デジタル大辞泉 「種子」の意味・読み・例文・類語
しゅ‐し【種子】
[類語]種・種物・核・
Sponserd by ![]()
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
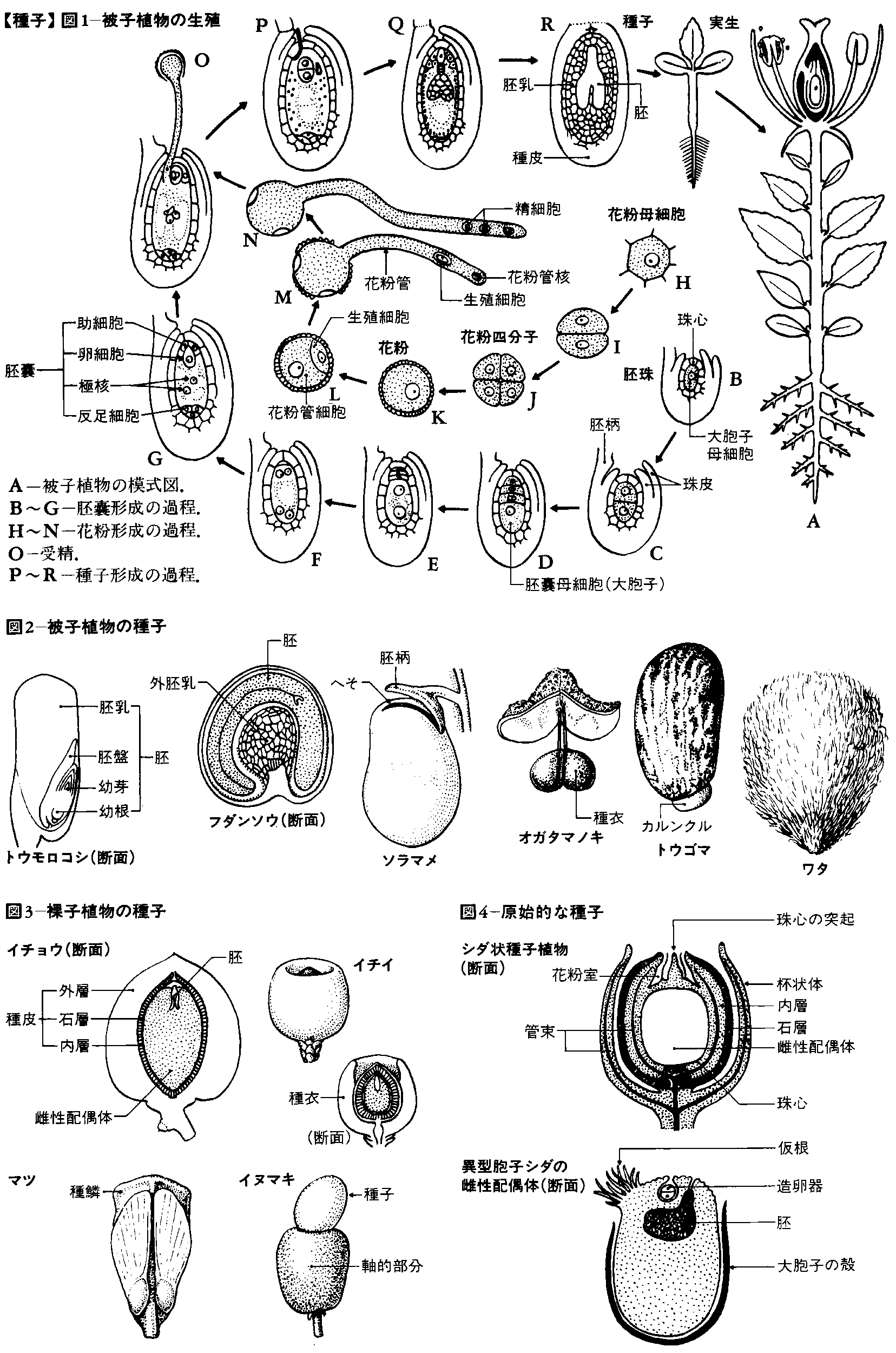
裸子植物および被子植物がもつ生殖器官の一つ。日常的には種(たね)とよぶことが多い。生活環のうえでの種子の機能は、中に包まれている胚(はい)(若い胞子体)を母(ぼ)植物とは別の場所に散布し、そこで胚を発芽させるか、または条件の悪いときには休眠させておくことである。なお、種子と胚珠とは同義に近いが、普通、成熟して休眠状態にあるものを種子、発達中のものを胚珠とよぶ。
種子のおもな構成要素は、種皮、栄養組織、胚の三つである。
[山下貴司]
種皮とは種子の周囲を覆う皮膜をいい、胚珠における珠皮が変化したものである。珠皮は1枚のもの(多くは裸子植物と被子植物合弁花類)と2枚のもの(多くは離弁花類と単子葉類)とがあり、後者では種皮もこれに対応して、2枚からなる。ときには、外種皮と内種皮とに区別する。いずれの場合でも、種皮はさらに何層かの性質の異なる組織に分化しているのが普通である。その分化の仕方は植物の種類によってきわめて多様であるため、すべてを示すことはできないが、代表的な例をいくつかあげることとする。
(1)種皮の組織 アブラナ属の種皮では、外種皮と内種皮が区別できる()。外種皮の最外層はペクチンを含む粘液細胞層である。この層は地中で水分に出会うと膨潤して、発芽の準備をする。その内側には、押しつぶされた外種皮の柔組織がある。これは若いときには数層の柔細胞であったが、完熟するとともに細胞質が抜け、内外からの圧力で押しつぶされたものである。外種皮の最内層は厚壁組織(厚膜組織、機械組織ともいう)で、細胞壁にはリグニン(木材素)が沈着して堅くなり、すきまのない層をつくっている。この層は中にある胚を保護するとともに、容易には水を浸透させず、条件がよくなるまで発芽を抑制する働きをもっており、外側の細胞が十分に吸水して一定の時間がたったあとに、初めて内側に水を浸透させる。厚壁組織の存在する場所は植物の種類によって異なり、アブラナ属よりもさらに外側の場合もあれば、内種皮に存在する場合もある。また、堅さの程度も植物によってさまざまである。なお、地中の微生物によって厚壁組織が徐々に分解され、水が浸透することも多い。アブラナ属の内種皮は、若いときには多層の柔組織からなっているが、完熟すると最内層以外は押しつぶされる。この最内層は黒色の色素を含む細胞からできている(色素細胞層)。この層より内側はもはや種皮ではなく、内胚乳(後述)の最外層であるアリューロン層(糊粉(こふん)層)と、細胞質が抜けてつぶれた内胚乳の組織となっている。
以上がアブラナ属の種皮であるが、他の植物の種皮でよくみられる組織として、結晶を含む組織、繊維状の組織、維管束、多肉質の柔細胞からなる組織(多肉組織)などがある。多肉組織は厚壁組織よりも外側にあるのが普通で(イチョウ、モクレン、ザクロなど)、このような種子を動物が飲み込むと、厚壁組織よりも内側は消化されずに排泄(はいせつ)され、散布に役だつとされる。しかし、なかには有毒な多肉組織も多い。また、種皮の表面にはクチクラ(角皮)、気孔、パピラ(乳頭状突起)、毛などが存在することがある。ワタの繊維は表皮細胞が細長く伸び出したものである。
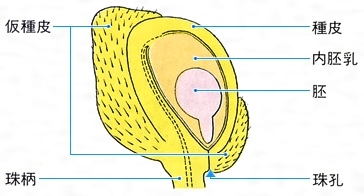
(2)へそ(臍) 種子の表面を観察すると、へそとよばれる特別な部位のあることが多い。これは、胚珠の基部である珠柄が子房内の胎座(裸子植物では種鱗(しゅりん)など)から離れて落ちた痕跡(こんせき)であり、その形と大きさは種類によってさまざまである。倒生胚珠(珠孔が珠柄と同じ側にあるもの)からできた種子では、珠柄の一部が種皮に沿着しており、へその近くに稜(りょう)のような隆起となっていることが多い。こうした種子では、胚珠の発達中に珠柄付近の組織が張り出し、種皮の一部または全部を覆うことがある。このような構造を仮種皮(種衣ともいう)とよぶ。被子植物ではキブシ()のほか、スイレン、ニクズクなどにみられ、仮種皮は本来の種皮よりも柔らかく、不規則な形となる。裸子植物ではイチイ、カヤ、イヌマキなどに多肉質の仮種皮がみられる。仮種皮のなかには、鳥や昆虫による種子散布に役だつものがあると考えられているが、その機能と起源については、まだ不明な点が多い。
(3)珠孔 種子の表面にはもう一つの特別な部位として珠孔がある。その位置は直生胚珠のものではへその反対側、倒生胚珠のものではへそのすぐ近くである。珠孔は胚珠における珠皮の開口部で、受精のときに被子植物では花粉管が、裸子植物では花粉が侵入した場所である。しかし、成熟した種皮では完全に閉鎖され、外見からでは確認するのが困難なことが多い。こうした珠孔の周りに特別な組織が発達することがある。トウゴマなどでは、珠孔を覆ってカルンクルcaruncleとよばれる柔らかい突起ができ、脂質を分泌する()。アリがこの部分を好み、種子を運搬するため、散布に役だつといわれる。また、ツユクサでは胚珠の成長の途中で珠孔の周囲の珠皮だけが早く堅くなる。しかし、他の部分はその後も成長を続けるため、珠孔付近にくびれができ、胚はそのくびれの中に入っている()。発芽のときにはくびれた部分の種皮が蓋(ふた)のように外れ、そこから主根が出てくる。
被子植物には種皮の一部が張り出して翼(よく)となり、風で飛ぶものがある(キリ、ヒメシャラなど)。裸子植物のマツなどの種子にも翼がついているが、これは種鱗の組織の一部がはがれて種子に付着したものである。
[山下貴司]
種子における栄養組織として胚乳の語が使われるが、これには二つの意味がある。広義には種子に含まれる胚以外のすべての栄養組織をさすが、狭義には被子植物の内胚乳をさしている。ここでは狭義としてとらえ、以下、種子における栄養組織に触れる。
(1)裸子植物 裸子植物の成熟した種子の栄養組織は、雌性配偶体(前葉体ともいう)からできている。雌性配偶体の発達は、まず、若い胚珠の珠心(中央を占める柔組織)の中に大胞子母細胞ができ、これが減数分裂して核相n(半数世代)の細胞ができる。この細胞は、初めのうちは核だけが繰り返して分裂し、細胞壁をつくらない、いわゆる自由核分裂を行うが、大きな1個の細胞の中央には液胞があり、周辺部には数千個の核ができる。その後、周辺部から細胞壁の仕切りができ、しだいに中央にまで及んでいく。このころ、珠孔に近い部分に数個の造卵器がつくられ、その中で卵細胞が受精して胚発生が始まるが、雌性配偶体はその後も成長と肥大を続ける()。成熟した種子では、珠心は雌性配偶体によって吸収し尽くされ、死んだ細胞からなる薄膜状の組織として残るだけである。その内側の雌性配偶体が種子の体積の大部分を占め、ここに貯蔵細胞が詰まっている。ただし、中央部は後述する初期胚によって消費され、空洞ができているのが普通である。貯蔵物質はソテツやイチョウではデンプン粒が多く、マツなどの球果類ではデンプンもあるが、脂質とタンパクからなるアリューロン粒が主体である。イチョウの種子(銀杏(ぎんなん))が秋に落下するとき、種皮の厚壁組織を割ると、中から薄膜に包まれた緑色で球形の組織塊が現れる。これが雌性配偶体であり、この部分を食用とする。このなかには初期胚も含まれているが、受精直後であるため、まだきわめて小さい。その後、種子が地中に埋没すると、胚は雌性配偶体を消費して成長を続ける。そして、翌春の発芽の直前には、雌性配偶体はほとんど吸収し尽くされることとなる。
(2)被子植物 被子植物の種子の栄養組織には外胚乳(外乳)と内胚乳(内乳)とがある。外胚乳は、珠心の組織の一部が受精後にもさらに発達して栄養を貯蔵するようになるもので、アカザ科、スイレン科などにみられる()。スイレンでは成熟した種子の体積の大部分を外胚乳が占め、その細胞は肥大して、中にデンプン粒をもっている。珠孔寄りの部分に薄膜状の内胚乳があり、胚はその中に包まれている。
内胚乳は、被子植物のほとんどすべてがもつもっとも主要な栄養組織である。被子植物の雌性配偶体は胚嚢(はいのう)とよばれ、通常は8個の自由核をつくる。そのうちの中央にある2個の核が合体して極核となる。花粉管のもっている2個の精核のうち、1個は珠孔寄りにできた卵細胞と合体して胚をつくり、他の1個はこの極核と合体して内胚乳をつくる。これを重複受精という。したがって内胚乳は核相3n(三倍体)であり、胞子体とも配偶体とも異なる独立した植物体であると考えられている。内胚乳の発達様式には、初め中央に液胞ができ、周辺部に多数の自由核をつくる自由核型(アブラナ科、イネ科など)、初めから細胞壁の仕切りができる細胞型(キク科など)、第1回の分裂のときに細胞壁をつくり、そのあとに自由核分裂をする沼生(しょうせい)型(ユリ科など)の三つがある。ラン科などでは、内胚乳は数回の核分裂を行うのみで退化し、栄養をほとんどもたない種子をつくる。自由核型や沼生型の内胚乳であっても、比較的あとまで発達する場合には周辺部から細胞壁をつくり始める。マメ科やオモダカ科の内胚乳は細胞壁をつくる段階にまで発達するが、その後は胚が急速に肥大して内胚乳は吸収し尽くされてしまう。植物の解説などで、「種子に胚乳はない」と書かれているのはこのような状態をさしている。また、ココヤシの成熟した内胚乳では、周辺部に細胞からできた組織ができ、中央部は巨大な液胞で埋まる。これがココナッツミルクとよばれるものである。イネ科の内胚乳では、成熟すると中心部まで細胞が詰まり、その中にデンプン粒を入れている。内胚乳の貯蔵物質としてもっとも一般的なのがデンプンであり、内胚乳の最外層は普通、タンパクを含むアリューロン層となる。しかし、ユリ科やヤシ科の内胚乳にはデンプンがなく、脂質とタンパク質、および肥厚した細胞壁についているヘミセルロース(セルロースとともに細胞膜を構成する炭水化物)となる。
[山下貴司]
受精卵がある程度発達した胞子体を胚とよぶが、裸子植物と被子植物では、その発達に違いがみられる。
(1)裸子植物 裸子植物の受精卵は、初めに自由核分裂を行ったあと、一部に細胞壁をつくり、この部分が細長い細胞列となって雌性配偶体の中に伸び出す。この段階のものを初期胚という。初期胚は配偶体の組織を溶かしながら曲がりくねって伸び、多くの分枝を行う。やがてそれぞれの枝の先端に分裂組織の塊(胚)ができ、これがそれぞれ子葉や主根などを備えた幼植物の形をつくり始める。したがって、1個の胚珠の中に多数の幼植物ができることになるが、通常は、このうちもっとも早く発達したものだけが完熟時まで残り、他はこれによって吸収される。
(2)被子植物 被子植物の受精卵は、まず細胞分裂をして組織の塊を形成し、これからただちに1個の幼植物をつくり始める。しかし、完熟時の種子に入っている胚をみた場合、胚の発達段階は、種類によってさまざまである。ニリンソウなどのキンポウゲ科の場合でみると、種子が落下したときに入っている胚はまだ分裂していない受精卵であるか、または少数の細胞からなる未分化な塊にすぎず、種子が地中に埋没してから胚は内胚乳から栄養をとって発達する。また、ラン科の完熟種子中の胚も同様な状態であるが、この場合は栄養組織がないので自力で発芽することはできず、地中にある共生菌の菌糸から栄養をとって発芽、成長する。しかし、他のほとんどの植物では胚は胚珠中でさらに発達し、子葉・茎頂・胚軸・主根などの幼植物の諸器官をつくったのち、休眠する。マメ科では胚は内胚乳を吸収し尽くすまで発達し、2個の子葉が肥大して種子の体積の大部分を占め、タンパク質、脂質などを貯蔵するようになる。ほとんどの種子で、胚は主根の根端を珠孔に向けて入っている。このため、休眠中の胚軸と主根の細胞は、長軸方向から扁圧(へんあつ)された形となっているが、これが吸水すると長軸方向に膨らむことから胚軸と主根が急激に伸び、珠孔付近の種皮を押し破って外に出る。
なお、種苗店などで「種子」として扱われるもののなかには、学問的にみると「果実」とよぶべきものも多数含まれている。たとえば、キク科などで種子というのは、実際には痩果(そうか)(単子房、単種の果実)であり、外側の堅い殻は子房壁に由来する果皮で、機能的にはこれが種皮のかわりをしている。果皮の中にはただ1個の種子があり、本来の種皮は薄膜状となっている。また、イネ科で種子とよぶのは、実際には穎果(えいか)(痩果の一種)であり、種皮は子房壁の裏側に合着し、別々にはがすことができなくなっている。
[山下貴司]
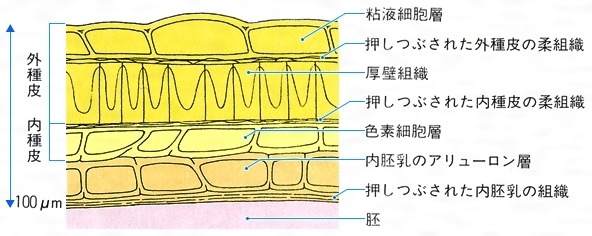
アブラナ属の種皮の断面〔図A〕
キブシの種子の縦断面〔図B〕
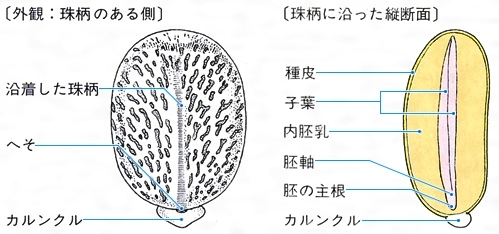
トウゴマの種子と縦断面〔図C〕
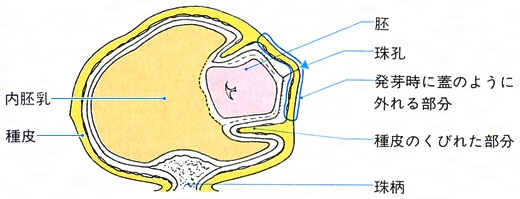
ツユクサの種子の縦断面〔図D〕
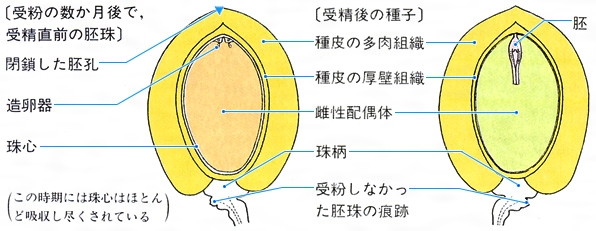
イチョウの胚珠と種子の縦断面〔図E〕
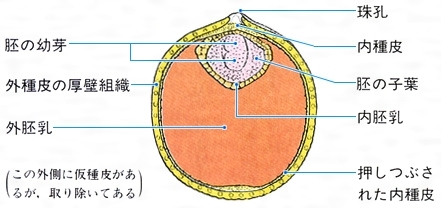
スイレン科の種子の断面〔図F〕

種子の散布方法
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
種(たね)ともいう。人類の主食がコメやムギであることからも明らかなように,植物の種子は古来より動物や人間の生活に欠くことのできないものであった。穀類のほか,クルミやマメ類も食物とされ,ナタネ,ゴマからは油をとり,コーヒー,カカオ,コプラ(ココヤシの胚乳)も種子が原料だし,杏仁(きようにん)(アンズ),蓮子(れんし)(ハス)のように薬としても使われるものもある。ワタの種子の毛からは綿がとられ,また首飾や数珠などの細工物にされる種子もある。
植物学的には,子房中の胚珠が受精後,親植物から養分を吸収しながら生長したもので,幼植物である胚と発芽のさい胚の栄養源となる貯蔵養分と,それらを保護する種皮とからなる。これに対して実は子房が発達したものをいう。種子は,種族を分散させる散布体として,また胚を発芽させ次の世代をになう新しい植物体を生んで,種族を維持する器官として,重要な働きをしている。ここから物事一般の始まりをさす言葉としても用いられることがある。花や球花は種子をつくるための器官である。
種子をつくる植物を種子植物と総称するが,現生のものでは被子植物と裸子植物に大別される。
被子植物の胚珠ovuleは珠心nuccellusとそれを包む珠皮integumentからなり,珠柄funicleにより子房の内壁につく。珠心には卵細胞,助細胞,反足細胞および極核からなる胚囊がある。二つの精核は花粉管とともに胚囊の中へ入る。そのうち一つの精核は卵細胞と接合し,胚embryoとなる。他の精核は二つの極核と受精し,染色体数が3倍(3n)の胚乳(内胚乳)endospermとなり,栄養分をたくわえる。珠皮は,内部の胚と胚乳を保護する種皮seed coatとなる(図1)。胚珠の珠孔は胚が発芽するところで,オニバスのように発芽孔として特別に残っていることもある。種子のへそhilumは珠柄がとれた跡で,水がしみ込みやすい部分と考えられている。
クリ,シソ,ヒマワリ,イネのように種子が子房壁の発達した果皮と一体となっているものは,種子が実に包まれたものであり,広義には実である。クリでは渋皮が,ヒマワリでは果皮の内側の薄皮が真の種皮である。逆に,実のようにみえる種子もある。ルイヨウボタン,ヤブラン,ジャノヒゲでは子房壁が開花後に破れ,胚珠は子房室から露出して種子となり,種皮は多汁質で美しく色づき,果皮状となる。
熟した種子の胚は,普通,子葉および胚軸と幼根からなっているが,イネ科のようにさらに発生が進み,特殊化が起こって複雑となったものもある。ラン科の種子はひじょうに小さく,胚に子葉や幼根の分化はみられない。種まき後ずっと後に胚の発生が起こる。なおラン科の種子は自力で生活できず,地中の菌類から栄養分をもらって発芽する。
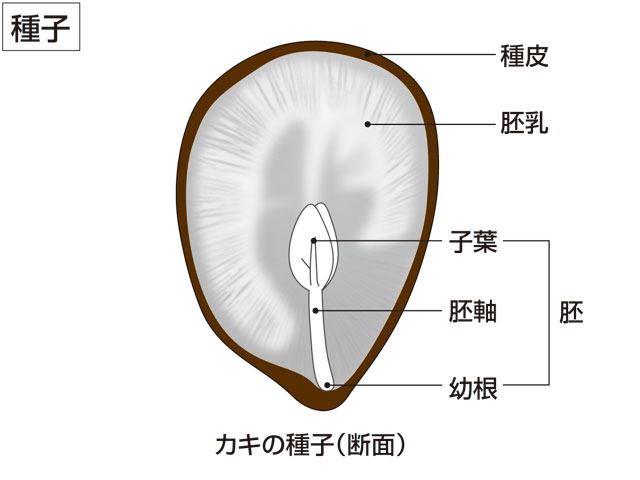
種子の貯蔵養分は胚が発芽し,光合成により自力で生活できるようになるまでの栄養源である。この養分を胚乳にたくわえたものに,セリ,カキ,ナス,イネがある。珠心は普通,消失したり膜状となるが,養分をたくわえて大きくなり,フダンソウのように外胚乳perispermとなることがある。胚乳や外胚乳に栄養分をたくわえたものを胚乳種子albuminous seedという。たとえ胚乳や外胚乳に一時的に栄養分がたくわえられたとしても,種子の成熟の過程で,栄養分がしだいに胚に吸収され,完熟したころには胚が種子の内部をいっぱいにしていることも多い。このような種子を無胚乳種子exalbuminous seedといい,子葉に栄養分を貯蔵している。マメ類,ダイコン,カボチャ,ヒマワリなどがその例であるが,クルミ,ドングリ類,モモなどの子葉は貯蔵器官となって,発芽のさい子葉自身は地上に出ることはない。種子の貯蔵養分としてはデンプン,タンパク質,油脂などであるが,発芽時には糖となって胚の生長に使われる。
胚珠の珠皮は1または2枚であるが,2枚の場合,外側は外種皮に,内側は内種皮になる。カボチャでは外種皮は堅い殻で,内種皮はその内側の薄皮である。一般に,種皮には堅い部分ができて物理的な力に対して強くなり,クチクラ層とともに水や空気を通さなくなる。種皮が水や空気を通さないことは,種子の休眠を可能にしているが,バラ,サンザシのように堅い種皮が胚の生長を制限していることもある。種皮は乾いていて堅いものが多いが,多汁質だったり,肉質の付属体をもつこともある。ザクロの外種皮は表皮組織が長くなり,しかも多汁質で,食べる部分は種皮の一部である。トマトでは柔らかい種皮のまわりに,胎座に由来するぬるぬるした透明な膜がある。しかし,乾いた種子は細胞壁の厚くなった部分だけが残って,毛がはえたようにみえる。多汁質の種皮は,ほかにトケイソウ,パパイア,ジャノヒゲにもみられる。
種皮の表面を包んでいる肉質の部分を広義に種衣arilとよぶ。モクレン,ニシキギ,マユミの種衣は胚柄の一部が発達したものという。ニガウリでは胚珠がついていた胎座に由来している。これらの種衣は普通,あざやかな紅色で,鳥に食べられることにより,種子の散布に役だっている。
トウゴマの種子には胚柄の一部が発達したカルンクルcaruncleとよばれる付属体があり,脂肪を多量に含んでいて脂肪体elaiosomeともいう。この脂肪体はアリが好んで食べ,巣穴へ運ばれるが,種子本体は食害からまぬがれ,種子の散布に役だつ。アリ散布としてはこのほかイカリソウ,クサノオウ,カンアオイなどが知られている。
種皮が粘液質をもっていて,水を含むと表面がねばねばしてくるものに,アマ,カボチャ,オオバコがある。種子の粘着性は,動物に付着して運ばれたり,土の表面に定着したり,水分の吸着,保持に役だつと考えられる。カタバミの種子は,袋状の種皮が反対にそり返ることにより飛散されるが,その表面はねばねばしている。
種子にはカギカズラ,ヤマノイモのように翼があったり,ヤナギ,テイカカズラのように綿毛がはえているものがある。これらも風により種子が散布されることと関係した形態であろう。
高等植物は大地に根をはって生活する動かない生物である。しかし,実や種子の時期には親植物から離れ,いろいろな場所へ分散することができる。上に述べたように,種子が散布のためのさまざまな適応形態をもつことは,種族維持のためである。休眠状態の種子は,低温や乾燥など悪い環境のもとでも耐えられる。移動する時期に,種子が休眠し発芽しないことは,種子の散布定着を安全にしている。マングローブ植物であるヒルギは休眠期がなく,親植物から栄養分を吸収しながら発根し,数cmにもなる。このように樹上で発芽するものを胎生種子という。
種子の寿命は普通1~数年であるが,ヤナギは短命で数日から数十日という。ハスは逆に数百年の長命である。
裸子植物には子房といった構造はなく,胚珠は裸出していて,実はできない。胚は造卵器の中の卵細胞が,精子(ソテツ,イチョウ)または精核と受精してできるが,貯蔵養分は大胞子が発芽してできた雌性配偶体にたくわえられる。ギンナンやマツの実(チョウセンゴヨウ)の食べる部分がこれにあたる。種皮は珠皮の発達したものだが,ギンナンでは外側の臭い肉質部(外層),俗に殻とよばれる堅い部分(石層),さらにその内側の薄皮(内層)の3層がある。マツの種子の翼は,種鱗(種子が付着した部分で,松かさの鱗片)の一部が薄くはがれて種子についたものである。イチイとカヤでは,胚珠の基部の隆起部がふくらんだ種衣があり,イチイではカップ状で赤く熟し,カヤでは樹脂に富んだ繊維質で種子全体を包む。イヌマキの種子は,赤く熟すふくらんだ部分についているが,これは胚珠のつく軸的な部分が肥厚したものという。
種子の起源についてはまだよくわかっていない。最も進歩した現生の異型胞子シダでは,大胞子囊の中で一つの大胞子が発芽し,雌性配偶体となり造卵器をつくるが,大胞子の堅い殻の中におさまっている。栄養的には親植物から独立していて,他のシダ植物と同じ胞子生殖である。デボン紀からジュラ紀のシダ状種子植物(シダ種子類)には種子の化石がみつかっている。大胞子囊に由来する珠心があり,珠皮で包まれ,さらに杯状体によりかこまれている。珠皮と杯状体には管束があり,大胞子の殻も薄くなり,雌性配偶体と胚は親植物から栄養分を吸収しながら生長したと考えられる。また珠皮(種皮)に石層とよばれる堅い部分があり,内部を保護し,休眠にも役だっていたかもしれない。もしそうであれば,シダ状種子植物は,現生の裸子植物のように,種子をつけていたことになる。ソテツなどの珠皮は杯状体と珠皮の合着したものとする見解もある。被子植物の胚珠は子房壁に包まれたため,配偶体が退化して胚囊となり,それを補うかのように,極核が受精し胚乳ができたものである。
執筆者:福岡 誠行
(1)仏教の唯識説の用語 なかでも法相(ほつそう)宗では〈しゅうじ〉,天台宗では〈しゅし〉と読む。認識する対象を生ずる可能性をいう。唯識説では人間の意識の奥にアーラヤ識を想定し,事物の存在はアーラヤ識に内蔵される原因が現れて認識されることによるとし,草木がその種によって異なる花を咲かせることにたとえ,その原因を種子という。(2)密教の用語 種子字ともいい,また種字とも書く。仏・菩薩などをサンスクリット文字(悉曇(しつたん)文字・梵字)の一字で象徴的に標示したものをいう。たとえば,aまたはvaṃが大日如来,hrīḥが阿弥陀如来,saが聖観音,hāṃが不動明王を表す。これらの種子は密教の修法において,本尊となる仏を想起するもととなるので,草木の種にたとえて種子という。お札やお守に種子を書くのは,その仏・菩薩を象徴する意味で,またその姿を書く代りでもある。
執筆者:和多 秀乗
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
seed
胚珠が受精後に発達したもの。次世代植物となる胚と栄養分である胚乳を備え,親個体から離れて発芽でき,休眠することも可能である。化石としてよく産出し,形・大きさ,および種皮の構造・臍点・珠孔などの特徴を用いて同定される。
執筆者:塚腰 実
参照項目:内果皮
出典 平凡社「最新 地学事典」最新 地学事典について 情報
Sponserd by ![]()
唯識説では「しゅうじ」とよむが,密教では種子字(種字)の意味で「しゅじ」とよみ,諸尊を1字で表した梵字をさす。梵字の一つ一つには哲学的意味があるため,それに仏・菩薩を象徴させた。種子を通して諸尊を観じることを種子観,種子を輪形につないで順・逆を観じることを字輪観,種子で描いた曼荼羅(まんだら)を種子曼荼羅という。
出典 山川出版社「山川 日本史小辞典 改訂新版」山川 日本史小辞典 改訂新版について 情報
Sponserd by ![]()
字通「種」の項目を見る。
出典 平凡社「普及版 字通」普及版 字通について 情報
Sponserd by ![]()
…種(たね)ともいう。人類の主食がコメやムギであることからも明らかなように,植物の種子は古来より動物や人間の生活に欠くことのできないものであった。穀類のほか,クルミやマメ類も食物とされ,ナタネ,ゴマからは油をとり,コーヒー,カカオ,コプラ(ココヤシの胚乳)も種子が原料だし,杏仁(きようにん)(アンズ),蓮子(れんし)(ハス)のように薬としても使われるものもある。…
…参加者が総当りするリーグ戦にくらべ試合総数が少なくてすむが,1,2位以外は順位がつかない。強い者同士が早い回で当たらないようにシード法seedを導入することが多い。これは強い者にあらかじめ順番をつけ,意図的に分散して配置する方法である。…
※「種子」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
半夏ともいう。七十二候の一つで,本来は夏至後 10日目から小暑の前日までをいったが,現行暦では太陽の黄経が 100°に達する日 (7月1日か2日) を半夏生とし,雑節の一つとして記載している。この頃半...