
内科学 第10版 「下垂体後葉ホルモン」の解説
下垂体後葉ホルモン(下垂体後葉)
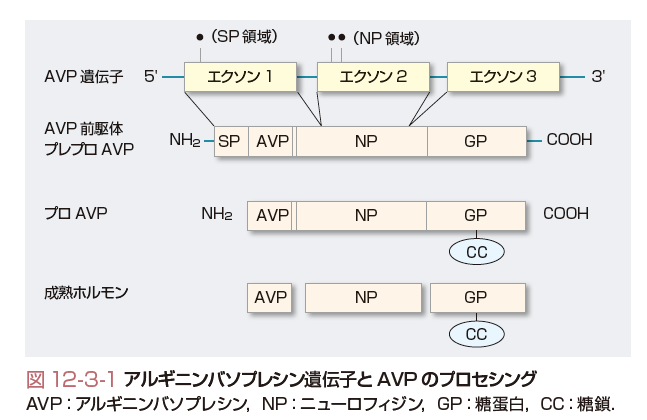
AVPは,9個のアミノ酸からなるペプチドホルモンである.AVP遺伝子は第20染色体にあり,3つのエクソンからなる(図12-3-1).エクソン1は,シグナルペプチド,AVP,ニューロフィジンのN端側をコードする.エクソン2は,ニューロフィジンの中央部をコードし,エクソン3はニューロフィジンのC端側と糖蛋白ドメインをコードしている.ホルモン前駆物質であるプレプロAVPは,シグナルペプチド,AVP,ニューロフィジン,糖蛋白を含む.プロAVPは,シグナルペプチドを除き,糖蛋白C端側にCOOHを加えた構造になっている.プロAVPは分泌顆粒内において下垂体茎内を移送される間に,AVP,ニューロフィジン,糖蛋白に分かれる.これらは下垂体後葉内に貯蔵されて,浸透圧や非浸透圧刺激に反応して血中へ放出される.ニューロフィジンや糖蛋白の機能はまだ明らかでないが,ニューロフィジンは小胞体におけるAVPのプロセシングに必須な蛋白であることは疑いない.事実,ニューロフィジン部分の塩基置換が生ずるとAVP合成障害を惹起して家族性中枢性尿崩症を発症することが知られている.
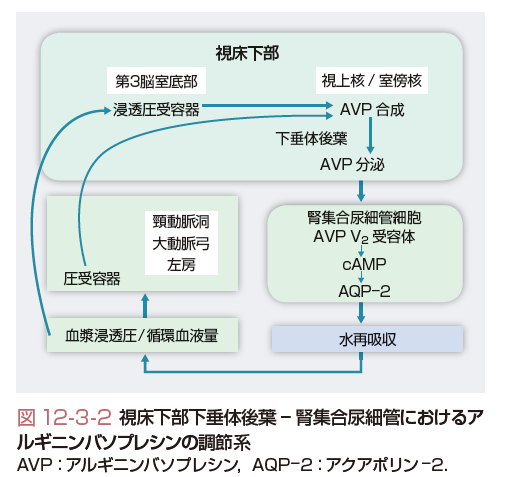
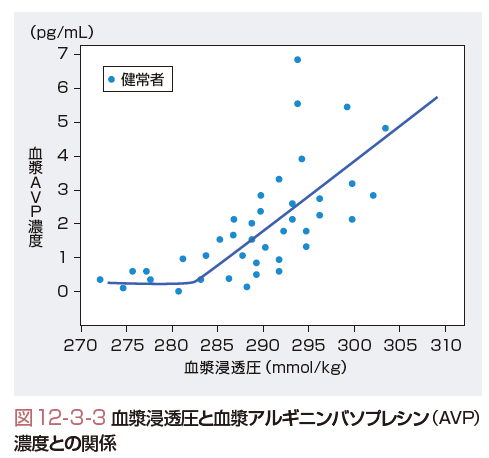
AVPの分泌調節は,浸透圧と非浸透圧刺激による.浸透圧受容器は前視床下部の第3脳室底部の終板に存在する.この部位は血液脳関門の外側になり,血漿浸透圧の変化を敏感に感受できる.浸透圧受容器から室傍核や視上核のAVP産生細胞へは神経経路が存在する(図12-3-2).この神経伝達はコリン作動性経路と考えられている.健常者では,血漿浸透圧と血漿AVP濃度の間には正の相関が認められる.図12-3-3に示すように,血漿浸透圧280 mmol/kgを臨界浸透圧として,血漿浸透圧の増加に伴って血漿AVP濃度は上昇する.一方,血漿浸透圧が280 mmol/kg以下ではAVPの分泌はみられない.血漿浸透圧の変化に対するAVPの分泌はきわめて鋭敏で,1 mmol/kgの増減に対応できる能力をもつ.
非浸透圧刺激は圧受容器で感受される.圧受容器は頸動脈洞,大動脈弓,左房にあり,その情報は迷走神経求心路を介して脳幹部延髄腹内側の孤束核に入力後,A1ニューロンを介して室傍核や視上核のAVP産生細胞に伝達される.この系は,恒常的にAVPの分泌を抑制的に制御する(tonic inhibition).非浸透圧性因子としては,循環血液量の減少,血圧の低下や左房圧の低下があげられる(図12-3-2).また,浸透圧受容器,圧受容器からAVP産生細胞への神経経路は互いに独立して入力するので,浸透圧,非浸透圧調節は各々独立した調節系を形成して,両者間に優位性は存在しない.
(2)バソプレシンの作用機構
AVPの作用はV1とV2作用に分けられる.V1作用は,AVP V1a,V1b受容体を介するもので,V1a受容体は血管平滑筋,腎メサンギウム細胞,肝細胞などに分布し,V1b受容体は下垂体前葉などに存在する.V2作用は,AVP V2受容体を介するもので腎集合尿細管での水の再吸収を引き起こす.AVP受容体蛋白は7カ所の膜貫通部をもつ典型的なG蛋白結合受容体である.V2受容体遺伝子はX染色体に位置し,アミノ酸は371個,分子量は40518である.V1a受容体のアミノ酸は418個で,分子量は46745である.
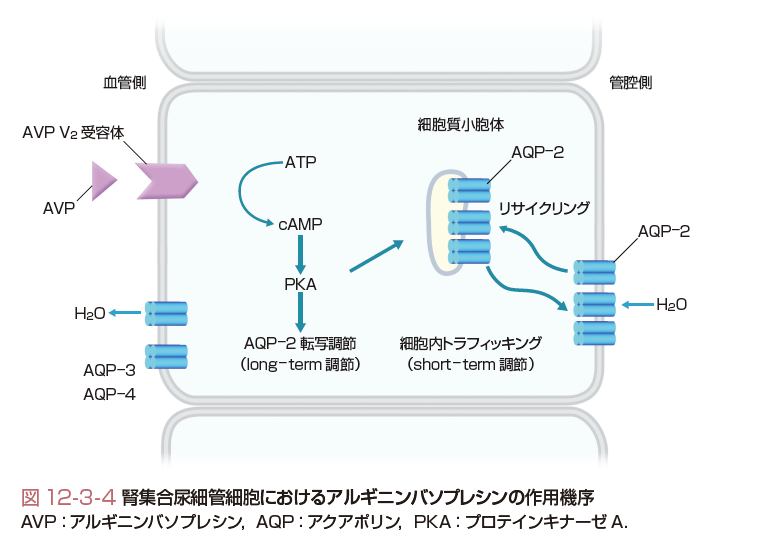
腎集合尿細管におけるAVPの作用は,主細胞の血管側細胞膜のV2受容体に結合後,グアニンヌクレオチド結合蛋白Gsを介して,触媒ユニットにおけるアデニル酸シクラーゼ活性を賦活する.産生された細胞内伝達物質cAMPは,プロテインキナーゼAを活性化後,アクアポリン-2(AQP-2)水チャネルを作動させる(図12-3-4).AVPによるアデニル酸シクラーゼの活性化は髄質内層集合尿細管に最も強く,ついで髄質外層,皮質部集合尿細管の順となる.AQP-2遺伝子は染色体12q13に位置し,アミノ酸は271個より構成される.AQP-2は膜6回貫通型の構造で2カ所にアスパラギン,プロリン,アラニンによるNPAボックスがあり,これが水分子の通過する孔形成に重要な構造となる.AVPによるAQP-2の調節はshort-termとlong-term調節に分けられる.short-term調節は,AVPによるAQP-2の細胞内トラフィッキングをいう.AQP-2が,AVPの刺激により管腔側細胞膜下の細胞質小胞体から管腔側細胞膜へ速やかに集簇されて,水の再吸収を引き起こす.また,AVPの作用がなくなると,AQP-2は再び管腔側細胞膜から細胞質小胞体へ移動する.集合尿細管細胞に吸収された水は血管側細胞膜上のAQP-3,AQP-4を介して血中に取り込まれる.long-term調節は,AVP依存性に集合尿細管細胞のAQP-2蛋白量を調節する.AVPは,AQP-2の5'上流域のプロモーターの転写活性を刺激して,AQP-2 mRNA,AQP-2蛋白合成を促進する.このように,AVPはAQP-2水チャネルの細胞内トラフィッキングと蛋白量の調節を行って腎における適切な水の再吸収を惹起する.
血管平滑筋細胞におけるAVPの作用は,V1a受容体を介する.AVPはV1a受容体に結合後,Gq蛋白を介してホスホリパーゼCを活性化する.ホスファチジルイノシトール4,5-二リン酸を加水分解して,イノシトール1,4,5-三リン酸(IP3)とジアシルグリセロール(DAG)を生成する.IP3はおもに小胞体よりCa2+を動員し,細胞外液からのCa2+流入の亢進とともに細胞内遊離Ca2+濃度を増加させる.DAGはCa2+とリン脂質の存在下にプロテインキナーゼCを賦活して,持続相の細胞収縮を維持させる.IP3により小胞体より動員された初期相の細胞内遊離Ca2+は,Ca2+結合蛋白カルモジュリンと結合した後,Ca2+-カルモジュリン依存性キナーゼを活性化する.ミオシン軽鎖キナーゼは,アクトミオシン複合体を形成して初期相の細胞収縮を引き起こす.DAGによるプロテインキナーゼCの活性化はNa+/H+交換系の活動を促進して,細胞内ナトリウム濃度の増加と細胞内pHのアルカリ化を惹起する.細胞内ナトリウム濃度の上昇は,Na+/Ca2+交換系に作用して,細胞内へのCa2+流入をさらに増加させて,細胞内pHのアルカリ化とともに持続相の細胞収縮を維持すると考えられる.また,AVPはアンジオテンシンⅡやエンドセリンなどと同様に血管平滑筋細胞や腎メサンジウム細胞の増殖も促す.このシグナル伝達系には蛋白質のチロシンリン酸化反応が関与する.プロテインキナーゼC以降,分子量42 kDaと44 kDaの蛋白質がリン酸化されて活性化されるmitogen-activated protein(MAP)キナーゼを介してその情報が最終的に核内へ伝達される.この伝達系の詳細はまだ明らかにされていない.
V1b受容体は,アミノ酸424個から構成され,下垂体前葉細胞に発現する.下垂体門脈系に放出されたAVPはV1b受容体を介して下垂体前葉からのACTH分泌を促進する.このほかV1b受容体は視床下部の室傍核や視上核,副腎髄質にも発現している.V1b受容体以降のシグナルは,V1a受容体以降と同様にGq蛋白を介してホスホリパーゼCの活性化から始まる.
(3)オキシトシンの分泌調節
オキシトシンは,視床下部で産生され,下垂体後葉から血中に分泌される神経内分泌ホルモンである.オキシトシンは,9個のアミノ酸よりなるペプチドホルモンで,おもな生理作用は子宮収縮と射乳である.乳頭刺激や,膣,子宮下部の開大はオキシトシン分泌を引き起こす求心性刺激となる.授乳時,児による乳頭の吸引刺激は脊髄の反射路を介して視床下部に投射され,下垂体後葉からのオキシトシン分泌を促進する.また分娩では児頭が子宮頸部と膣に侵入して拡張すると,その進展刺激によって脊髄路を介して視床下部に投射され,オキシトシンの分泌を惹起する.また,妊娠時ではオキシトシンの基礎分泌は妊娠週数とともに増加し,妊娠30週以降では非妊娠時の5~6倍に達する.
(4)オキシトシンの作用
オキシトシンは妊娠子宮に作用して平滑筋の収縮を引き起こして陣痛促進因子として分娩にかかわる.乳腺では,同様に平滑筋に作用して収縮を起こし乳頭の吸引刺激に反応した射乳に重要な役割をもつ. オキシトシンの作用はオキシトシン受容体を介して発現される.オキシトシン受容体は,膜7回貫通型のG蛋白結合受容体である.この細胞内シグナルはホスホリパーゼCを活性化後,AVP V1受容体以降のシグナル伝達と同様の経路で伝達される.[石川三衛]
出典 内科学 第10版内科学 第10版について 情報
Sponsored by ![]()