
内科学 第10版 の解説
甲状腺ホルモン・ヨウ素代謝(甲状腺)
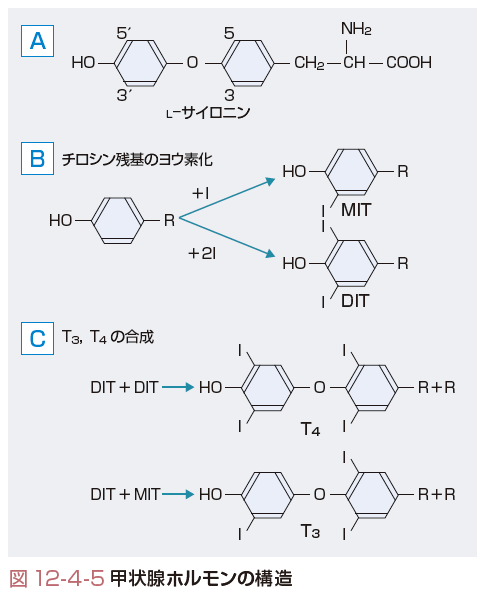
a.甲状腺ホルモンの構造
甲状腺ホルモンは,l-サイロニン(l-thyronine)を基本骨格とするヨウ素化アミノ酸である.図12-4-5に示すようにl-サイロニンに結合するヨウ素の数と部位により,3,5,3′,5′に4個のヨウ素の結合したテトラヨードサイロニン(T4, thyroxine)と3,5,3′に3個のヨウ素の結合したトリヨードサイロニン(T3),その他リバースT3(rT3),T2などがある.甲状腺から分泌されるのはおもにT4で,T3は全体の5~10%であるが,強い活性を有するのはT3で,末梢では後述するように脱ヨウ素反応でT4からT3へと変換される.rT3
はほとんど活性がない.
b.甲状腺ホルモンの合成と分泌
1)サイログロブリンの合成と貯蔵:
甲状腺ホルモンの合成は,食事より摂取されたヨウ素を原料とし,甲状腺特有の蛋白質であるサイログロブリンの分子内のチロシン残基で行われ(図12-4-3),コロイドとして濾胞腔内に貯蔵される.サイログロブリンは分子量約66万の高分子糖蛋白質で1分子中のチロシン残基数は120~130で,その約25%がヨウ素の受容体として機能する(図12-4-3).
2)ヨウ素の取り込みと濃縮:
食事中に含まれる無機ヨウ素は腸管より吸収され,甲状腺の濾胞上皮細胞により摂取される.この甲状腺によるヨウ素の摂取は,おもにNa+-I-共輸送体(NIS)とよばれる細胞膜を12回貫通するトランスポーターが関与する.摂取されたヨウ素は,25~100倍に濃縮される(図12-4-3).このヨウ素の取り込みは,下垂体からのTSHの刺激により促進され,チオシアン酸塩あるいは過塩素酸塩などの無機イオンで抑制される.チオシアン酸塩は,キャベツなどの植物に多く存在する.
3)チロシン残基のヨウ素化:
取り込まれて濃縮されたヨウ素は,濾胞腔内でH2O2と細胞膜に局在する甲状腺ペルオキシダーゼ(thyroperoxidase:TPO)の存在下で酸化され,I2となりサイログロブリンのチロシン残基と結合する(ヨウ素の有機化).この際のヨウ素の濾胞内への放出にPendred症候群の原因遺伝子産物であるPendrin(PDS)が関与している(図12-4-3).このヨウ素の有機化の結果,3モノヨードチロシン(MIT)が生成され,さらに十分のヨウ素があると3,5ジヨードチロシン(DIT)の生成へと進む(図12-4-5).このヨウ素の有機化はTSHによって促進され,抗甲状腺薬,スルホンアミド,大量のヨウ素,チオシアン酸塩で阻害される.
4)MITとDITからのT4,T3の合成:
図12-4-5に示すように,サイログロブリンと結合しているMITあるいはDITが酸化され,アラニン残基を失って縮合し,T3基およびT4基が生成される.この縮合反応にもTPOとH2O2系が関与する(図12-4-3).この反応は,抗甲状腺薬,スルホンアミド,および大量のヨウ素により抑制される.
上記過程を経て生成されたT3基,T4
基は,コロイドとして甲状腺濾胞内に貯蔵される.コロイド内には体内で必要とされる約1カ月分の甲状腺ホルモンが貯蔵される.
5)サイログロブリンの再吸収,加水分解とT4,T3分泌:
甲状腺がTSHなどにより刺激されると貯蔵されていたサイログロブリンは,コロイド小滴として濾胞上皮細胞内へ再び取り込まれ,プロテアーゼで加水分解されT3ならびにT4が形成され,エクソサイトーシスにより血中に分泌される.この加水分解の際に,サイログロブリン分子内のヨウ素チロシン基も同時に遊離される.さらに脱ヨウ素化され,遊離したヨウ素の一部は血中へ放出され,またほかの一部は甲状腺ホルモン合成に再利用される.甲状腺から分泌されるT4は,ヒトでは1日約100 μgである(図12-4-3).
6)甲状腺ホルモンの血中存在様式:
血中に分泌されたT4やT3は脂溶性であるため,ほとんどがサイロキシン結合グロブリン(thyroxine-binding globulin:TBG)に結合している.しかし,実際に生物学的活性を有しているのはTBGに結合していない遊離ホルモンである.TBGのほかにも,トランスサイレチン(thyroxine-binding prealbumin:TTR)とアルブミンなどにも結合する.T4の75%はTBGに結合しており,ほかの約25%はTTRやアルブミンと結合しており,0.02~0.03%が遊離型で存在する.T4はTBGに親和性が強く,TBG 1分子にT4の1分子が結合する.T3も親和性は弱いがTBGと結合している.T4の血中半減期は約7日であり,T3は2日と短い.血中の総T4,T3値を測定した際,その値はこれらの蛋白質に結合したT4,T3も測定しており,特に妊娠時やエストロゲン投与時,急性の肝機能障害時などではTBGの値が増加し,またネフローゼ症候群,大量ステロイド剤投与時,肝硬変症などではその値が低下するので,T4値の解釈には注意が必要である.このような際にも遊離T4,遊離T3値は影響を受けにくい.TBGの異常としてはTBG欠損症があり,またTTRの異常としては家族性アミロイドポリニューロパチーがある.
7)細胞内,核内への移動:
末梢へ到達したT3,T4は,細胞膜そして核膜を貫通して甲状腺ホルモン受容体へと到達する.甲状腺ホルモンが細胞膜を貫通する際は,有機アニオンやモノカルボン酸が甲状腺ホルモンのトランスポーターとしても機能している.そのなかでも,monocarboxylate transporter 8の異常では,血中甲状腺ホルモンは高値を示し,神経発達の異常を伴う.
(2)ヨウ素代謝
正常な甲状腺ホルモン産生に必要なヨウ素の量は,1日100~200 μgで,それは海草類,特に昆布に多く含まれている.ヨウ素の摂取不足により甲状腺ホルモン合成能が低下し,甲状腺腫を伴う甲状腺機能低下症になるが,日本などのヨウ素の摂取量が多い国ではまれである.しかし,大量のヨウ素摂取は逆に甲状腺ホルモン合成を抑制する(Wolff-Chaikoff効果).ヨウ素を含む造影剤やうがい薬,薬剤としては抗不整脈薬のアミオダロンなどの投与により甲状腺機能障害を起こすことがあるので注意が必要である.
(3)甲状腺ホルモンの代謝
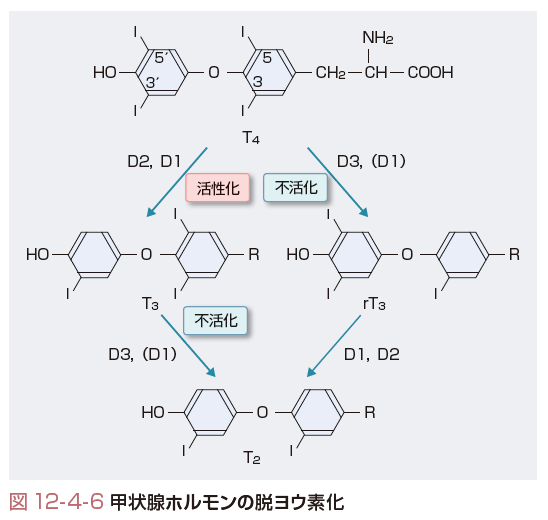
T3の約80%は,末梢組織でT4の5′の位置が脱ヨウ素反応(5′-deiodenase)が起きて生成される(図12-4-6).一方,甲状腺より分泌されたT4の約40%は,5位の脱ヨウ素反応(5-deiodenase)により活性を有さないリバースT3(rT3)に変換される.これらのT3およびrT3は,さらに脱ヨウ素反応によりT2からT1へと代謝される.したがって,甲状腺ホルモンの作用を考える上ではT4から活性型T3への変換および不活性型rT3への変換も考慮しなくてはならない.脱ヨウ素化する酵素にはタイプ1~3の3種類がある.いずれの酵素もまれなアミノ酸であるセレノシステインを活性中心にもち,アミノ酸レベルで,相互に約50%の相同性をもつ.
タイプ1(D1)はT4から活性型のT3へ,さらにT3から3,3′ T2へと変換する酵素で,肝臓,腎臓,甲状腺などに発現している.この酵素活性は,抗甲状腺薬であるpropilthiouracil(PTU)で阻害され,甲状腺ホルモンにより刺激される.Basedow病では甲状腺のD1活性が増加し,T4の分泌とともにT3の分泌も増加する.さらに,末梢組織でもT4からT3への変換が増加し,血中T3の約70%はD1活性によるT4からの変換であるとされ,血清T4値よりT3値の増加が顕著になる.
タイプ2(D2)は,タイプ1同様T4からT3への変換も行うが,5位の脱ヨウ素反応は示さず,rT3がT2へと転換する.D2は,下垂体や脳,血管内皮,筋肉などに発現しており,甲状腺ホルモン値が低下すると酵素活性は増加し,末梢での活性型T3の濃度を制御している.甲状腺機能低下状態での末梢T3
産生の約70%はD2活性による. 一方,タイプ3(D3)は,脳や胎盤,皮膚などの限られた臓器で発現し,5位の脱ヨウ素反応を行い,甲状腺ホルモンを不活性化する作用をもつ.甲状腺ホルモンが高いとその酵素活性が増加する.肝臓血管腫などで本酵素が大量に発現すると甲状腺機能低下症が生ずる.また,先天的に本酵素が欠乏するマウスは中枢性甲状腺機能低下症を示す.
(4)甲状腺ホルモン作用
甲状腺ホルモンは,核内受容体である甲状腺ホルモン受容体に結合し,標的遺伝子の転写制御を行う(genomic,核内)作用と遺伝子調節作用を介さない(non-genomic,核外)作用をもつ.現在明らかとなっている甲状腺ホルモンの作用のほとんどが遺伝子調節を介する作用である.甲状腺ホルモンにより制御される遺伝子は多岐にわたり,その総合的な生体内作用として以下のような作用を示す.
1)熱産生作用:
酸素消費を増加させ,基礎代謝率を高めることによりカロリー消費を促す.甲状腺中毒症では基礎代謝率が亢進し,甲状腺機能低下症では低体温や基礎代謝率の低下を示す.
2)成長に関する作用:
脳や骨の発育に必須である.先天的に甲状腺ホルモンが不十分な場合は知能遅延を示すクレチン症となる.
3)脂質代謝に関連した作用:
肝臓でのLDL受容体を増加させ,コレステロール合成を低下させ,その胆汁酸排泄を促し,結果として血清コレステロール値を低下させる.甲状腺中毒症では,低コレステロール血症を,甲状腺機能低下症では高コレステロール血症を示す.このため潜在性の甲状腺機能低下症は動脈硬化の危険因子となる可能性がある.
4)糖代謝に関連した作用:
腸管からの糖の吸収を促進し,インスリン分泌を低下させ,食後の急激な血糖値の上昇を示すことがある.
5)蛋白質代謝に関連した作用:
特に肝臓や筋肉における種々の酵素などの遺伝子の発現を増減する.適切な濃度の甲状腺ホルモンは,蛋白質合成の維持に重要であるが,過量になると蛋白質異化作用が強くなる.
6)自律神経への作用:
交感神経β受容体の増強作用や心筋のミオシン重鎖遺伝子発現亢進などにより心拍数が増加する.甲状腺中毒症では頻拍が,機能低下症では徐脈が認められる.
7)皮膚への作用:
グリコサミノグリカンの皮下組織への沈着を抑制する.甲状腺機能低下症では,粘液水腫様顔貌や四肢でのnon-pitting edemaが認められる.
8)甲状腺ホルモンによる
non-genomic(核外)作用:
核外作用については,ミトコンドリアへの作用,PI3K-Akt/PKBを介した作用,カルシウムATPase,アデニル酸シクラーゼへの作用など多くの報告がある.
(5)甲状腺ホルモン作用機構
甲状腺ホルモン受容体は,図12-4-7に示すような共通の構造を示す核内受容体スーパーファミリーに属する.甲状腺ホルモン受容体には,2つの異なる遺伝子から由来するTRαとβのアイソフォームが存在する.さらに,それぞれにsplicing variant であるTRα1,2,Δα1,Δα2とTRβ1,2,3,Δβ3が存在する.その構造は,核内受容体スーパーファミリーに相同性の高いC領域にzinc finger構造をもつDNA結合部位があり,E/F領域にはリガンドであるT3が結合するリガンド結合領域(LBD)がある.TRα1,TRβ1,2,3はT3のリガンド結合部位ならびに転写活性化能があるが,TRα2はC端側に構造変化があるためDNA結合能はあるがT3結合能はない.TRのアイソフォームは臓器ごとにそれらの発現が異なる.TRα1は心臓や脳に多く,甲状腺ホルモンのおもな心臓作用を担う.TRβ1はほとんどの臓器に発現するが,肝臓に多く甲状腺ホルモンによる脂質合成やコレステロール排出などの作用に関与する.一方TRβ2は下垂体や視床下部に発現し,後述する視床下部-下垂体-甲状腺フィードバック機構に関与する.
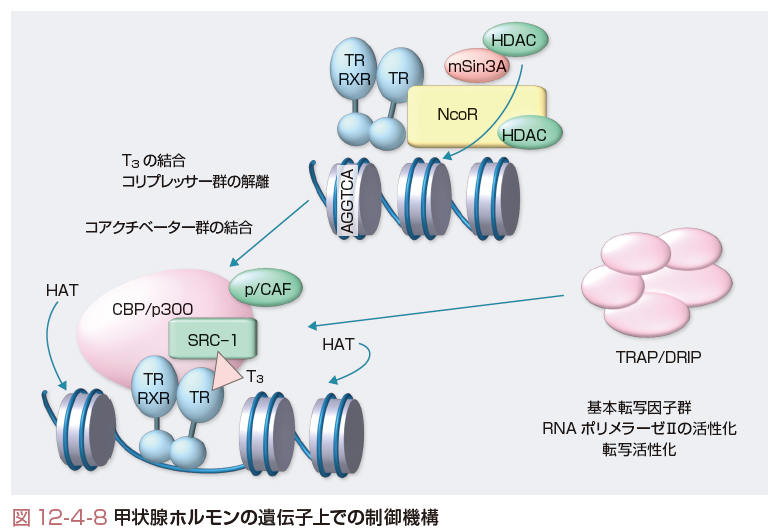
甲状腺ホルモン受容体は標的遺伝子上のプロモーター領域に存在する甲状腺ホルモン応答配列(TRE)に結合しており,T3がその受容体に結合することによりT3誘導性の転写因子として作用する.代表的なTREとして,AGGTCA(nnnn)AGGTCAという,任意の4塩基配列(n)を挟持したdirect repeat 4(DR4)がある.甲状腺ホルモン作用は標的遺伝子の違いにより,転写を活性化する系と転写を抑制する系がある.特にその活性系においては,分子機構が明らかとなりつつある.
現在想定されているモデルとしては図12-4-8に示すように,T3が結合していない状態でも甲状腺ホルモン受容体は標的遺伝子上に結合し,さらに転写活性抑制型の転写共役因子であるコリプレッサーのNCoRなどと結合している.このコリプレッサーは,ヒストンの脱アセチル化(HDAC)酵素と結合し,甲状腺ホルモン受容体が結合している周囲のヒストンを脱アセチル化し,クロマチンを凝集することにより転写活性を抑制している.T3がその受容体に結合すると,コリプレッサーが解離し,活性型の転写共役因子であるコアクチベーターのSRC-1やCBP,p/CAFなどが結合する.このコアクチベーターにはヒストンをアセチル化するヒストンアセチル化活性能(HAT)があり,ヒストンのアセチル化が起こり,クロマチンが弛緩状態となる.そこにTRAP/DRIPといったメディエーター群や基本転写因子群がRNAポリメラーゼⅡと結合し,転写が活性化することが提唱されている.
一方,甲状腺ホルモンによって転写が抑制される系には,後述する視床下部-下垂体-甲状腺フィードバック機構の主体をなす甲状腺ホルモンによるTRH遺伝子やTSH遺伝子を抑制する系がある.しかし,甲状腺ホルモン受容体が標的遺伝子上に直接結合することが必要か否かなども含めて詳細な分子機構はなお不明である.
(6)甲状腺ホルモン分泌調節機構
甲状腺における甲状腺ホルモンの合成分泌は自動性に乏しく,おもに視床下部-下垂体-甲状腺系によって制御されている.
a.甲状腺刺激ホルモン放出ホルモン(thyrotropin-releasing hormone:TRH)による下垂体TSHの調節
視床下部の室傍核で合成されたTRHは下垂体門脈系へと分泌され,下垂体前葉のTSH産生細胞膜に局在する膜7回貫通型のG蛋白質共役型受容体であるTRH受容体に結合し,TSHの産生ならびに分泌を刺激する.TRHはTSHα鎖,β鎖の重合や糖鎖の付加を刺激し,このTSHの糖鎖は生物学的活性に重要である.
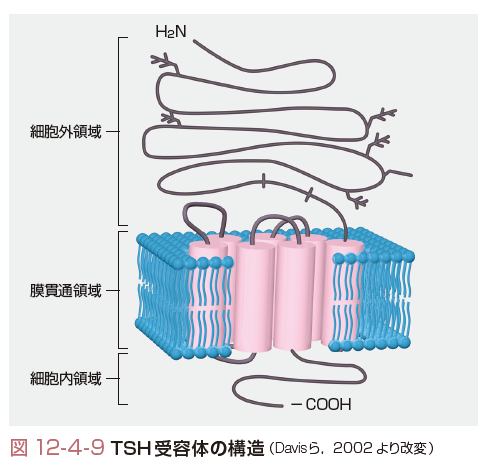
b.TSHによる甲状腺の調節
下垂体前葉から分泌されたTSHは甲状腺細胞膜に局在するTSH受容体に結合し,甲状腺ホルモンの合成,分泌を促進する.TSH受容体も膜7回貫通型のG蛋白質共役型受容体に属するが,LH,FSH受容体などと類似してN端側の細胞外領域が長い特徴を有する(図12-4-9).TSH刺激後,甲状腺細胞内では,cAMP,IP3が産生される.Basedow病の抗TSH受容体抗体は主としてこの細胞外領域のN端側に結合する.また,下垂体からはTSHとはまったく異なるα鎖,β鎖からなるサイロスティムリンが分泌されることが報告されているが,その生理学的作用は不明である.
c.甲状腺ホルモンによる視床下部TRHと下垂体TSHへの負の調節
分泌された甲状腺ホルモンは全身臓器に作用するが,それと同時に視床下部ならびに下垂体においてTRH,TSHの合成,分泌を抑制するネガティブフィードバック作用を有する.甲状腺ホルモン受容体の異常である甲状腺ホルモン不応症では,このフィードバック機構の障害のため血中甲状腺ホルモン値は高いにもかかわらず血清TSH値は抑制されない.
d.その他の甲状腺ホルモン分泌調節因子
前述したように大量のヨウ素は甲状腺機能に対して抑制的に作用する.また,妊娠時や胞状奇胎の際に高値を示すhCGは,TSHに比較すると作用は弱いが,それが大量に存在するとTSH受容体に作用して甲状腺機能亢進作用を示す.その他,ドパミンは下垂体のTSHの合成分泌を抑制し,寒冷刺激は中枢性にTRHを介してTSHの分泌を亢進する.[山田正信・森 昌朋]
■文献
Braverman LE, Utiger RD eds: The Thyroid, 9th ed, Lippincott-Raven Publishers, Philaderphia, 2004.
Davis TF, Larsen PR: Thyrotoxicosis. In: Williams Textbook of Endocrinology, 10th ed (Larser PR, Kronenberg HM, et al eds), p382, Saunders, Philadelphia, 2002.
Melmed S, Polonsky KS, et al: Williams Textbook of Endocrinology, 12th ed, Saunders, Philadelphia, 2011.

出典 内科学 第10版内科学 第10版について 情報
Sponserd by ![]()