デジタル大辞泉 「動物」の意味・読み・例文・類語
どう‐ぶつ【動物】
2 人類以外の動物。特に、哺乳類をいう。獣類。
[類語](1)生き物・
Sponserd by ![]()
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
動物とは,他の生物を食べて独立生活をする生物の総称で,分類学上,植物界に対して動物界Animaliaを構成する。
動物も植物もその体は,水,無機塩,炭水化物,脂肪,タンパク質からなるが,消耗した成分を補い,新しい組織をつくるなど,生活に必要なエネルギーを得るためには栄養分が必要である。緑色植物は栄養分としての炭水化物を,光のエネルギーを用いた炭酸同化(光合成)によって大気中の二酸化炭素と水からつくり出す能力をもっている(独立栄養)。これは葉緑体のなかで行う。ところが動物は,このように無機質を組み合わせて有機質をつくる能力をもたないから,栄養分としての炭水化物やタンパク質は,複雑な有機物,つまり生物自身の生産物という形でなければ取り入れられない(従属栄養)。すなわち動物は,生きるために植物や動物を食べ,それを分解して自己に特有な物質に変えなければならない。
植物は光合成を行うために,葉緑体のある葉や枝を広げ,できるだけ多くの日光を受けとめるように進化した。二酸化炭素,水,無機塩はどこにでもあるから,それらを得るために移動する必要はなく,地中に根をはって静止していればよかった。ところが動物は,食物を探し求めて動き回る必要があった。もちろん動物にも,サンゴ,カイメン,ウミユリのように移動をしないものもあるが,それらは水に浮いて漂うプランクトンを食べているためで,動物としては例外である。
このように動物は運動する必要があるので,体はやわらかく,自由に曲げられなければならない。植物の細胞を包んでいるのがセルロースからなる硬い細胞壁であるのに対して,動物の細胞がやわらかい原形質膜で包まれていることや,体壁と内臓の間などに体腔があるのは,おそらくこのためであろう。体は,移動に便利な球または楕円に近い形となって,運動器官(筋組織,支持組織などで形成されたひれ,脚など)と消化器官(歯,胃,腸など)が発達し,それらの組織に酸素を供給するのに必要な呼吸系(えら,気管,肺など)と循環系が現れ,運搬役の血液などとともに,それらはしだいに効率のよいものに改良されていった。また,代謝の最終産物を体外に排出する腎臓のような排出器官が発達した。一方,食物を探すのに必要な目(眼),触角などの感覚器官と神経系およびその中枢である脳が現れ,脳の発達につれて記憶や学習の能力がしだいに高まり,直面した事態を判断し,推理に基づいて行動するものさえも見られるようになった。
以上のような諸点で,動物は植物と顕著に異なっている。もちろん動物も種々の方法で生殖し,特殊な生殖器官が発達するが,この点は基本的には植物と異ならない。しかしこのような動物と植物の違いは,原生動物,細菌など,単細胞の下等生物では明らかでないことが多い。たとえば,原生動物の鞭毛虫類には,植物のように独立栄養のものと,動物のように従属栄養のものがみられ,動物とも植物ともつかないものが少なくない。
現在地球上に生息(棲息)する動物はきわめて多種多様で,125万以上の種が知られているが,いまだに研究の十分でない分野が少なくないので,将来その総数はおそらく200万種以上に達するであろう。
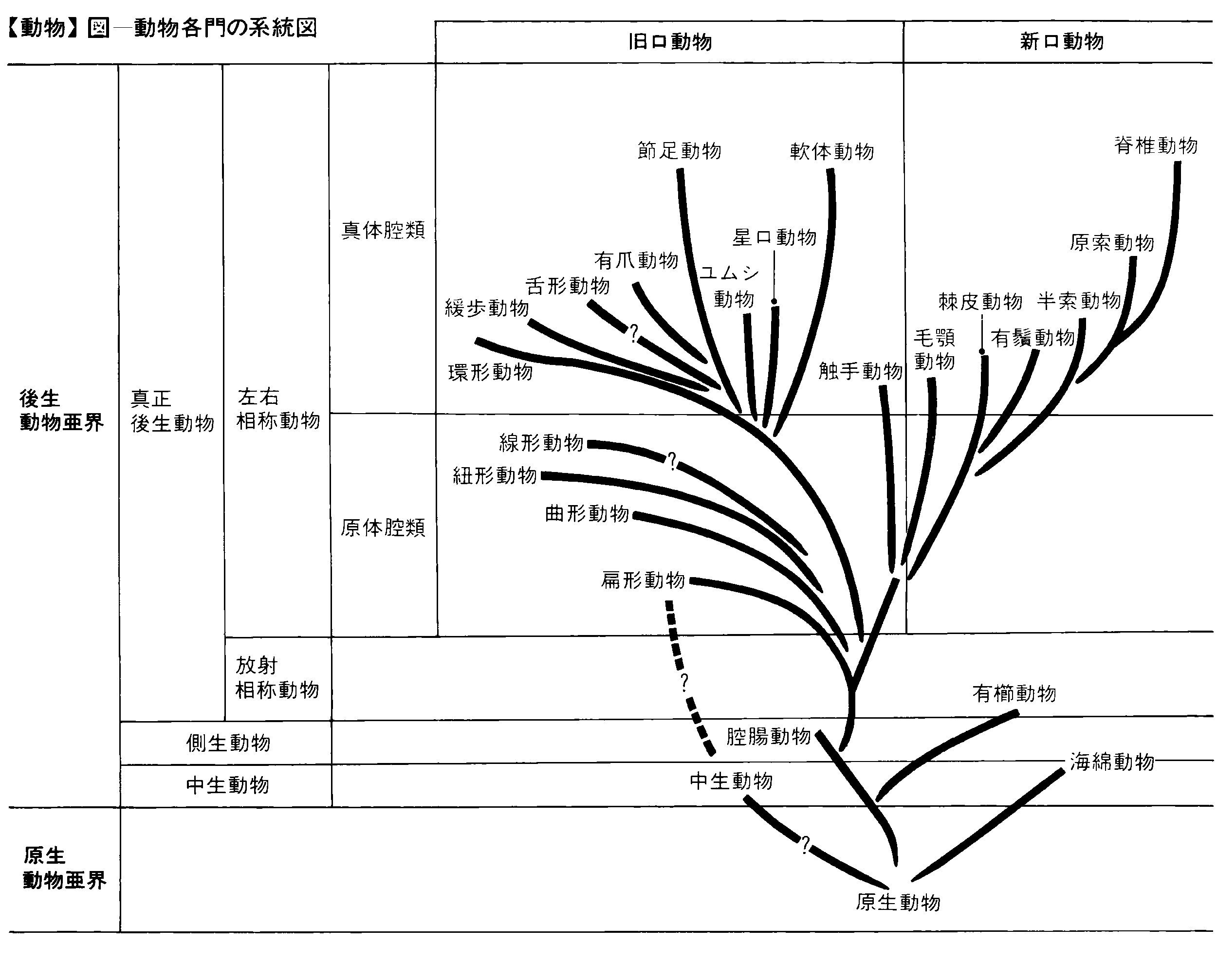
動物の化石は先カンブリア時代末期からも発見されているが,それらはきわめて不完全である。しかしカンブリア紀になると,原生動物の有孔虫や放散虫,海綿動物のフツウカイメン(普通海綿),腔腸動物(刺胞動物)のクラゲ,棘皮(きよくひ)動物のウミユリ,星口(ほしくち)動物,軟体動物,環形動物の多毛類,節足動物の三葉虫,鋏角(きようかく)類および甲殻類など,形態的にはっきり異なった門が突然現れるので,各門の間の系統的な関係を化石をたどって確かめることはほとんど不可能である。したがって門の間の系統関係は,形態や発生から推定するほかない。このため,系統関係にはいくつかの異なった説があり,いまだに安定するに至っていない。以下は,動物の進化経路の概略を示して,複雑きわまりない動物界を単純化して示そうと試みたものである。示した種数は現生種の概数。
動物界には,個体が単一の細胞からなり,偽足(ぎそく)や鞭毛,繊毛などで運動する単細胞動物と,体が複数の細胞からなる多細胞動物がある。前者は原生動物亜界,後者は後生動物亜界に分類される。最近では前者を原生生物界,後者を動物界とすることも多い。
原生動物亜界には,鞭毛虫類(ミドリムシ,ボルボックス),根足虫類(アメーバ,有孔虫),胞子虫類(胞子虫),繊毛虫類(ゾウリムシ,ツリガネムシ)など多数の類がある(3万種以上)。これらはそれぞれ,独立の門とされることもあるが,すべてをまとめて原生動物門とするのがふつうである。現在見られる原生動物は,どれも高度に進化したもので,単細胞とはいえその構造はきわめて複雑で,後生動物の祖先らしい原始的なものは見当たらない。
原生動物以外のすべての動物を一括して後生動物亜界とする。後生動物は,原生動物の原始的なものから分かれ出たに違いないが,どのようなプロセスで多細胞の動物になったのか明らかでない。後生動物亜界には体制の異なった中生動物,側生動物,真正後生動物の3類がある。
この範疇(はんちゆう)には,中生動物と海綿動物が含まれる。
(1)中生動物門 タコなどの腎臓に寄生するニハイチュウなど,約80種からなる〈中生動物〉は,体皮細胞が集まってできた表層が管をつくり,その中に生殖細胞を包んでいる。しかし食物を取り入れる口や胃をもたないだけでなく,細胞が規則正しく並んで組織を構成することもない。原生動物の胞子虫類に近縁で,それと真正後生動物の中間のものではないかともいわれるが,寄生生活のため組織が退化した真正後生動物(扁形動物に近い)とも見られ,真相がよくわかっていない。
(2)側生動物 〈海綿動物門〉(カイメン,約9000種)だけからなり,海または淡水に生息,雌雄異体または同体。体は2種類の細胞(アメーバ状細胞と襟細胞)からなり,ふつう壺状で下端で他物に着生する。上端に1個の大孔,体壁には多数の小孔があり,中央に胃腔がある。体壁は,アメーバ状細胞からなる外側の皮層と,襟細胞からなる内側の胃層で形成され,それら2層の細胞層の間は間充(かんじゆう)ゲルで満たされ,骨片がある。襟細胞は鞭毛をもち,その運動で水は小孔から胃腔に流れこみ,大孔から外へ出る。襟細胞は,水とともに胃腔に入ってきた食物をとらえて細胞内に取り入れ,原生動物のようにその中で消化する。したがって,胃腔は実は胃でなく,大孔も口ではない。皮層と胃層を形成する細胞は互いにゆるく集合しているだけで,真の組織を形成しない。各細胞は刺激感受性をもつが,細胞間の協同作業はまったく行われず,個体としてのまとまりが貧弱で,形や大きさが不安定である。もちろん筋肉,神経,感覚細胞などもない。この類は,原生動物より一段と進化した状態にあることは確かであるが,真正後生動物への進化の一過程を示すものとは考えられない。
このような動物を真正後生動物という。いくつかの組織は集まって,一定の働きをもつ器官を形成し,寄生生活で体制が退化した類を除けば,口と消化管がある。原生動物,中生動物,側生動物以外のすべての動物を含み,体制がきわめて変化に富むが,放射相称動物と左右相称動物に大別できる。
(1)体が放射相称の動物 このような動物を放射相称動物という。腔腸動物門と有櫛動物門からなり,もっとも原始的な真正後生動物で,この類から残りのすべての類が分かれ出たのではないかと考えられている。〈腔腸動物門〉は刺胞動物門ともいい,約9000種よりなる。体が放射相称で,体壁は外胚葉と内胚葉の2層からなり,内胚葉に囲まれた原腸が消化器官の腔腸となり,内胚葉細胞が細胞内消化を行う。腔腸は消化のほか,呼吸や排出の作用もする。原口は成体の口となるが,肛門はなく,中胚葉もない。しかし外胚葉の円柱状の細胞の間には,感覚細胞,神経細胞,刺細胞などがある。海,まれに淡水に生息し雌雄異体。ヒドラ,イソギンチャク,サンゴなどのように海中の岩などに固着しているものと,クラゲのように浮遊性のものがある。クラゲの動きを見れば明らかなように,いくつかの筋肉が統制のとれた運動をし,個体としてのまとまりがある。〈有櫛動物門〉はクシクラゲ類ともいい,約140種よりなる。海生。雌雄同体で刺細胞がなく,体が放射相称でないが,その他の点は腔腸動物によく似ている。
(2)体は左右相称となり,一定の方向に進むのに適する動物 左右相称動物と呼ばれ,前記以外のすべての動物を含み,中胚葉がある。体は相称面が一つしかない左右相称で,前後,背腹の別があり,前端には感覚器官と口が集まっていることが多く(頭化),一定の方向に進むのに適している。ヒトデやウニの成体は放射相称であるが,幼生は左右相称であるから,成体の体型は二次的に変形したものと考えられる。扁形動物を除けばつねに口と肛門があり,消化した食物と未消化のものが消化管のなかで混じる不都合がない。左右相称動物は,発生に際して放射相称動物と同じ状態(囊胚)をかならず経過するので,後者から分かれて進化したものと考えられている。分かれた時代は明らかでないが,およそ10億年ほども昔であったろう。左右相称動物には,胚の原口が成体の口となり,肛門が別にできる旧口動物と,成体の口が原口とは別に口陥から形成され,肛門が原口またはその付近に形成される新口動物がある。これらは,それぞれ別個に腔腸動物から分かれ出て,互いに異なった方向に進化していったものと考えられる。
(1)旧口動物 胚の原口が成体の口になった類で,先口動物ともいう。多くは中枢神経系が体の腹側にあるので,腹神経類ともいう。環形動物,軟体動物,節足動物など多数の門が含まれ,次の2類に大別できる。
 体が前体,中体,後体の3部に分かれないもの 骨格がある場合は,外骨格である。これは細胞が分泌した硬い物質からなり,脊椎動物の骨のような生きた組織ではない。
体が前体,中体,後体の3部に分かれないもの 骨格がある場合は,外骨格である。これは細胞が分泌した硬い物質からなり,脊椎動物の骨のような生きた組織ではない。
(a)二次体腔がないもの 〈扁形動物門〉は寄生生活のフタゴムシ,カンテツ,エキノコックス,ジョウチュウなどのほか自由生活のウズムシなど,約1万7000種がある。体制が簡単で肛門がない。外胚葉と内胚葉の間に中胚葉からできた間充織があり,そこにある隙間に血液やリンパに似た働きをする体液があるが,二次体腔(真体腔)はない。血管系がないかわり,盲囊の腸が体全体に広がっていて栄養を運ぶ。しばしば眼がある。雌雄同体,ときに異体。〈曲形動物門〉は内肛動物ともいい,スズコケムシ約150種よりなる。口と肛門,排出孔,生殖孔を囲む触手冠があり,有柄で多くは群体をつくって海や淡水にすむ。〈紐形動物門〉はヒモムシとも呼ばれ,約1400種よりなる。表皮筋肉層と血管があり,多くは海産(まれに淡水や陸生)で伸縮自在な吻(ふん)を長くのばして獲物を捕食する。この2門は扁形動物に近縁なものと思われる。〈線形動物門または袋形動物門〉には寄生生活のギョウチュウ,カイチュウ,糸状虫(フィラリア),センモウチュウ(旋毛虫),ジンチュウ(腎虫)などのほか,海,淡水,陸で自由生活をするワムシ,イタチムシ,センチュウ(線虫)など約2万4000種。体は長い紐状で,クチクラの外甲がある。腸はまっすぐ体を貫き,肛門が総排出口に終わり,体腔は,扁形動物の間充織に見られる隙間が合一してできた一次体腔で,真の体腔ではない。雌雄異体である。腔腸動物に近い原始的なものとも,また真体腔動物が退化したものともいわれ,系統がはっきりしない。イタチムシを腹毛動物門,ワムシを輪形動物門,ハリガネムシを類線形動物門,トゲカワを動吻動物門などと細分することが多い。
(b)中胚葉で囲まれた二次体腔(真体腔)ができ,体が曲げやすくなっているもの 多くは外套(がいとう)膜からの分泌物で形成された殻があり,軟らかい体を保護する。〈軟体動物門〉は多板類(ヒザラガイ),腹足類(巻貝),掘足類(ツノガイ),二枚貝類(斧足類),頭足類(イカ,タコ)など,海,淡水,陸に生息する約7万種が含まれる。二枚貝類では頭部が退化しているが,他のものには頭部があり,そこに口と重要な感覚器官がある。開放血管系があり,心臓は二次体腔(囲心腔)に囲まれる。水生のものはえらで呼吸するが,陸生のものは外套膜の一部が変化した肺で呼吸する。雌雄異体,または同体。多くのものでは性細胞が囲心腔の中にあり,繊毛ろうと(漏斗)を通って外に出るが,この点は環形動物に等しい。腎臓が腎管であること,トロコフォラに似た幼生とらせん卵割が見られることも環形動物によく似ているので,これらは近縁の関係にあると考えられる。頭足類は,無脊椎動物としての進化のほとんど頂点に達している。すなわち,循環系は大部分が閉鎖的で,食道を取り囲む大きな脳があり,眼はレンズ眼で,構造や性能が脊椎動物の眼によく似ている。血管系のない〈星口動物門〉(ホシムシ類約140種,海生)と,閉鎖血管系をもつ〈ユムシ動物門〉(ユムシ類約145種,海生)は,性細胞を運ぶ腎管や幼生の形態から見て,軟体動物と環形動物の中間に位置するものと考えられる。
(c)二次体腔がある点は軟体動物に等しいが,体が体節に分かれ,表面が硬い外骨格でまもられていて,足はあってもごく短いもの 〈環形動物門〉はゴカイ,ミミズ,ヒルなど,約1万4000種以上よりなる。海,淡水,陸で自由生活をするものが多いが,寄生性のものもある。体は紐状に長く,体節に分かれている。頭部以外の体節は,互いによく似ていて,それぞれに1対の二次体腔と神経節および腎管がある。腸はまっすぐな管で,口は頭部の腹側に,肛門は体の後端部にある。血管系は完全に閉鎖され,血液細胞には呼吸物質(クロロクルオリンまたはヘモグロビン)がある。前進は,表皮にある剛毛と,体側部にあるいぼ状の足(いぼ足)で行う。雌雄異体または同体。ミミズは毛細管で,ヒルは皮膚で呼吸する。ゴカイにはレンズ眼をもったものがある。この類は,外形は顕著に異なるが,軟体動物に近縁と考えられる。水辺のコケの中などにすむ微小なクマムシ類〈緩歩動物門〉(約110種)は,つめを備えた4対のごく短い足で歩く。呼吸器官や循環器官はない。陸生の肉食脊椎動物に寄生する〈舌形動物門〉(シタムシ類,約60種)も4対の足をもつが,そのうちの2対はふつう退化する。これらは環形動物と節足動物の中間のものであろう。熱帯の湿地に生息する〈有爪(ゆうそう)動物門〉(カギムシ類,120種)は,目だった頭節がなく,体の分節が同質的で,いぼ足に節がないなどの点が環形動物に等しいが,いぼ足の先端にはつめがあり,体腔が一次と二次がいっしょになった混成体腔で,キチン性のクチクラ(外骨格)をもち,原始的な気管があるなどの点は節足動物に等しく,後者の祖先に近いものと考えられる。
(d)節のある長い足ができて歩行が自由になり,感覚器官が発達するもの 〈節足動物門〉は体は体節の一部が合一して,頭部,胸部,腹部などとなり,足(脚)は長くのびて多くの可動性の節に分かれ,先端にはつめがあって歩くのに適する。体の表面はキチン性のクチクラでおおわれる(キチン骨格)。心臓は長い管状で,感覚器官としては,この類に固有の1対の複眼が発達するほか,たいてい単眼や触角などがある。脳は前大脳,中大脳,後大脳に分かれる。触角とあごがない節口(せつこう)綱(カブトガニ,約5種,海生),蛛形(ちゆけい)綱(サソリ,クモ,ダニなど,約8万5000種,陸生)および皆脚(かいきやく)綱(ウミグモ,約1200種,海生),対をなしたはさみ状の上あご(大あご)をもつ甲殻綱(ミジンコ,フジツボ,エビ,カニなど,約4万8000種,海,淡水,水辺),気管で呼吸し多数の脚がある多足上綱(ムカデ,ヤスデなど,1万3500種,陸生),同じく気管で呼吸するが,脚が3対(6本)しかない昆虫綱(約95万種以上100万種を超えるともいわれる)がある。昆虫類は地上のあらゆる環境に進出し,その多くが2対の翅を獲得して空中にまで生活圏を広げた。またシロアリ,アリ,ハチなどのように,社会生活をするものも見られる。気管系は翅の中にまで広がり,全身にくまなく酸素を送るが,そのかわり血管系は退化している。節足動物は,種類数において全動物の半分,あるいはそれ以上を占め,海,淡水,陸のあらゆる環境に栄えている。
 体が前体,中体,後体の3部に分かれるもの 細胞内にできた内骨格が見られ,この点は原始的な新口動物に似ている。と同じく旧口動物である。〈触手動物門〉がこれに属し,海生のホウキムシ類(ホウキムシ綱,約10種),コケムシ類(外肛綱,約5000種)およびシャミセンガイ類(腕足綱,約380種)を含む。これらはそれぞれ,別の門とされることもあるが,どれも固着性の原始的な旧口動物で,口を取り巻く触手の集り(触手冠)の外側に肛門がある。触手冠は,中体の上皮細胞中にできる軟骨状の内骨格で支えられる。後体は袋状に管または殻のなかにぶら下がっている。繊毛をもった触手は,食物のプランクトンだけでなく,酸素を含んだ水をも口に送る。体が3部に分かれること,内骨格ができる傾向が見られることなどは原始的な新口動物に等しいから,この類は,旧口動物と新口動物の分岐点近くに位置するものと考えられる。最古の化石は約6億年前のカンブリア紀下部から知られるが,現在と形態がほとんど違っていない。
体が前体,中体,後体の3部に分かれるもの 細胞内にできた内骨格が見られ,この点は原始的な新口動物に似ている。と同じく旧口動物である。〈触手動物門〉がこれに属し,海生のホウキムシ類(ホウキムシ綱,約10種),コケムシ類(外肛綱,約5000種)およびシャミセンガイ類(腕足綱,約380種)を含む。これらはそれぞれ,別の門とされることもあるが,どれも固着性の原始的な旧口動物で,口を取り巻く触手の集り(触手冠)の外側に肛門がある。触手冠は,中体の上皮細胞中にできる軟骨状の内骨格で支えられる。後体は袋状に管または殻のなかにぶら下がっている。繊毛をもった触手は,食物のプランクトンだけでなく,酸素を含んだ水をも口に送る。体が3部に分かれること,内骨格ができる傾向が見られることなどは原始的な新口動物に等しいから,この類は,旧口動物と新口動物の分岐点近くに位置するものと考えられる。最古の化石は約6億年前のカンブリア紀下部から知られるが,現在と形態がほとんど違っていない。
(2)新口動物 胚の原口が成体の肛門になり,口が二次的にできる類で,体は3部に分かれ,真の体腔(二次体腔)がある。骨格ができる場合は,生きている硬い組織で形成された内骨格で,外骨格はない。
血管系がないもの ヤムシ類約110種よりなる〈毛顎動物門〉は,体が左右相称で頭部,胴部,尾部の3部からなり,神経系は腹側にあり,呼吸器,排出器がない。胚の原口の位置に成体の肛門があるが,発生は触手動物に似ている。ヒトデ,ウニ,ウミユリ,ナマコなど,約7400種からなる〈棘皮動物門〉は,体が放射相称であるが,腔腸動物の放射相称とは違って二次的に変わったもので,幼生には左右相称の時期がある。外胚葉の下にある結合組織中に内骨格がある。血管系はなく,そのかわり呼吸,循環,運動に関係した特有の水管系がある。神経系には中枢(脳)がない。最古の化石はカンブリア紀から知られる。
血管系ができ,神経系は背側にあるもの 〈有鬚(ゆうしゆ)動物門〉はヒゲムシの類,約160種。体は3部に分かれ,前体に多数の触手があり,呼吸と食物の摂取をする。口や腸は発生初期にはあるが,後になくなる。血管系と心臓があり,神経系は背側にある。体腔は半索動物と同じく,各体部に分かれている。ギボシムシなど約95種からなる〈半索動物門〉は,体が前体(吻),中体(襟),後体(腹または尾)の3部に分かれ,前体には無対の,中体と後体にはそれぞれ1対の体腔がある。口と腸があり,呼吸は脊索動物と同様,前腸にできた鰓裂(さいれつ)で行う。血管系と心臓があり,主部は腹側に位置し,神経系は腹側にもあるが,主部は背側のものである。
 体の背側に体を左右にくねらせて運動するのに適した脊索ができるもの 〈脊索動物〉がこれに属する。残りの動物全部を含む類で,ホヤ,ナメクジウオからヒトまでが入り,少なくとも発生の途中では次のような特徴を示す。細胞でできた弾力性のある棒状の脊索があって,中軸骨格の役をする。脊索の両側には多数の筋節があり,これらを収縮させ,体を左右にくねらせて前進する。しかし脊索は,多くの高等な類では,発生の途中で脊椎に置き換えられる。中枢神経は脊索の背側にあり,中空,管状で曲げやすく変わっている。前腸に鰓裂,中胚葉で包まれた体腔,ほんとうの尾(肛門より後ろの部分)がある。この類は,このように多数の重要な特徴を共有するため,単一の門〈脊索動物門〉とされることがある。しかし,これらを二つの門に分ける見解もあり,ここでは後者に従っておく。
体の背側に体を左右にくねらせて運動するのに適した脊索ができるもの 〈脊索動物〉がこれに属する。残りの動物全部を含む類で,ホヤ,ナメクジウオからヒトまでが入り,少なくとも発生の途中では次のような特徴を示す。細胞でできた弾力性のある棒状の脊索があって,中軸骨格の役をする。脊索の両側には多数の筋節があり,これらを収縮させ,体を左右にくねらせて前進する。しかし脊索は,多くの高等な類では,発生の途中で脊椎に置き換えられる。中枢神経は脊索の背側にあり,中空,管状で曲げやすく変わっている。前腸に鰓裂,中胚葉で包まれた体腔,ほんとうの尾(肛門より後ろの部分)がある。この類は,このように多数の重要な特徴を共有するため,単一の門〈脊索動物門〉とされることがある。しかし,これらを二つの門に分ける見解もあり,ここでは後者に従っておく。
(a)はっきりした頭がなく,体は胴と舵状の尾からなり,軟骨または硬骨の脊柱や頭骨はないもの 〈原索動物門〉が含まれ,これに脊索が幼生の尾部にしかない被囊(ひのう)亜門と,脊索が成体にもあり,体の全長にわたっている頭索(とうさく)亜門がある。被囊亜門はオタマボヤ,ホヤ,サルパの類で,約2500種があり,海生。血管は開放性,口から入った水を鰓裂から体外に出し,呼吸をすると同時にプランクトンをとらえて食べるろ過食者である。雌雄同体。頭索亜門は無頭類ともいい,ナメクジウオ約35種がある。海生。神経管の前端が広がって脳胞となるが,これはほんとうの脳ではない。赤血球がない。前者と同じくろ過食者であるが,雌雄異体で,眼点がある。
(b)軟骨または硬骨の脊柱と頭骨ができて脳を包み眼はレンズ眼で,ヘモグロビンを含んだ赤血球ができて酸素を運ぶ能率が高まるもの 〈脊椎動物門〉がこれに属する。これには次のような進化の段階を異にする現生の綱があり,これらは魚類と四足動物の2上綱に整理される。なお魚類と両生類は,胚に羊膜ができないので一括して無羊膜類と呼ばれることがあり,ふつう体外受精である。爬虫類,鳥類,哺乳類の3綱は,胚が羊膜で保護されるので有羊膜類と呼ばれ,終生空気呼吸で,体内受精である。鳥類,哺乳類は恒温性で,心臓は2心房2心室よりなる。
(イ)魚類 水生でひれとえらがあり,脚がない。そのうち無顎(むがく)綱はヤツメウナギなど,約45種からなる。海,淡水生であごがなく,鼻孔は1個,対鰭(ついき)はない。原則としてろ過食性で,歯は表皮性で角質。以下のものには可動性の上あごと下あごがあり,真皮性の歯をそなえ,鼻孔が2個,対鰭が2対ある。軟骨魚綱はサメ,エイなど,約900種からなる。海生で骨格は軟骨性,鰓蓋(さいがい)がなく鰓裂が露出する。ろ過食性でなく,おもに遊泳動物を捕食する。硬骨魚綱は,チョウザメ,ウナギ,アユ,シーラカンス,肺魚など,約2万4000種が含まれる。前者に似るが骨格が硬骨に変わり,鰓蓋がある。海から淡水まで生息圏を広げている。
(ロ)四足動物 成体は空気呼吸で,四肢をもち,あごがある。骨格は硬骨性,多くは鼓膜がある。両生(棲)綱はイモリ,カエル,アシナシイモリなど,約4500種を含む。幼生は淡水生でえらで呼吸するが,成体は多くは陸生で肺で空気を呼吸し,対鰭が四肢に変わって陸上の歩行に適する。爬虫綱はカメ,トカゲ,ヘビ,ワニなど,約6000種からなる。多くは陸生だが,淡水,海にも生息する。殻のある卵を陸上に産み,変温性。中耳小骨は1個,体は表皮性のうろこでおおわれる。鳥綱は,ダチョウ,ペンギン,カモメ,キジ,スズメなど,約8600種を含む。中耳小骨は1個,上あごと下あごが方骨(ほうこつ)と関節骨で関節し,下あごが数個の骨でできているなど,爬虫類に似るが,前肢が翼になり,皮膚がうろこの変化してできた羽でおおわれる。現生のものには歯がない。卵生で,巣をつくり,雛を育てる。哺乳綱は,カモノハシ,カンガルー,サル,ウシなど,約4500種よりなる。下あごは歯骨だけでできていて,上あごの鱗骨(りんこつ)と関節する。方骨と関節骨が加わって中耳小骨が3個になり,内耳の骨性迷路が蝸牛殻(かぎゆうかく)となり,コルチ器が発達して,聴覚が鋭くなった。コウモリ,トガリネズミ,イルカなどでは超音波を声帯から発して方向探知を行う。また,鼻腔(びこう)ができ,多くのものでは嗅覚(きゆうかく)が鋭い。ほとんどのものが胎生で,子を乳腺から分泌する乳汁で育てる。皮膚には毛,汗腺,臭腺ができる。大脳は体の大きさに比べきわめて大きいだけでなく,記憶,思考などの精神作用をつかさどる新皮質がその表面をほとんどおおい,いわゆる知能が高い。歯が分化するなど,器官系が極度に発達し,体制が動物中もっとも複雑である。
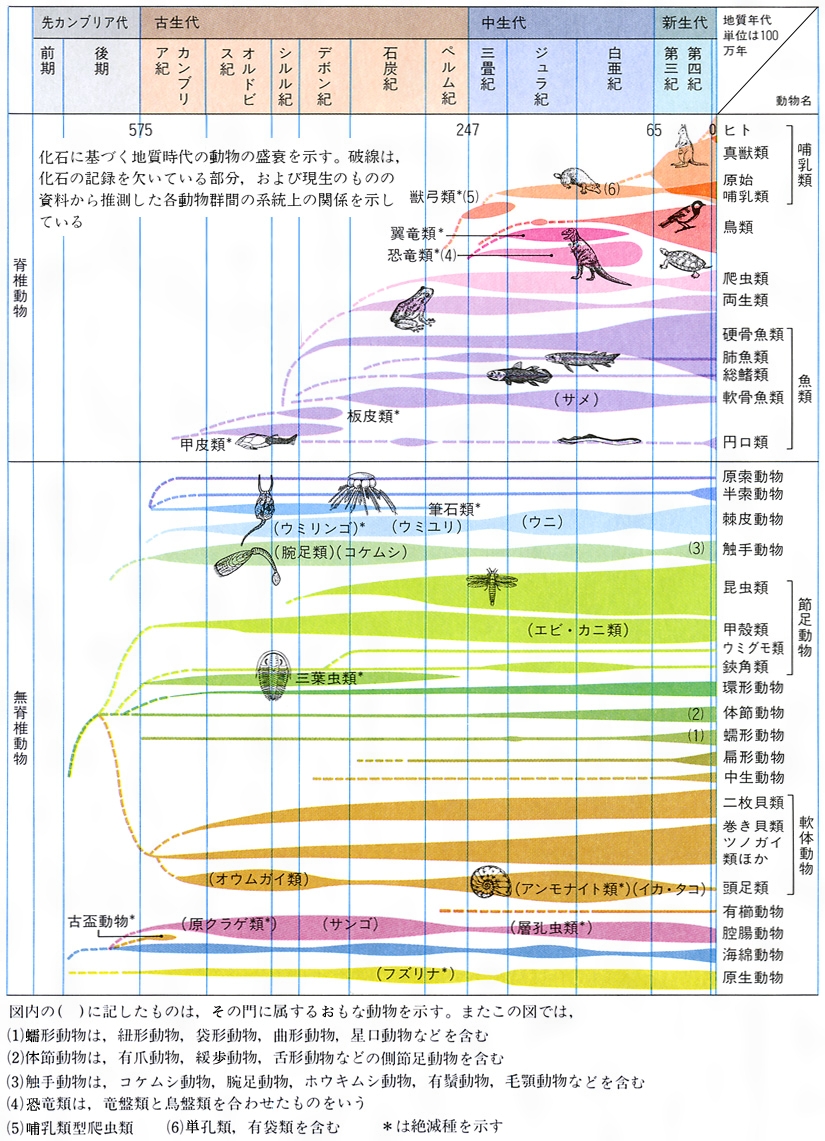
脊椎動物は化石が多く,系統発生がほぼ完全に近いまで解明されている。最古の無顎類はオルドビス紀前期に現れ,これから絶滅した板皮(ばんぴ)綱(あごがあり,体の前半は骨質の甲でおおわれる。骨格の一部は骨質)がシルル紀に分かれ,デボン紀に栄えた。軟骨魚類と硬骨魚類は,生まれたばかりの板皮類からシルル紀にほぼ同時に分かれたと推定されるが,最古の化石は硬骨魚類がシルル紀後期,軟骨魚類はデボン紀中期にならないと見られない。両生類はデボン紀後期に,原始的な硬骨魚類(肺魚類)から分かれ,爬虫類は石炭紀中期に,原始的な両生類のセイモウリアから,鳥類は三畳紀後期に,爬虫類の中の恐竜とワニの共通の先祖から分かれたと推定されるが,最古の化石はジュラ紀後期に始祖鳥が見られるにすぎない。哺乳類も三畳紀後期に爬虫類の中の単弓類から分かれ出たらしく,三畳紀末に最古の哺乳類エオゾストロドンが現れている。しかし,単弓類は石炭紀後期に早くも他の爬虫類と分かれて別個の進化経路を取っているから,単弓類は原始的な哺乳類だと考えることもできる。
執筆者:今泉 吉典
いかなる民族も,さまざまの動物を比喩に使ってことわざや寓話をつくりあげている。何種類もの動物が登場する《イソップ物語》がそのもっとも代表的な例である。しかしそうした雑多な動物イメージの使用になんらかの法則性はないものだろうか。ヨーロッパに関する限り一応次のような傾向性に基づいて,動物イメージが変化していったと考えることができる。すなわちまず鹿のシンボルであり,次いで羊のシンボル,それから牛馬のシンボルといった順序での変化である。そしてそれらのシンボルの移行は,ヨーロッパ人が狩猟段階から牧畜段階,次いで農業段階に移行したことに対応する。つまり狩猟段階には鹿のイメージが,牧畜段階には羊のイメージが,農業段階には牛馬のイメージが対応するわけである。
実際,デンマーク,ゴネスドロブの泥炭の沼から出土した銀めっきの銅の大鉢にはいろいろの森の動物に囲まれて鎮座する〈角のある神〉がレリーフで描かれているが,これは鹿の角をもつ神像であり,ケルト人が崇拝していたものである。またパリのノートル・ダム大聖堂の真下から発掘された祭壇の石にも,Cernunnos(ラテン語で〈角あるもの〉の意)の銘とともに,やはり2本の鹿の角を生やした神像が刻まれている。そしてこれもキリスト教布教以前にケルト人の間で崇拝されていた神の姿である。そしてこれらの有角神は,ギリシア神話の半獣神パンなどのイメージと複合しながら,やがてキリスト教文化の下で悪魔として零落していくことになる。その場合は鹿とともに山羊のイメージも付与された。
次にキリスト教では好んで羊のシンボルが使われる。羊は主人に従順な動物として,神およびキリストに信仰をささげるキリスト教徒のシンボルとされる。しかしキリスト教文化圏では単に羊を信者のシンボルに使うだけでなく,羊をめぐるさまざまのイメージ群を総動員して,キリスト教的世界の構造の総体を表現しようとする試みがなされた。すなわち,羊を信者のシンボルとすれば,羊飼いは信者の保護をつとめとする聖職者,そしてその聖職者に司牧の任を授けるキリストのシンボルとなる(善き羊飼い)。キリスト自身が小羊にたとえられることもあり,〈神の小羊Lamb of God〉はキリストの称号の一つである。また羊を守る囲いは教会のシンボルとなった。さらにそうした羊に襲いかかって連れ去るオオカミは,キリスト教徒を教会から離脱させ,異なった教団へと誘惑する異端のシンボルとなり,そうしたオオカミと身体を張って戦い,羊飼いおよび羊たちのために全力を尽くす牧羊犬は,異端撲滅のための説教修道士や十字軍戦士のシンボルとなった。そのもっとも典型的な図像表現がフィレンツェ,サンタ・マリア・ノベラのフレスコ画であろう。そこにはカタリ派に説教するドミニクスとともに,羊,オオカミ,牧羊犬が描かれている。羊がキリスト教徒の,オオカミがカタリ派の,そして牧羊犬がドミニコ会士の寓意であることはいうまでもない。
キリスト教徒はやがて羊だけでなく,子牛を使ってたとえられるようになる。16世紀ころのヨーロッパの絵本には,子牛が牧場で跳びはねており,そこに天上から神の2本の手がのばされ,一方の手はむちをにぎり,もう一方の手は香油のつぼをもっているといったたぐいの図柄がしばしば見受けられる。これはむちによる懲らしめと,香油による鎮痛,癒(いや)しということを意味しており,キリスト教徒に対する神の権威と慈愛の両方を象徴したものである。このようにキリスト教徒をたとえるのに,羊よりは牛が好んで使われだしたということは,牛のほうが羊より体も大きくて力も強く,ときには反抗的であることからみて,近代における人間の神に対する自立を暗示しているのかもしれない。
次にヨーロッパにおける馬のもつイメージであるが,ヨーロッパ中世では馬は一方では騎士の乗物であり,それゆえ力強く,しかも高貴な存在とされた。しかし他方では馬は農民,とくに富農の所有物であり,犂耕(りこう)に際しては牛に比べてはるかにスピーディであり力においても勝っていた。それゆえ馬はまた強力なエネルギーのシンボルだったのである。さてこうした強力な馬は近代の蒸気機関の出現の結果無用の存在となる。とはいえ,蒸気機関のみならず電力を使うモーターに至るまでその出力が何馬力(馬力は文字どおり英語のhorse powerの直訳)であるなどといわれるのは,人間が馬のエネルギーを使っていたことの記憶からくるのである。西ヨーロッパ世界は世界中で最初にしかもスムーズに農業社会から工業社会へと移行したのであるが,それは西ヨーロッパの農業が人力に頼らず,牛馬の力に頼っていたからであり,蒸気機関の発明は,牛馬よりももっと便利で強力な動力源を見つけだそうとした努力のあらわれだったといえよう。本来,騎乗を意味するrideや,家畜を追ったり馬車を駆ることを意味するdriveが,自動車などにも使われること,機関車をiron horse(鉄の馬)ということなどにはその間の事情がうかがえる。なお,馬はプラトンからS.フロイトに至るまで情欲のシンボルとしても考えられてきた。仏教にも〈意馬心猿〉ということばがあって統制しがたい私欲のたとえとする。
ヨーロッパ以外の地域でも,狩猟生活,牧畜生活,農業生活に応じて,それに対応する動物イメージが生まれてくる。例えば中国では,狩猟に関しては,〈中原に鹿を逐(お)う〉という表現があり,牧畜に関しては,〈民を牧するは羊を牧すると異ならず〉ということばがある。そして日本には〈牛馬の粉骨〉といういい方がある。しかし中国はともかくとして日本は過去において牧畜になじみが薄かったので,当然のこととして牧畜イメージは皆無に近い。そのかわり日本には狩猟イメージが豊かであり,鹿や猪は神格化されたり,使わしめとなったりした。しかし鹿や猪のイメージも狩猟段階から農業段階に移るにつれて変化し,例えば京都府北桑田郡の諏訪神社での御狩の神事では,神主や氏子が実際に狩りを行い獲物を神に供えるのに対し,京都市の松尾神社での追上げの神事は猪や鹿を里から山に追い返すだけのものとなっている。
執筆者:山下 正男
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
生物を三つの界に大別したとき、植物界、菌界に対し動物界を構成する一群をいう。現在地球上には100万~150万種もの動物が生存しており、形態も生活様式も多種多様である。今後さらに探究され続ければ、少なくとも100万種の動物が新たに発見されるであろうといわれている。
[江上信雄・島田義也]
動物の特徴を、植物との対比でみてみると、次のようなものがあげられる。〔1〕栄養 植物は光合成や窒素固定によって無機分子から有機物をつくりだしエネルギー源とするが(独立栄養)、動物はこのような能力をもたず、植物や動物のつくりだした有機物を餌(えさ)として摂取する(従属栄養)。〔2〕運動 動物は運動器官(鞭毛(べんもう)、繊毛、筋肉と骨格)と神経をもち自由に動くことができる。〔3〕感覚 動物は外界からの刺激を感ずる受容器をもつ。〔4〕器官 動物は、体の各部が消化、排出、呼吸、循環、運動、内分泌、神経、感覚、免疫などの諸器官に分化している。〔5〕細胞壁 植物細胞はセルロースの細胞壁をもつが、動物細胞にはない。〔6〕炭水化物 動物細胞は炭水化物をグリコーゲンの形で蓄えるが、植物細胞はデンプンの形で蓄える。〔7〕細胞分裂 動物細胞は周りがくびれて2細胞に分裂するが、植物細胞は細胞板によって仕切られる。
一般にはこのように、栄養、運動、感覚、器官のほか、細胞レベルでも〔5〕~〔7〕のような特徴が認められる。しかし例外も少なくない。オジギソウは感覚をもち運動を行うのに対し、海綿動物ではあまり顕著ではない。また、ミドリムシは鞭毛をもち運動性に富むが、葉緑体をもって独立栄養である。
[江上信雄・島田義也]
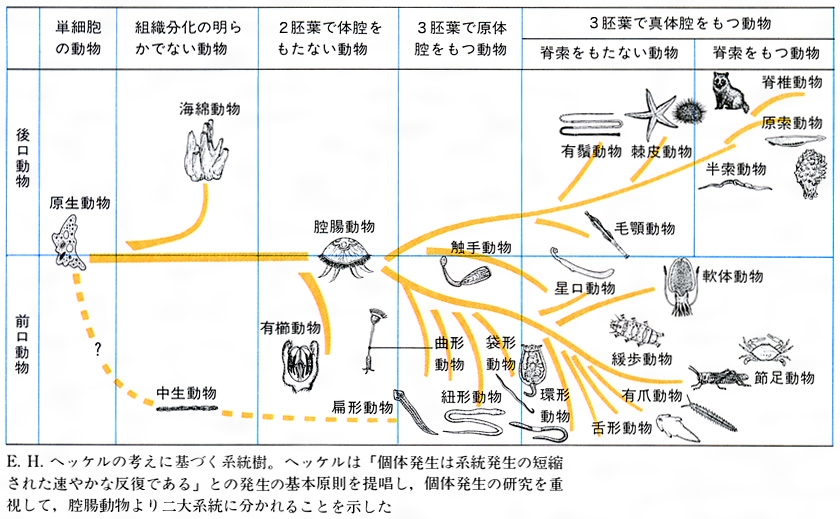
地球上の多くの生物をいろいろな形質の差異や類似性から区別し整理することを分類という。分類の基本単位を種とよび、これはスウェーデンの博物学者C・リンネによって18世紀なかばに確立された。種とは、同一祖先から分化し、同じ形質を備えた繁殖可能な個体群のことをいう。その後、19世紀後半に、イギリスの博物学者C・ダーウィンによって進化論が唱えられ、生物を進化的な類縁関係に従って系統的に分類するようになった。すなわち、化石の研究、比較形態学、比較発生学、核型分析、タンパク質のアミノ酸配列、DNA(デオキシリボ核酸)の塩基配列の比較などによって系統関係が調べられ、系統樹がつくられている。近年の系統樹は、分岐法(一つの種が二つの種に分岐し、それが繰り返されることを示す)によるもの、あるいは幹と枝が離れたものになってきた。これは、種と種の中間的生物の化石が存在しないためで、たとえば、爬虫(はちゅう)類と鳥類の中間的生物とされてきた始祖鳥も、形態の再検討から原始的な鳥の一種であるという説が出ている。この系統分類によっておのおのの生物は、界・門・綱・目・科・属・種にまとめられている。その際、次に記す特徴を系統分類の基準とすることが多い。
〔1〕細胞の分化 単細胞の原生動物、多細胞で組織分化のない中生動物、組織・器官の分化した後生動物に分かれる。〔2〕発生段階 個体発生の段階により次の五つに分かれる。受精卵段階と考えられる原生動物、桑実胚(はい)にあたる中生動物、胞胚段階の海綿動物、嚢胚(のうはい)段階の腔腸(こうちょう)動物、嚢胚以上に発生し、3胚葉からなる器官を分化させた三胚葉動物がそれである。〔3〕消化管のでき方 原口がそのまま将来の口になる前口(旧口)動物(扁形(へんけい)・袋形(たいけい)・紐形(ひもがた)・環形(かんけい)・軟体・節足・触手動物)と、原口は肛門(こうもん)となり将来の口は反対側にできる後口(新口)動物(毛顎(もうがく)・棘皮(きょくひ)・半索・原索・脊椎(せきつい)動物)に分かれる。〔4〕中胚葉のでき方 原則的に、前口動物では、卵割腔に2個の原中胚葉細胞が落ち込み、それが増殖して中胚葉を形成する(原中胚葉幹)。後口動物では、中胚葉は原腸の膨らみから切り出される(原腸体腔幹)。〔5〕幼生の比較 幼生の類似は近縁性を示すと考えられる。環形動物と軟体動物はともにトロコフォラ幼生を経る。棘皮動物のナマコのアウリクラリア幼生は、半索動物のギボシムシのトルナリア幼生と似ている。〔6〕体腔 三胚葉性動物は、体腔が卵割腔に由来する原体腔類と、中胚葉に囲まれた体腔をもつ真体腔類に分かれる。〔7〕脊索・脊椎の存在 これらの有無も分類の重要な基準となる。〔8〕タンパク分子のアミノ酸配列 チトクロムcやヘモグロビンなどのアミノ酸配列の比較によって類縁性が再検討されている。以上のような基準をもとに動物は約15から23の門に分類されている。
(1)原生動物門Protozoa 単細胞で、収縮胞や食胞などの発達した細胞内器官をもつ。群体を形成するものもある。鞭毛、繊毛、仮足(かそく)により運動する。アメーバ、ヤコウチュウなど約5万種。
なお、分類上、原生動物を一門ではなく、動物や植物に匹敵する生物群、あるいはプロチスタ群に属する一群とする考え方もある。
(2)中生動物門Mesozoa 多細胞であるが特別に分化した器官をもたない。ある種の多細胞生物が退化したものと考えられ、原生動物から後生動物へ進化したものとは認めにくい。門としない人もいる。ニハイチュウ(タコの腎臓(じんぞう)に寄生)など約50種。
(3)海綿動物門Porifera 胞胚段階の動物で、地質時代を通じて進化しなかったと考えられる。消化管や筋肉、神経はないが、骨片が分化している。襟細胞や遊走細胞があるので、鞭毛虫から進化したものとみなされる。しかし、嚢胚・三胚葉動物への進化の一過程にあるとは考えられていない。カイメンなど約4500種。
(4)腔腸動物門Coelenterata 体制は放射相称で、内外の2胚葉からなる。嚢胞段階にある。感覚器や筋肉が発達し、中枢のない散在神経をもつ。また、刺細胞をもつのが特徴である。クラゲ、サンゴなど約1万種。
(5)有櫛(ゆうしつ)動物門Ctenophora 腔腸動物と同様に、体制は放射相称で、2胚葉からなる。嚢胞段階にある。刺細胞をもたず、繊毛が集まってできた櫛板をもつ。フウセンクラゲ、クシクラゲなど約100種。
(6)扁形動物門Platyhelminthes 原体腔をもつが、きわめて不明瞭(ふめいりょう)である。消化管は未分化で肛門をもたない。神経系は籠(かご)状である。螺旋(らせん)卵割を行うことから、紐形・環形・軟体動物門との類縁性が示唆されている。最近のリボゾームRNAの研究では、この門は変化に富み、とくにプラナリア類はすべての多細胞動物のうちもっとも原始的なものであるといわれている。プラナリア、肝蛭(かんてつ)など約7000種。
(7)紐形動物門Nemertinea 扁形動物門と多くの点で類似するが、肛門と閉鎖血管系をもち、一部の器官(腸、生殖腺(せん))に体節制がある点で異なる。排出器は原腎管。ヒモムシなど約1000種。
(8)曲形動物門Kamptozoa 一見コケムシ類(触手動物)と似ている。消化管はU字状に曲がり、口と肛門をもつ。循環系はなく、原腎管で排出する。原体腔をもつ。ペディケリナなど約70種。
(9)袋形動物門Aschelminthes 従来の線形動物と輪形動物をあわせて袋形動物とよぶ。体表はクチクラで覆われ、原体腔(擬体腔)が発達している。口と肛門をもつが、寄生性のものは消化系が退化している。ヘテロゴニーheterogony(異常生殖または周期性単性生殖)をするものが多い。ワムシ、回虫など約1万3000種。
(10)軟体動物門Mollusca 閉鎖血管系であり、腎管をもつ。体は頭部、足、内臓塊の3部に分かれる。中枢神経系や中腸腺が発達している。幼生はトロコフォラ、ベリジャーを経る。二枚貝類、ウミウシ(貝殻が退化)、カタツムリ(肺呼吸)、イカ、タコ、アンモナイト(絶滅種)など約10万種。
(11)環形動物門Annelida 体は、同じような構造をもった多数の体節(等体節)からなる。真体腔が発達し、体壁には環状筋と縦走筋がある。排出器は腎管(体節器)で、梯子(はしご)状神経節、閉鎖血管系をもつ。ゴカイ、ミミズ、ユムシなど約7000種。
(12)有爪(ゆうそう)動物門Onychophora 脚(あし)の末端に2個の鉤(かぎ)づめをもつ。粘液腺をもち、攻撃、捕食に用いる。呼吸器官として気管をもつ。開放血管系で、排出系は腎管である。カギムシなど約80種。
(13)緩歩動物門Tardigrada 後生動物としては最小で、体長は1ミリを超えるものはない。梯子状神経系をもつ。体表はクチクラで覆われ、循環系と呼吸系はない。乾燥させると体は収縮し、乾眠状態になり、4~7年は耐える。これを湿らせると、ただちに回復し、活動する。乾眠状態では100℃の高温や零下250℃の低温にも耐え、放射線照射や高圧、真空状態にも強い。クマムシなど約280種。
(14)舌形(したがた)動物門Linguatulida 脊椎動物に内部寄生する。体表はクチクラで覆われ、循環器を欠き、神経系の発達も悪い。シタムシなど約60種。
(15)節足動物門Arthropoda 動物界最大の門で、全動物種の7、8割を占める。体は、部分により分化した体節(不等体節)からなる。キチン質の外骨格があり、梯子状神経管、開放血管系をもつ。腎管またはマルピーギ管で排出する。脱皮、変態をするものが多い。エビ・カニの甲殻類(幼生はノープリウス、ゾエアなどを経る)、クモ・ダニ・トンボなどの昆虫、サンヨウチュウ(絶滅種)など約100万種。
(16)星口(ほしぐち)動物門Sipunculoidea 細長い吻(ふん)をもつ。体節制はない。排出器は一対の腎管である。トロコフォラ幼生を経る。ホシムシなど約280種。
(17)触手動物門Tentaculata 原腸体腔幹であるが前口動物。ある種の腕足類(古生代に繁栄した生きた化石)の幼生は毛顎動物と似ている。コケムシ類、シャミセンガイやホオズキガイなどの腕足類など現存は約5000種、化石には約2万種以上ある。
(18)毛顎動物門Chaetognatha 目や、剛毛の生えた口をもつ。循環・排出系はないが、体腔のでき方は棘皮動物、原索動物に似る。プランクトンのヤムシなど約60種。
(19)有鬚(ゆうしゅ)動物門Pogonophora 左右相称で、体は前体、中体、後体の3部に分かれる。前体には触手冠と脳がある。クダヒゲムシなど約70種。
(20)半索動物門Hemichordata 脊索と相同と考えられる盲管と、咽頭(いんとう)に鰓裂(さいれつ)をもつ。神経系が背側にあり、前述(1)~(19)の無脊椎動物と異なる。幼生のトルナリアは、ナマコの幼生アウリクラリアに似る。ギボシムシなど約100種。
(21)棘皮動物門Echinodermata 幼生は左右相称であるが、成体は放射相称となる。水管系、内骨格をもち、管足で移動する。ヒトデ(ピピンナリア幼生を経る)、ウニ(プルテウス幼生)、ナマコ(アウリクラリア幼生)など約6500種。
(22)原索(脊索)動物門Prochordata 発生のある時期または一生、脊索をもつ。鰓裂、内柱があり、神経管が発達する。ホヤなどの尾索綱と、脊椎動物の原型と考えられる体制(発達した脊索と神経管)をもつナメクジウオの頭索綱に分かれる。約1600種。
(23)脊椎動物門Vertebrata 発生初期に脊索を生じ、のちに脊椎に置き換わる。脳が分化している。消化系、循環系、呼吸系などの器官系が発達する。胚が水中で発生する魚類と両生類では、胚は羊膜に包まれていない(無羊膜類)。胚が陸上で発生する爬虫類、鳥類、哺乳(ほにゅう)類の胚は羊膜に包まれて保護されている(羊膜類)。鳥類、哺乳類は恒温動物で、カモノハシなど一部を除いて哺乳類は胎生である。約4万3000種。
なお、分類上、原索動物と脊椎動物についてはそれぞれを門とせず、脊索動物を1門とし、尾索動物、頭索動物、脊椎動物を3亜門としてこの門に含める考え方や、このうち脊椎動物亜門を設けず、無顎類と顎口類の2亜門を加えて合計4亜門とする研究者もある。
[江上信雄・島田義也]
地球上に最初に生物が現れたのは30億~35億年前のことである。南アフリカではそのころの岩石から、細菌や細菌の祖先であるエオバクテリウムの化石がみつかっている。原始大気中のメタン、水素、アンモニア、水蒸気などが、紫外線や空中放電によってアミノ酸となり、さらにタンパク質がつくられた。それに脂質などのコロイド粒子が結合し、周囲の海水から独立したコアセルベートになった。コアセルベートが酵素や核酸を取り込んで代謝や自己増殖能を獲得することにより、生物が生まれたと考えられている。その後、核をもった細胞(真核細胞)が現れ、菌類や植物や動物に分化していったと思われる。また、単細胞動物の鞭毛虫が群体をつくり、一部が分化して多細胞動物が進化してきたと考えられている。
地質時代は、化石をもとにした生物の進化過程に大きな段階の認められるところを境にして、先カンブリア時代(約46億~5億7500万年前)、古生代(5億7500万~2億4700万年前)、中生代(2億4700万~6500万年前)、新生代(6500万年前~現在)の四つに分けられる。先カンブリア時代には、藍藻(らんそう)類が増加し、末期には海産無脊椎動物が出現した。古生代に入ると、動物の化石が豊富にみつかっている。古生代の前半には、節足動物の三葉虫、触手動物の腕足類、軟体動物のオウムガイ、腔腸動物のサンゴ、棘皮動物のウミユリなど、海産無脊椎のほとんどの門の動物が繁栄し、やがて魚類が出現した。三葉虫は世界に広く分布し示準化石となっている(現在のカブトガニは三葉虫と似た体制をもつ生きた化石である)。古生代中期には魚類が繁栄し、陸地の拡大と高温多湿な気候により植物が上陸した。続いて魚類のなかに、肺魚や総鰭(そうき)類(シーラカンス)などのように、直接肺に空気を送ることができ、じょうぶなひれで歩くことのできるものが出現し、進化して両生類となった。古生代後半には、陸生の昆虫の繁栄とともに、羊膜をもつ卵を産む爬虫類が出現した。また原生動物のフズリナも急激に増加した。古生代末期には、地殻の変動や氷河の発達によって多くの生物が絶滅してしまった。しかし、生き残った生物は中生代の温暖で乾燥した気候のなかで急に発展した。とくに爬虫類が全盛となり恐竜が繁栄した。続いて哺乳類や鳥類(始祖鳥)も現れた。海中には、示準化石となっている軟体動物のアンモナイトが爆発的に進化した。しかし多くの生物は新生代には絶滅した。新生代には、現存のすべての生物の祖先が現れ、とくに哺乳類が広く適応放散し、やがて人類が出現し発展するようになった。
[江上信雄・島田義也]
動物は、従属栄養、すなわち他の生物が合成した有機物を栄養とするため、消化系、循環系、排出系が体内に発達し、高等動物ではさらに運動系、感覚系、神経系が発達している。これらの器官系は、上皮組織、結合組織、筋肉組織、神経組織からなる。〔1〕上皮組織 体の外表面、消化管や血管の内表面を覆っていて、内部組織の保護や、吸収、分泌、感覚、生殖細胞の形成などの働きを行っている。〔2〕結合組織 組織や器官の間を満たし、それらを結合したり支持したりし、多量の細胞間物質を含む。腱(けん)や靭帯(じんたい)、腸間膜、軟骨、骨組織、血液、リンパなどを構成している。〔3〕筋肉組織 収縮性に富み、横紋筋と平滑筋がある。横紋筋は骨格筋や心筋にみられ、節足動物の運動器官にもあり、急激な運動に適する。二枚貝の貝柱の横紋は斜めであり斜紋筋とよばれる。〔4〕神経組織 神経細胞と神経膠(こう)から構成され、刺激の伝達を行う。一般に脊椎動物の神経突起は、髄鞘(ずいしょう)と神経鞘という二重の鞘(さや)に包まれ(有髄神経)、刺激の伝達は速い。無脊椎動物の神経の多くは髄鞘がなく(無髄神経)、刺激の伝達は遅い。
以上の四つの組織が組み合わさって、いくつかの器官をつくっている。高等動物では、多数の器官が集まって、次のような器官系をつくり、全体としてまとまった働きをする。
(1)消化系 高等動物は細胞外消化を行うが、原生動物や海綿動物の襟細胞は、細胞内に食胞をつくり細胞内消化を行う。腔腸動物や扁形動物は肛門がない。扁形動物より高等な動物では消化器官が発達し、ある動物は特有の器官をもつ。たとえば、ミミズは嗉嚢(そのう)や砂嚢、カタツムリは歯舌(しぜつ)をもつ。寄生や共生によって消化器官が退行する例が多い。
(2)呼吸系 下等な動物は体表を通してガス交換を行うが、体制が複雑な高等動物では呼吸器官が発達している。原生・海綿・腔腸・扁形・陸生の環形動物は、体表から酸素を取り込んでいる。水生の環形・軟体・節足(甲殻類)・原索動物、そして脊椎動物のうち魚類や両生類の幼生(オタマジャクシ)は、えら呼吸をする。トンボの幼虫のヤゴやドジョウは腸呼吸もできる。肺魚は乾期になるとうきぶくろで呼吸ができる。一方、陸生の節足動物は気管呼吸をする。昆虫類は、脊椎動物と異なり、ガスの運搬は気管系、栄養分は血管系という分業を確立している。肺呼吸は脊椎動物にみられる。
(3)循環系 栄養分の運搬やガス交換を行う。温度や塩濃度が一定な海中にすむ小動物は循環系をもたないことが多い。体が大きくなるにつれ、体内に海水を入れる水管系を発達させ(クラゲ)、さらに進化して体液を体内に閉じ込める体制を確立し、陸上生活を可能とした。血管系には、開放血管系(節足・軟体動物)と閉鎖血管系(環形・脊椎動物)がある。脊椎動物の循環系も進化に応じてすこしずつ異なる。魚類の心臓は1心房・1心室。両生類、爬虫類は2心房・1心室で、肺循環と体循環の区別がある。鳥類、哺乳類は2心房・2心室で、肺循環できれいになった血液が体循環する。酸素を運ぶ呼吸色素には、鉄を含む赤いヘモグロビン(脊椎動物、ナマコ、ミミズ)や銅を含むヘモシアニン(甲殻類、軟体動物)などがある。
(4)排出系 水と老廃物を排出する器官系で、多くは浸透圧調節も兼ねる。原生動物では細胞内器官の収縮胞が、原体腔をもつ動物では原腎管が、真体腔をもつ動物では腎管が排出器である。軟体動物のボヤヌス器や甲殻類の触角腺はともに腎管の変化したものである。昆虫はマルピーギ管、線虫類は側線管を発達させている。脊椎動物の排出器官は腎臓で、高等になるほど、前腎・中腎・後腎と分化してゆく。タンパクの分解物で細胞に有害なアンモニアは、無脊椎動物や硬骨魚類ではそのままの形で、軟骨魚類、両生類や哺乳類はアンモニアを尿素に変えて、爬虫類、鳥類、昆虫は尿酸に変えて排出する。
(5)内分泌系 脊椎動物の場合、下垂体、甲状腺、副腎、生殖腺などの内分泌器官から分泌された種々のホルモンは、エネルギー生産の調節、成長や変態の促進、生殖活動および体内環境の調節などの恒常性の維持を行う。無脊椎動物には、性ホルモンのほか、体色変化・脱皮調節のホルモン(甲殻類のサイナス腺)、変態調節のホルモン(昆虫のアラタ体や前胸腺)がみつかっている。
(6)神経系 神経系は海綿動物にはないが、腔腸動物より高等な動物にある。腔腸動物のヒドラの神経細胞は全身に散在し、散在神経とよばれる。伝達速度は遅い。扁形動物以上の神経細胞は神経節をつくっている(集中神経系)。クラゲや棘皮動物の神経節は環状につながっているので環状神経節、環形・節足動物の神経節は対(つい)になっていて、梯子状神経節という。原索動物と脊椎動物の神経系は、神経節が管状で体の中軸に集まり、管状神経節とよばれ、脳が発達している。
(7)感覚系 光、音、重力、化学成分、温度、圧力などの刺激を受容する。視覚を例にとると、ミドリムシには色素が集まった眼点があり、ミミズは体表の視細胞が光を感じる。アワビはピンホール式の杯状眼を、タコはカメラ眼をもつ。節足動物は複眼と単眼をもつ。
(8)免疫系 個体維持のため、細菌や老朽細胞に対する防御反応をつかさどる器官系。脊椎動物では、骨髄、胸腺(哺乳類)、ファブリキウス嚢(鳥類)、脾臓(ひぞう)や腎臓、無脊椎動物では、腎臓や腸間膜、消化管などの諸器官に付随する造血器やリンパ組織で、マクロファージや高等な動物では特殊化されたリンパ球が免疫担当細胞として分化する。無脊椎動物ではレクチン、補体、ある種の抗菌物質が防御因子として働き、脊椎動物では免疫グロブリンが現れる。海綿動物の同種細胞の凝集反応、腔腸・環形・棘皮・原索動物の異種および同種移植片拒絶反応は免疫反応の一種である。
そのほか、運動・生殖・骨格系の器官が各動物に発達している。特殊器官としては、横紋筋が変化したシビレエイの発電器、求愛のシグナルとなるホタルの尾部発光器などがある。
[江上信雄・島田義也]
生殖とは新しい個体をつくりだすことで、無性生殖と有性生殖がある。無性生殖は下等動物の一部にみられ、大部分の動物は有性生殖を行う。しかし、クラゲやウミタルは、有性生殖と無性生殖を交互に繰り返すメタゲネシスmetagenesis(真正世代交代)を行う。二分裂による増殖は原生動物やイソギンチャク、出芽による増殖はカイメン、ゴカイ、コケムシ、胞子により増殖する方法はマラリア原虫にみられる。有性生殖の確立によって、遺伝子の新しい組合せをもつ個体が発生し、変化する環境により広く適応できる可能性が生まれた。ゾウリムシは二つの個体が接合し核を交換する。高等動物では、卵と精子の合体によって新個体が生まれる。多くの動物は雌雄異体であるが、ミミズやカタツムリは、一つの個体内に卵と精子をつくる雌雄同体である(しかし、自家受精は一般に行わない)。ミツバチやワムシでは、有性生殖のほか、卵が受精をしないで単為発生して新個体が生じ、タマバエや肝蛭は、幼生体内の卵が単為発生を始める幼生生殖を行う。ワムシのように有性生殖と単為生殖を繰り返すことをヘテロゴニー、タマバエのように有性生殖と幼生生殖を繰り返すことをアロイオゲネシスalloiogenesis(混合生殖)とよぶ。
有性生殖を行う動物では、配偶者をうまくみつけるためのいろいろの仕組みがある。たとえば、昆虫では一般に雄が目だつ色彩や形態をもち、雌は性フェロモンを分泌するものが多い。ある種の鳥は、求愛のさえずりやダンスなど一連の配偶行動をする。
一方、発生とは、受精卵が細胞分裂を繰り返し、分化し、組織・器官を形成し、胚や幼生を経て成体となる過程をいい、胚発生には各動物共通の点が多い。受精卵は、卵割とよばれる細胞分裂をして細胞数を増やす。卵割を続けた卵は、内部に大きな空所(卵割腔)をもつ胞胚になる。その後、一部が内側に向かって陥入し原腸をつくり、嚢胚となる。原腸はのちに消化管となる。嚢胚をつくる壁を胚葉といい、外側の外胚葉、内側の内胚葉に分かれ、腔腸動物より高等な動物では、さらに中胚葉が分化する。外胚葉は表皮や神経に、中胚葉は脊索や骨、筋肉、腎臓、生殖器に、内胚葉は消化管や内分泌腺、肺やえらに分化してゆく。
発生中の胚は環境の変化や外敵に弱いので、下等な動物は多くの卵を産み、種属を維持している。脊椎動物の爬虫類や鳥類などの陸上に産卵するものは乾燥を防ぐため、胚を保護する胚膜が形成されている。哺乳類では、胚発生は母体内で進行し、胎盤を通して直接母体から栄養を受け取る胎生が発達している。このような胚膜の形成や胎生の発達が、動物の生活圏を水中から陸上へ広げたと考えられる。
[江上信雄・島田義也]
動物の地理的分布は、哺乳類、鳥類をもとにして、旧北区(ヨーロッパ、アジアの大部分、アフリカ北部)、新北区(北アメリカ大陸とグリーンランド)、エチオピア区(サハラ砂漠以南のアフリカ、アラビア半島)、オーストラリア区(オーストラリア、ニューギニア島)、新熱帯区(南アメリカ、中央アメリカ、西インド諸島)、東洋区(中国南部、インド、東南アジア)に区分されている。
エチオピア区と東洋区には、サイ、ゾウ、旧世界ザル、大形のネコ科動物など似た類が多い。これは、新生代第三紀まではアフリカ大陸とインドが陸続きであったことと関係している。オーストラリア区には、カンガルー、コアラなどの有袋類、カモノハシ、ハリモグラなどの単孔類、ゴクラクチョウなど特異的な動物が多い。大陸移動説によれば、古生代には一続きであった陸地(ゴンドワナ大陸)が中生代に移動し始め、単孔類と有袋類以外の哺乳類が栄えていない新生代第三紀にオーストラリア大陸が分離した。その後、北の大陸ではより高等な動物が進化し前述の動物は絶滅したが、大陸から離れたオーストラリアでは滅びることなく生き残ったと考えられる。もうすこし早く大陸から分離したマダガスカル島やガラパゴス諸島に特異的な生物がみられることも進化を考えれば理解できる。東洋区とオーストラリア区の境界をワラス線(ウォーレス線)とよぶ。
[江上信雄・島田義也]
日本は大部分が旧北区で、沖縄以南が東洋区に属す。この境界が渡瀬(わたせ)線で種子島(たねがしま)と奄美(あまみ)大島の間に引かれる。本州と北海道もブレーキストン線で分かれる。北海道にはヒグマ、ナキウサギ、エゾライチョウなどシベリアと同種の動物がいる。本州は中国の動物相と似ていて、ツキノワグマ、シカ、キジがいる。また、両生類と無脊椎動物の分布から宗谷(そうや)海峡に境界を置く八田(はった)線もある。対馬(つしま)は、アカネズミやツシマテンなどの日本本土と共通の種と、チョウセンモグラ、キタタキなど朝鮮半島と共通の種がいることで、動物分布の点から興味深い。最近、DNAの塩基配列や酵素の多型を調べることによって、本州の動物(ネズミ、メダカ)も北型と南型に分かれるものがあることが明らかとなった。
[江上信雄・島田義也]
動物の特性の一つは、環境からの刺激を感じ、反応し、行動することである。刺激に対し一定の方向に行動することを走性という。刺激には、光、重力、水流、温度、化学物質などがある。ゾウリムシは重力と反対の方向へ、また電気に対しては陰極へ移動する。昆虫や魚の多くは光の方向へ、ミミズは光を避けるように移動する。高等な動物では、脳が発達し、その行動は本能行動、学習行動、知能行動などに分けられる。経験や学習なしに生まれつきの性質として遺伝的に決まった行動を本能行動という。たとえばミツバチは分業して巣づくりと育児をする。また、臀(しり)振りダンスをして餌の所在を知らせる。カイコガの雄は、雌の分泌する性フェロモンにひかれて雌に近づく。サケは、生まれた川をさかのぼって産卵する。トゲウオの雄は巣をつくり求愛のダンスをするが、これも本能行動である。またガチョウの雛(ひな)は、最初に出会う動くものを親と認知し、ついて歩く。この認知能力は孵化(ふか)直後の特定の時期にのみみられ、刷り込みという。脳の発達した動物では、さらに経験に基づく行動、すなわち条件反射、学習、知能行動が可能となる。条件反射は、イヌを使った当時のソ連の生理学者パブロフの実験が有名であるが、タコでも条件づけの実験に成功している。学習では、経験したことを記憶することが重要である。マウス、タコ、ミツバチ、ミミズなど広い動物群で学習の効果がみられる。大脳皮質の発達した哺乳類は、経験や学習をもとに、未経験のことに対して、その事態に適した新しい行動をする能力(知能)をもっている。
[江上信雄・島田義也]
生物と自然環境は互いに作用しあい、また生物間も捕食や寄生、共生などの関係で結び付いている。
多くの動物は普通は同じ仲間で群集をつくって生活をしている。いつもは単独で生活していても、生殖期(ウニ、ヒトデ)、移動期(バッタ)、冬眠時(ヘビ、コウモリ)に群集をつくるものがいる。個体群の移動は季節に依存し、餌や生殖などの環境を求めて行う。一定地域内の個体群の動物では、個体間に優劣の順位があったり(サル、カラス、メダカ)、リーダーがいて秩序のある社会集団を形成する場合(ニホンザル、アリ、ミツバチ)がある。また、縄張り(テリトリー)があって餌の捕獲や繁殖に役だつ。
一方、同じ環境内にすむ異なった動物の間に、お互いに影響を与え合っている例がある。その場合の利害関係は食物とすみ場所の獲得であることが多い。生存競争の結果、多くの場合一方が消えていく。しかし、魚のイワナとヤマメは同じ川でも上流と下流に分かれて生活し、餌の競争を避けている(すみ分け)。カワウとヒメウは同じ場所にすむが、とる餌が異なり競争を回避している(食い分け)。そのほか、個体群の相互作用には、お互いまたは一方が利益を受ける共生関係がある。アリとアブラムシはともに利益を受ける相利共生、サメとコバンザメは後者のみ利益を受ける片利共生をしている。ダニや回虫は動物に寄生している。生物間でもっとも重要な関係は捕食関係である。捕食者と被食者の関係は網の目のように複雑である。植物は草食動物に食べられ、草食動物は肉食動物に食べられる。その肉食動物はさらに大形の肉食動物に食べられるというような食物連鎖がみられる。動物の死体や排出物は、カビや細菌によって分解されて、食物連鎖の源である植物の養分となり、生態系のなかでは栄養物質の循環関係がみられる。普通、生態系を構成している生物の種類組成や個体数などはつり合いがとれている。
[江上信雄・島田義也]
安定している生態系も、火山の噴火や台風などの自然災害によって平衡が破れる。しかし、もっとも影響が大きいのは人間による人為的破壊である。大気・水質・土壌の汚染、ダム建設による生息地の破壊や乱獲によって絶滅した動物は少なくない。最近400年間にオーロックスやニホンオオカミなど30~40種の哺乳類、リョコウバトやドードーなど100種の鳥類が滅び、さらに数百種の動物(ジャワサイ、イリオモテヤマネコ、トキなど)が滅びつつあるといわれる。また、ウシガエルの餌として輸入したアメリカザリガニのように、原産地から移動し、他の地区に適応し繁殖した帰化生物も生態系を破ることが多い。これは天敵がいないため、急激に数が増え、植物や下位の動物を食い荒らすからである。
近年、自然の生態系の安定化を図るため、環境の保全と自然の保護に対する要求が高まってきた。その結果、自然公園の設定や野生動物捕獲禁止の法律を制定し、絶滅の危機に瀕(ひん)している動物を天然記念物に指定して保護している。
[江上信雄・島田義也]
『石田寿老他著『現代動物学』(1972・裳華房)』▽『江上信雄・飯野徹雄著『生物学』上下(1984・東京大学出版会)』▽『アテンボロー著、日高敏隆他訳『地球の生きものたち』(1983・早川書房)』▽『江上信雄他著『UPバイオロジー・シリーズ』(1975~ ・東京大学出版会)』▽『内田享監修『動物系統分類学』全10巻、17冊(1961~ ・中山書店)』▽『内田享他監修『現代生物学大系』全14巻(1965~ ・中山書店)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
字通「動」の項目を見る。
出典 平凡社「普及版 字通」普及版 字通について 情報
Sponserd by ![]()
…生物界を動物と植物に二大別するのは,常識の範囲では当然のように思えるが,厳密な区別をしようとするとさまざまな問題がでてくる。かつては生物の世界を動物界と植物界に二大別するのが常識だったが,菌類を第三の界と認識すると,それに対応するのは狭義の動物(後生動物),狭義の植物(陸上植物)ということになり,原生動物や多くの藻類などは原生生物という名でひとまとめにされ,また,これら真核生物に比して,細菌類やラン藻類は原核性で,原核生物と別の群にまとめることができる。…
…いずれにせよ,エネルギー転換にかかわるエネルギーのシステムと,それの方向を指示し,自己保存,自己増殖を行う情報のシステムとが組み合わさったとき,初めて,生物が生じたことは確かである。
[生物の分類]
生物は動物と植物に大別されるが,これは外界からの物質やエネルギーのとりこみ方の違いに基づくものである。すなわち,主要なエネルギー源として太陽エネルギーを,体を構成する物質源としては無機物質をとりこむ方式で生きるものが植物であり,エネルギー源,物質源を有機物質に頼るのが動物であると考えてよい。…
※「動物」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
乞巧奠〈公事十二ケ月絵巻〉〘 名詞 〙 陰暦七月七日の行事。乞巧は技工、芸能の上達を願う祭。もと中国の行事であるが、日本でも奈良時代以来、宮中の節会(せちえ)としてとり入れられ、在来の棚機津女(たなば...