)細胞内で自己増殖する核外遺伝子プラスミドのDNAと目的とする特定の遺伝子DNAとの間で組換えDNAをつくり、それを宿主細胞に導入することで遺伝子のコピーを増やすことができる。これは遺伝子のクローン化とよばれる現象で、遺伝子工学の主要な手段となっている。
[黒岩麻里 2023年10月18日]
遺伝子は遺伝形質を決定する。アメリカの遺伝学者のビードルとテータムは1941年に特定の栄養分を必要とするアカパンカビの突然変異体の研究を行い、「一遺伝子一酵素説」を提唱した。この仮説は、遺伝子は一つの酵素の構造や働きを支配し、遺伝形質を発現するとするものである。その後、1958年にクリックが分子生物学の「セントラルドグマ」(中心教義)として主張しているように、遺伝子DNAの遺伝情報はまず伝令RNAに転写されて、細胞質に移動する()。伝令RNAはリボゾームとよばれる小粒に付着し、運搬RNAなどの働きで翻訳され、翻訳されたものから遺伝暗号に従ってアミノ酸が結合され、最終的に結合されたものからタンパク質のポリペプチド鎖が合成される()。タンパク質は酵素として細胞内の代謝反応を触媒し、また細胞構造をつくり遺伝形質を発現させる。
遺伝子のもつ遺伝暗号は、1961年から約5年間をかけて解読された。解読された遺伝暗号は、遺伝子DNAの三つのヌクレオチドがコドンcodonとよばれる暗号の単位となって一つのアミノ酸を指定するというもので()、トリプレット暗号ともいわれる。コドンは64種あり、そのうち61種はタンパク質をつくる20種のアミノ酸のどれかを指定するが、3種のコドンはどのアミノ酸も指定せず、遺伝暗号の読みの終止暗号として働く。また、必須アミノ酸メチオニンの暗号(AUG)は遺伝暗号の読みの開始暗号となる。ウイルスからヒトに至るまで、どの生物も同じ遺伝暗号を用いている。
遺伝子が働くか働かないかは、環境条件や遺伝子がもつ調節機能により制御されている。フランスの分子遺伝学者F・ジャコブと分子生物学者J・L・モノーは1961年に大腸菌の乳糖代謝の調節機構の研究から、酵素合成が調節遺伝子によりオペロンとよばれる遺伝子群を単位として調節されるという「オペロン説」を提出した。オペロン説によると、大腸菌の培養中に乳糖がないときには、調節遺伝子からつくられる調節物質(リプレッサーとよばれるタンパク質)がオペロンの一端(オペレーターとよばれる)に結合し酵素合成を止めているが、乳糖が加えられるとリプレッサーは不活性化され、オペロンから酵素が合成される。真核細胞は多数の遺伝子をもつが、組織に特有の遺伝子のみが働き、ほかは働きを停止している。このような真核細胞における遺伝子作用調節機構は、染色体構造と密接な関係をもつものと考えられている。
[黒岩麻里 2023年10月18日]
DNA複製の際のミス、動く遺伝子(転移因子)トランスポゾンの転移、物理的要因によるDNAの損傷などにより、DNA配列に起きる変化を突然変異とよぶ。物理的要因としては、放射線の照射や、アルキル化剤のようなDNAに作用する化学物質などが知られている。突然変異はDNA塩基対の置換、欠失、転座、逆位、挿入などにより、遺伝子配列上に変異が生じ、指定するアミノ酸配列が変わることによって起こる。突然変異で遺伝子機能の性質が変化することで、生存に有利に働いたり、逆に遺伝子機能が失われたりする。
[黒岩麻里 2023年10月18日]
遺伝子の名称は遺伝子記号で表される。遺伝子記号は、その遺伝子の決定する形質の特徴を示す英語やラテン語などの省略形と、番号や記号からつくられる。たとえば、微生物でアミノ酸の一種であるトリプトファンがないと生育できない形質を示す遺伝子の記号はtrpである。トリプトファン合成系にはいくつもの酵素反応があり、それぞれの反応を支配する遺伝子には番号やアルファベットをつけtrp1、trp2あるいはtrpA、trpBのようによぶ。DNAやRNAをさす場合はイタリック(斜体)、タンパク質をさすときはローマン(正体)文字とする。また、遺伝子とアレル(対立遺伝子)の記載は区別する必要があるが、混同されている場合も多い。遺伝子の記載方法は生物種によって異なるが、アレルの記載は、潜性アレルを小文字、顕性アレルを大文字で表す。
遺伝子の固有名とは別に、遺伝子が細胞のどの部分にあるか、どのような形質を支配するか、どのような働きをするかなどにより、種類分けがなされている。核にある遺伝子は核遺伝子あるいは染色体遺伝子とよばれる。細胞質にあって細胞質遺伝の原因となるものは細胞質遺伝子とよばれ、とくに細胞質中の細胞小器官であるミトコンドリアに存在するものはミトコンドリア遺伝子とよばれる。一遺伝子一酵素説に従いタンパク質の構造を決定しているのは構造遺伝子であり、構造遺伝子の働きを誘導したり抑制したりしているのが調節遺伝子である。また、生物個体の発生過程で致死作用を示す遺伝子は致死遺伝子とよばれる。自然界に普通にみられる正常な遺伝子は野生型遺伝子で、変化すると突然変異型遺伝子になる。また、一つの形質を発現するのに2種類以上の異なる遺伝子座にある遺伝子が働いていることがあり、これらの遺伝子は同義遺伝子とよばれる。これらのほかにも、各種の遺伝現象を表すために、多数の名称が記載されている。
[黒岩麻里 2023年10月18日]
『ホーグランド著、市場泰男訳『遺伝子のはなし』(1981・社会思想社)』▽『石川辰夫著『分子遺伝学入門』(岩波新書)』▽『ジェームス・D・ワトソン他著、中村桂子監訳、滋賀陽子他訳『ワトソン 遺伝子の分子生物学 第7版』(2017・東京電機大学出版局)』▽『ブルース・アルバーツ他著、中村桂子・松原謙一監訳、青山聖子他訳『細胞の分子生物学 第6版』(2017・ニュートンプレス)』▽『池内昌彦他監訳『キャンベル生物学 原書11版』(2018・丸善出版)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
遺伝形質を規定する因子。
G.J.メンデル(1865)はエンドウの子葉の色の緑と黄というような対立的な形質を支配する遺伝因子として対立する要素を想定し,両親由来のこのような対立要素,例えばAとa,をもつ雑種が配偶子を形成するとき,Aとaが分かれて別々の配偶子に入り,これが子どもに伝えられてその形質を規定すると考えた。1900年のメンデルの遺伝法則の再発見以降,多くの生物でこのような対立形質の遺伝様式が盛んに研究されるようになり,それぞれの形質に対応してそれを規定する仮想的な遺伝因子が設定されてきた。このような遺伝因子はメンデル因子,または単に因子とよばれていたが,W.L.ヨハンセン(1909)の提案した遺伝子という語がしだいにこれにとって代わるようになった。
サットンW.S.Sutton(1902)らはいち早く成熟分裂における染色体の行動がメンデル因子の行動と一致することを明らかにしたが,さらに,1910年に始まるT.H.モーガンらのキイロショウジョウバエの研究により,遺伝子は染色体に線状に配列して連鎖群を形成しており,一つの遺伝子は特定の染色体の特定の部位を占めていることがわかった。このような研究により,それまで仮想的な存在であった遺伝子が物質的基礎をもつことになり,その構造と機能を物質的に研究する道が開けた。
英語の〈gene〉に対し,日本で〈遺伝子〉という語をあてるようになったのは1936-37年ごろである。1936年に編まれた《遺伝学用語》第1輯ではgeneに対し〈遺伝因子〉が,genotypeやgeneanalysisに対しては〈遺伝子型〉や〈遺伝子分析〉があてられている。そして,37年の田中義麿著の《遺伝学》再版で初めてgeneに対し〈遺伝因子〉〈ゲン〉と並んで〈遺伝子〉が登場し,同年の井上頼数の論文に〈易変遺伝子〉の語が現れる。この時期より古い著書・論文では〈遺伝単位〉〈遺伝因子〉〈因子〉〈ゲン〉〈ジーン〉が用いられており,〈遺伝子〉は見あたらない。
1940年代に入り,ビードルG.W.BeadleとテータムE.L.Tatum(1941)らはアカパンカビの栄養要求性の遺伝を研究し,生体内における物質の生合成経路の各段階がそれぞれ固有の遺伝子に支配されていることを知った。すでに,生化学的研究により生合成経路の個々の過程が酵素に触媒されていることがわかっていたので,ビードルはこれらの知見を総合して一遺伝子一酵素説を立てた。こうして,遺伝子が直接支配するのは特定酵素の生産であり,この酵素を媒介にして遺伝子が生合成産物を支配するものと考えられるようになった。その後,酵素はもちろんのこと,酵素以外のタンパク質の生産も遺伝子の直接的支配下にあること,およびタンパク質の多くは複数の同種あるいは異種ポリペプチドからなることがわかってきたため,一遺伝子一酵素説は一遺伝子一ポリペプチド説に修正・拡張されることになった(ハルトマンP.E.Hartman,1965)。
一方,ベリングJ.Belling(1928)らにより太糸期染色体が染色小粒とそれをつなぐ糸状部分からなる数珠状構造を示すこと,およびC.B.ブリッジズ(1935)らにより唾腺(だせん)染色体が染色性の高い横縞部分と染色性の低い介在部分からなることが明らかにされ,染色小粒や横縞が遺伝子に対応するという考えが生まれた。この考えは偽対立遺伝子の組換えや遺伝子の突然変異機構の研究成果をふまえ,遺伝子は機能(すなわち形質の支配)・組換え・突然変異という三つの現象に共通な基本単位であるとするグリーンM.M.Green(1955)らの《遺伝子の統一概念》へと発展した。
しかし,バクテリアやウイルスが研究材料に登場するに及び,これまで検出不能であったごく低頻度の組換え体の検出が可能となった。そして,ベンザーS.Benzer(1957)らにより,交叉(こうさ)部位や突然変異部位は一つの遺伝子が占める染色体上の領域内に多数存在することが明らかにされ,究極的には単一のヌクレオチド対が交叉や突然変異の単位であることが推定されるに至った。同じことは高等動植物の若干の遺伝子についても証明されてきた。一方,遺伝子の本体がDNAであることが確実になり,バクテリアやウイルスでは一つのゲノムの全遺伝子が単一のDNA分子に組み込まれていること,また,高等動植物においても電子顕微鏡で確認できるかぎりでは,1本の染色体に含まれるDNAは連続した1本の糸であることもわかり,遺伝子の統一概念は自然に消滅した。
1950年代以降の分子遺伝学の発達により,まず,遺伝子の本体とポリペプチドの生産を支配する機構の概要が明らかとなった。ほとんどの生物では二重鎖のDNA(一部のウイルスでは1本鎖のDNAや二重鎖あるいは1本鎖のRNA)が遺伝子の本体をなしており,個々の遺伝子の特異性はそれを構成する4種類のヌクレオチド対の数と配列順序によって決められる。二重鎖DNAの転写によりRNAがつくられるが,この際,2本のDNA鎖のうち1本の鎖における4種類のヌクレオチド,すなわち,デオキシアデニル酸(Aで表す),デオキシグアニル酸(G),デオキシチミジル酸(T),デオキシシチジル酸(C)の配列順序に従ってRNAにおける4種類のヌクレオチド,すなわち,アデニル酸(A),グアニル酸(G),ウリジル酸(U),シチジル酸(C)の配列順序が決まる。転写されるDNA鎖のA,G,T,Cに対応してRNA鎖にU,C,A,Gが配位される。
RNAは機能的に3種に分かれる。タンパク質とともにリボソームを構成するリボソームRNA(rRNAで表す),リボソームへアミノ酸を運ぶ転移RNA(tRNA),およびポリペプチドの一次構造,すなわちアミノ酸配列を規定するメッセンジャーRNA(mRNA)である。mRNAに転写された遺伝情報,すなわちそのヌクレオチド配列がポリペプチドの一次構造に変換される過程を翻訳という。このとき,mRNAを構成するヌクレオチドの三つずつが一組になってポリペプチド鎖の一つ一つの位置に入るアミノ酸の種類を規定してゆく。このヌクレオチドのトリプレットには43,すなわち64の種類がある。個々のトリプレットとポリペプチドに含まれる20種類のアミノ酸との対応関係を遺伝暗号とよぶが,これはニーレンバーグM.W.Nirenberg(1966)らの研究によって完全に解読された。64のトリプレットのうち三つはどのアミノ酸にも対応しない。これらはナンセンスコドンとよばれ,翻訳において終止符の役をする。単一のmRNA分子の翻訳により,単一または複数のポリペプチドがつくられる。生産されるポリペプチドの数はmRNA分子に含まれるナンセンスコドンの数で決まる。mRNAを介して個々のポリペプチドに対応する二重鎖DNAの領域が,一遺伝子一ポリペプチド説にいうところの遺伝子である。このカテゴリーの遺伝子をシストロンまたは構造遺伝子とよぶ。これはポリペプチドの一次構造を決定する遺伝子という意味である。これに対し,転写によって個々のrRNA分子やtRNA分子をつくるDNA領域をrRNA遺伝子やtRNA遺伝子といい,これらがつくるRNAは翻訳されることがない。
遺伝子の本体や作用機構に関する研究と並んで,その作用の調節機構も研究されるようになった。F.ジャコブとJ.モノー(1961)らの研究から構造遺伝子の作用は作働遺伝子や促進遺伝子の働きにより調節されていることがわかってきた。大腸菌のLac遺伝子の場合,その作働遺伝子は他の構造遺伝子が生産するタンパク性抑制物質の結合部位であり,促進遺伝子は転写をつかさどるRNAポリメラーゼの結合部位である。ここに,遺伝子作用の調節に直接関与する種々の物質の結合部位を構成しているDNA領域が新しいカテゴリーの遺伝子として浮かび上がってきた。
その後,転写中のDNA分子や,DNA・DNAあるいはDNA・RNA分子雑種の電子顕微鏡による直接的観察法の確立,およびmRNAの逆転写によるcpDNAの調製,DNAのクローン化,そのヌクレオチド配列の決定などに関する技術的進歩により,真核生物の遺伝子の構造や作用発現について多くの新知見が加わった。遺伝子と遺伝子の間には,スペーサーとかサイレントDNAとよばれる転写されないDNA領域がふつう存在する。その一部はヌクレオチド配列からみて,現在も活動中の遺伝子と起源を同じくする遺伝子の残骸とみなされ,偽遺伝子とよばれる。また,一つの遺伝子の領域内に,転写に先立ってDNA分子内組換えによって除去されてしまうDNA部分や,翻訳に先立ってRNA分子から除去されるRNA部分に対応するDNA部分が存在することもわかった。構造遺伝子の領域内にあって翻訳に関与しないこのようなDNA部分をイントロンintronまたは介在配列とよぶ。これに対し,翻訳にあずかるDNA部分をエクソンexonという。真核生物の構造遺伝子はふつう複数のエクソンとイントロンがモザイク状に配列したものであり,イントロンを包含しない原核生物の遺伝子とは内部構造が異なっている。
このように多様な構造と機能を考慮するとき,遺伝子の包括的定義としては〈DNA(RNAウイルスにあってはRNA)分子中の,遺伝的になんらか固有の意味をもつ,決まった長さのヌクレオチド配列〉とするのが適当と考えられる。ここにいう“遺伝的な意味”は主として機能的なものであり,RNAやポリペプチドの構造を規定したり,遺伝情報の発現・調節に関与する種々の分子の結合や離脱部位を構成したりすることであるが,偽遺伝子の場合のように,その起源を示唆する歴史的な意味も包含する。
遺伝子はその細胞内の所在する場所により,核内遺伝子,染色体遺伝子,染色体外遺伝子,細胞質遺伝子,葉緑体遺伝子,ミトコンドリア遺伝子などに分類される。核内遺伝子と染色体遺伝子,染色体外遺伝子と細胞質遺伝子は同義である。葉緑体遺伝子とミトコンドリア遺伝子はともに細胞質遺伝子に含まれる。これらの遺伝子の間にはその構造や機能に関して本質的な差がない。しかし,染色体と細胞小器官(オルガネラ)DNAの間には次代への伝達様式に差があり,核内遺伝子はメンデルの遺伝法則にのっとって遺伝するが,細胞質遺伝子の伝達はこれに従わない(細胞質遺伝)。遺伝子はまた,その効果の表れ方により主働遺伝子,微働遺伝子,致死遺伝子,ポリジーンなどに分類される。
執筆者:常脇 恒一郎
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
gene
生物の親から子へ遺伝情報を伝え,親子が似た性質を示す遺伝現象を担保する物質。本体はDNA(一部のウイルスではRNA)である。DNAが通常示す二重らせん構造が相補的構造をもち自己複製ができるため,細胞分裂の際に同じ情報を継承できる。核酸分子のうち,タンパク質のアミノ酸配列に翻訳される領域(翻訳領域)と,翻訳領域の発現を調節する情報をもつ領域(転写調節領域)とを合わせて遺伝子と呼ぶことが多い。転移RNA(tRNA),リボソームRNA(rRNA)等の転写産物そのものが機能をもつ非翻訳RNAをコードする領域も遺伝子に含めることがある。細胞の生存や増殖のために必要な酵素の遺伝子のように,どの細胞でも発現するものをハウスキーピング遺伝子という。一方,ヒトのような多細胞生物では,多種多様な分化した細胞があり,それぞれで特有の遺伝子が働いている。これら多細胞生物における細胞分化や形態形成は,転写因子やシグナル伝達因子等の遺伝子発現の調節や情報伝達を行う遺伝子産物により制御される。もともと遺伝子は,それが決定する生物の特性(表現型)や,それがコードするタンパク質の機能に応じて定義されたが,近年のDNA塩基配列解読の高速化に伴い,対応する表現型やコードするタンパク質の機能が未知の遺伝子が多数同定されている。遺伝子は,生命の起源以来,地球環境の変遷とともに進化してきたものであり,その塩基配列には地球と生命の歴史が刻まれている。
執筆者:遠藤 一佳
出典 平凡社「最新 地学事典」最新 地学事典について 情報
Sponserd by ![]()
一つの形質を決定する遺伝単位で,その本体は,RNAウイルスの場合を除いて,一般にはDNAであって染色体上に存在する.遺伝子にはタンパク質の一次構造を決定し,RNAポリメラーゼⅡによって転写されるものや,リボソームRNAの遺伝子のようにRNAの一次構造のみを決定し,RNAポリメラーゼⅢで転写されるものなどがある.タンパク質の遺伝子はDNAの三つの塩基配列によってアミノ酸を指定し,これはそのタンパク質のアミノ酸配列を決定している(コドン).このとき,1遺伝子-1酵素の対応関係が成立する.遺伝学的にシス-トランス相補性試験によって判定されるシストロンは,1個のタンパク質分子の構造遺伝子に対応するものといえる.
出典 森北出版「化学辞典(第2版)」化学辞典 第2版について 情報
Sponserd by ![]()
Sponserd by ![]()
…一つ一つの性質(形質という)をとっても,子どもが親にそっくり似ることはむしろまれである。しかし,このような場合でも,これから述べるように,子どもは両親からそれぞれの形質を支配する遺伝子をちゃんと受け継いでいる。そこで学問的には,形質と遺伝子の両方について,それらが親(または先祖)から子ども(または子孫)へ伝わることを遺伝といっている。…
…しかし文化の発達とともに環境自体が変化しつつあるから,昔の環境に対する適応が現在の選択に対して必ずしも最も有利であるとはいえない。(2)変異度の増加 二つの人種集団から生じた混血集団における遺伝子度数は,両母集団の中間にあって,母集団の遺伝子度数と混合率によってきまる。また混血集団の変異度は一般に母集団相互間の変異度よりも小さい。…
…人類における遺伝現象を研究する医学,生物学の一分野で,ヒトの生命現象とその変異を遺伝子や染色体との関連のもとに研究する学問である。 ヒトの遺伝子の大部分は細胞の核の染色体に存在し,両親はそれぞれ自己の遺伝子の半分を,精子と卵子(配偶子)の染色体を通して子に伝達する。…
…デオキシリボ核酸deoxyribonucleic acidの略称。核酸のうち糖成分がD‐デオキシリボースであるもので,遺伝子の本体をなす。教育や体験によらずに親から子へと自動的に伝わる性質を遺伝形質という。…
…生物のもつ遺伝物質が量的・質的に変化すること,およびその変化によって生じた状態を指す。細胞核内の遺伝物質の突然変異(核性突然変異)は,変化が起こる遺伝物質の単位に応じて,ゲノム突然変異,染色体突然変異,遺伝子突然変異に分けられる。優性突然変異,劣性突然変異という区別は,突然変異がヘテロ二倍体で表現型の変化をもたらすか否かの違いをいうのであり,また常染色体突然変異,性染色体突然変異という区別は,突然変異が起こった染色体の違いをいうものであるが,いずれも突然変異の分類としては基本的ではない。…
※「遺伝子」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
二十四節気の一つで,二至 (夏至,冬至) ,二分 (春分,秋分) として四季の中央におかれた中気。元来,春分は太陰太陽暦の2月中 (2月後半) のことで,太陽の黄経が0°に達した日 (太陽暦の3月 2...