
内科学 第10版 の解説
心筋の収縮弛緩機構と心拍出量の調整(心血管代謝と機能)
心臓の機能はその役割から血液を駆出するポンプとしての機能,すなわち心拍出量によって一般的に評価される.とくに1心拍ずつの調節は急性の循環調節の理解にきわめて重要であり,その時点での,心筋自身の収縮能力(心収縮性)は,前負荷と後負荷によって調節されている.
(1)心筋細胞の構造と機能
心筋の収縮・弛緩機構を理解するには,基本となる心筋細胞の構造と機能を理解する必要がある.
成人心臓では心筋細胞は体積比で約75%,細胞数比で約30%を占めており,40~100 μmの長さで,10~20 μmの直径を有している.中央に核が存在し,筋原線維(myofibril)が長軸方向に並走して,ところどころ枝分かれし,細胞どうしは端々接合して,接合する細胞のあいだは介在板(intercalated disc)とよばれる特別の細胞膜よりなっている. 筋原線維は連続的に連なる心筋収縮の収縮構造単位であるサルコメアで構成されている.筋原線維は心筋細胞容積の約50%を占め,その間隙にはミトコンドリアが散在し,筋原線維を取り囲むように網状に筋小胞体(sarcoplasmic reticulum)が存在している.細胞膜にあたる筋鞘(sarcolemma)から細胞内に陥入しているT管は,Z線ごとに存在し,筋小胞体と接する部分でCa2+
トランジェントに重要な働きをしている.
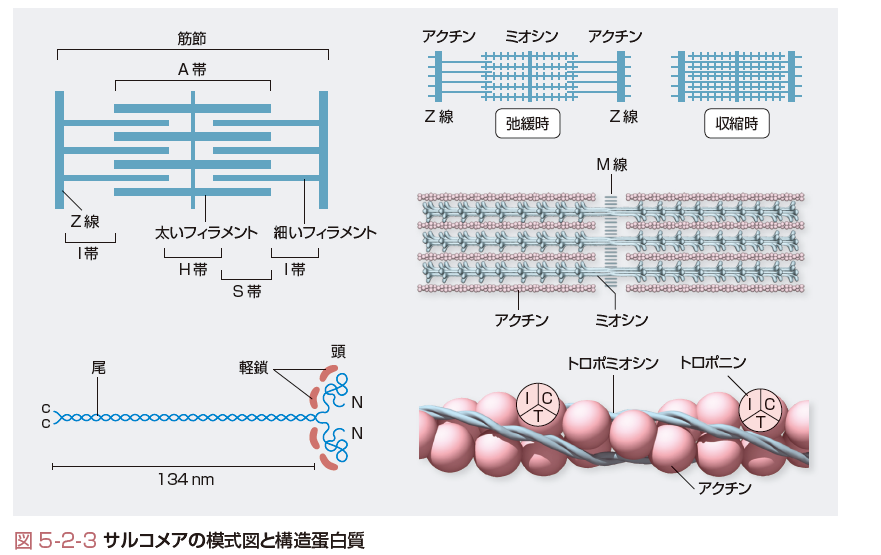
(2)サルコメアの構造と筋収縮(図5-2-3)
心筋内に張力が発生する機序はサルコメアの中に配列した収縮蛋白質が相互作用を行うことによる.サルコメアはZ線とZ線で区切られた間であり,約1.5 μmの長さの太いフィラメントと約1.0 μmの長さの細いフィラメントから構成されている.A帯は中央の肥厚部分M線に結合する太いフィラメントとI帯より延びた細いフィラメントの重なり合う部分であり,太いフィラメントのみの部分がH帯である. 太いフィラメントはミオシンより構成されており,ミオシン分子は2本の重鎖と4つの軽鎖よりなる.それぞれのフィラメントは規則正しく重合した300個のミオシンで構成されている.細いフィラメントはおもにアクチンより構成されており,Z線に端を発し,細いフィラメントのみのI帯,太いフィラメントとの重なり(S帯)となり,H帯の両端で終わる.アクチンフィラメントは球状の分子量47 kDaのアクチンが重合して線状となったものが2本で2重らせん構造を呈している.この2重らせん上の長い棒状の細いフィラメントにはトロポミオシンが結合しており,1つのトロポミオシンに7個のアクチン分子が連続的に一体となって結合し,7個目に1個のトロポニン複合体が結合している.トロポミオシンは3つのサブユニット,すなわち,トロポミオシン結合部(トロポニンT),Ca2+結合部(トロポニンC),アクチンと結合したATPase抑制部(トロポニンI)が存在する.
(3)興奮収縮連関
心筋細胞では,電気刺激により脱分極をきたし,これがトリガーとなってCa2+濃度の上昇をきたして収縮が起こる.これを興奮収縮連関という.
a.カルシウム・トランジェント
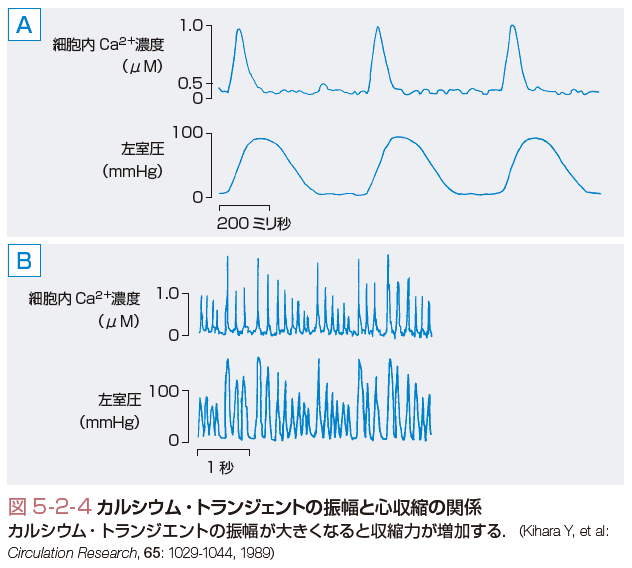
心筋細胞の収縮を起こす電気的興奮後のCa2+濃度の上昇は,速やかに低下し,収縮は弛緩へと移行する.このようなCa2+濃度の一過性変化をカルシウム・トランジェントという.カルシウム・トランジェントの振幅が大きくなれば発生する心筋の収縮力は増加し,小さくなれば減少する(図5-2-4).膜の脱分極に遅れてCa2+濃度は急激に増加し,その後,指数関数的に減衰する.張力はCa2+濃度の立ち上がりに遅れて発生し,張力がピークに達したときは,Ca2+濃度は静止時のレベル近くにまで減少している.カルシウム・トランジェントは,Ca2+の供給系,除去系,緩衝系の間の動的平衡により形成される.
b.カルシウム供給系
細胞内Ca2+は,筋細胞膜上のCaチャネルと筋小胞体膜上のCa2+チャネルから供給される.膜の脱分極に引き続いてT管上に存在する電位依存性L型Ca2+チャネル(LCC)を介して細胞外から細胞内にCa2+が流入する.このチャネルからのCa2+流入は少量であるが,筋小胞体からのリアノジン受容体を介するCa2+放出を引き起こすトリガーとして働く.この現象をCa2+誘発性Ca2+放出(calcium-induced calcium release)という.T管上のCaチャネルと筋小胞体上のリアノジン受容体は,互いが向き合うように接して存在している.リアノジン受容体からのCa2+の放出により,細胞内Ca2+濃度は10-7Mから10-5Mまで上昇し,Ca2+とトロポニンCのCa2+の結合が可能となり,サルコメアの収縮が開始される.calcium-induced calcium releaseは心筋に特異的で骨格筋には認められず,これが心筋収縮が細胞外Ca2+に依存する原因と考えられている.
c.カルシウム除去系
収縮したサルコメアが速やかに弛緩するためには,細胞内Ca2+濃度が速やかに低下する必要がある.脱分極後筋小胞体から放出されたCa2+はおもには再び筋小胞体にCa2+ポンプ(SERCA2a:sarcoplasmic reticulum Ca2+ ATPase)を介して汲み取られる.このCaポンプの活性は,Caポンプと結合しているホスホランバンにより調節される.ホスホランバンはCaポンプの取込能を抑制しているが,リン酸化されるとCaポンプの取込能を増加させる.カテコールアミン刺激はAキナーゼの活性化を介してホスホランバンをリン酸化し,Caポンプの機能を亢進させる.したがって,カテコールアミン刺激は,筋小胞体へのCa2+取り込みを亢進させ筋弛緩を促進するとともに,筋小胞体のCa2+貯蔵を増加させ,結果的にCa2+の放出を増加させる.
Ca2+を除去する主要な系として,筋小胞体のCaポンプ以外には,細胞膜上のナトリウム・カルシウム(Na/Ca)交換機構が働いている.この機構は膜電位などにより調節されているが,静止膜電位では,細胞外からのNa+流入と交換に細胞内Ca2+を汲み出す方向に働いている.
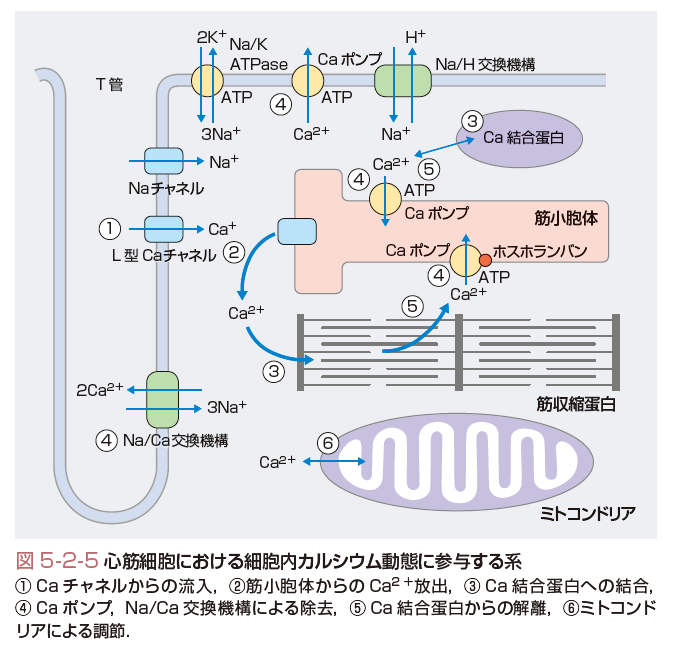
d.心筋細胞質内のカルシウム動態(図5-2-5)
以上,Ca2+トランジェントをまとめると以下のようになる.心筋細胞では,膜の脱分極によりCaチャネルからCa2+が流入し(①),流入したCa2+が筋小胞体からのCa2+放出を惹起し(②),細胞内Ca2+濃度が上昇する.Ca2+濃度の上昇に伴い,トロポニンなどのCa2+結合蛋白がCa2+と結合し(③),筋が収縮する.この間,筋小胞体や細胞膜のCaポンプが,Ca2+を細胞質から除去し続けており(④),徐々に細胞内Ca2+濃度は低下する.細胞内Ca2+の低下とともに,Ca結合蛋白から逆にCa2+が解離し(⑤),筋は弛緩する.さらに解離したCa2+は,筋小胞体上のCaポンプやNa/Ca交換機構により除去される.このとき,細胞膜Caチャネルより流入したCa2+とほぼ同量がNa/Ca交換により細胞外に汲み出され,筋小胞体から放出されたCa2+とほぼ同量が再度筋小胞体へ汲み上げられる.これにより,筋小胞体内のCa2+の量は一定に保たれる.さらに全過程を通じて,余剰のCa2+が生じればNa/Ca交換機構やミトコンドリアといったCaイオン緩衝系が調節する.
(4)サルコメアレベルでの収縮・弛緩機序
心筋の張力発生の分子的機構は,骨格筋と同様,アクチンとミオシンの相互作用による.アクチンとミオシンはフィラメント状になって,互いに重なり合うように平行に存在し,重なり合った部分で張力が発生する. 心筋細胞が興奮すると,心筋細胞内のCa2+濃度が上昇し,Ca2+がトロポニンCに結合し,トロポミオシンの構造変化をもたらし,その結果,トロポミオシンによって抑制されていたアクチンとミオシン頭部の会合が可能となり,ミオシン頭部の屈曲が生じ,筋収縮が生じる.ミオシン頭部にATPが結合することにより,ミオシン頭部とアクチンの会合がくずれ,ATPase活性でATPが分解され頭部はミオシン頭部が結合可能な状態に戻るが,ミオシン頭部は2~4個離れたアクチンに結合し,同じように筋収縮が繰り返される.この動作はCa2+濃度が高い間持続するが,拡張期に入りCa2+濃度が減少するとトロポニンCとCa2+の結合が解除され,筋は弛緩する. 収縮力の大きさは拡張期のサルコメア長に依存して大となる.これはin situでみれば心室拡張末期容積に依存してサルコメア長も長くなり駆出量,発生圧が大となるわけである(これは,従来よりFrank-Starlingの法則としてよく知られている).
(5)心機能と血行動態
a.心臓の収縮性
心臓の臓器レベルでの心収縮性の指標としては,左室圧の一次微分値dP/dt,左室の収縮末期の圧-容積関係から求めるEmaxなどがある.
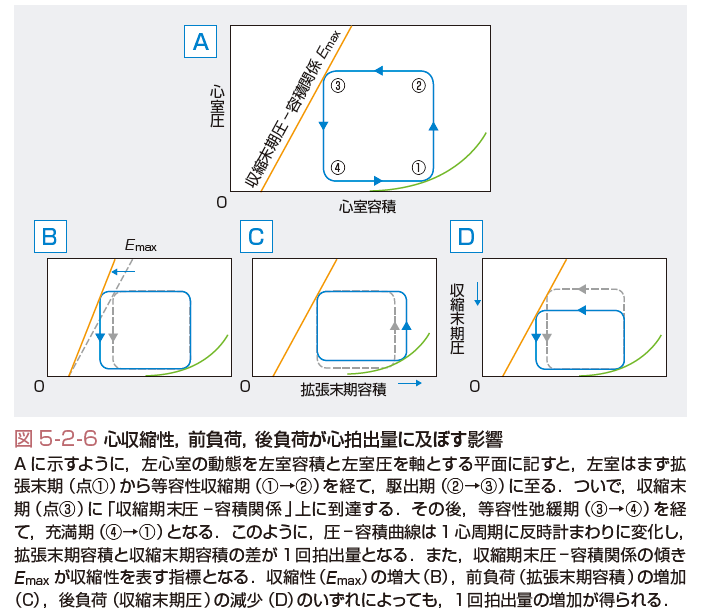
心室の圧-容積関係は,心室を可変弾性体と見なし,その内圧と容積に着目している(図5-2-6).心室の圧/容積比は,収縮開始後,時間の経過とともに上昇し,収縮期末の時点で最大となる.この値がEmaxであり,収縮性の指標となる.Emaxは,下大静脈バルーン閉塞法などにより前負荷を変化させ,その間の収縮期末での圧・容積を数点プロットし,回帰直線の傾きとして求められる.
b.前負荷の影響
前負荷とは,心筋が収縮する直前の拡張末期に心室筋に加えられている負荷をいう.臨床的には,拡張末期容積,または拡張末期圧をもって当てることが多い.また,前負荷の指標として,肺動脈圧,肺動脈楔入圧,中心静脈圧が用いられる.静脈還流量は拡張末期容積の変化に大きく関与している.
1回の心拍出では,直前の拡張期の心室充満がその心拍にとっての前負荷となる.1心拍の前負荷が少ない例として期外収縮があげられる.心室が十分に充満されず,前負荷が少ないため,1回拍出量が減少する.上室性頻拍などの頻脈でも,拡張期時間の短縮により心室充満(前負荷)が不十分となり,1回拍出量が減少する.逆に,代償性期外収縮の次の心拍では,代償性に心室充満時間が延び前負荷が増加するため,1回拍出量が増加する(post-extrasystolic potentiation).心房細動では心拍間隔が一定しないため,心室充満の程度も一定せず,心拍出量も毎回異なる.
心室充満には,拡張期時間のみならず,心房収縮による心室への血液の送り込みも重要である.房室解離では心室への血液の送り込み効率が低下し,心拍出にとって不利になる.心室ペーシング時の血行動態は房室解離に類似し十分な心拍出量が得られない場合があるので,心房心室順次ペーシング(生理的ペーシング)が望まれる.
c.後負荷の影響
後負荷とは,収縮期において,心室から大動脈に血液を拍出する際に心室筋に加わる負荷をいう.臨床的には,左室圧,最大収縮期血圧,平均動脈圧,末梢血管抵抗,左室壁応力などによって評価される. 後負荷の指標として全末梢血管抵抗が用いられるが,これは心臓からの血液の拍出や血流が定常状態にある,すなわち,直流と考えており,拍動流の近似にすぎない.末梢血管の拍動流に対する抵抗をより正確に表現するには,心拍出量や血流の心周期変化も考慮し,大動脈インピーダンスなど,交流に基づく考えを導入する必要がある. 大動脈弁狭窄症,動脈硬化症などの疾患では後負荷が増大し,敗血症性ショックやアナフィラキシーショックでは後負荷が過小になる.カテコールアミンは後負荷を増加させ,Ca拮抗薬や亜硝酸剤などの血管拡張薬,あるいは入浴は後負荷を減少させる.また,大動脈内バルーン・ポンピングも後負荷を減少させる効果を有する.
d.心拍数の影響
心拍数に変化のない場合,1回拍出量と心拍出量は同様に変化する. (心拍出量)=(1回拍出量)×(心拍数)
一方,心拍数が増加すると拡張期は短くなり,拡張期流入血液量が減少する.これは前負荷の減少をもたらし,1回拍出量を減少させるが,当初は心拍数の増加でもって代償し,心拍出量は増加する.しかし,心拍数が極端に増加すると,1回拍出量の低下が著明になり,心拍出量を維持することができなくなる.
e.Starlingの心臓法則と心機能曲線
Starlingは,1回心拍出量は,後負荷が一定のとき,左室への流入量(静脈圧,もしくは,左室拡張末期容積)によって決まると結論づけた.これをStarlingの心臓法則とよぶ.
心筋では,筋節の長さが長いほど発生張力は大きい(Starling効果).心室への流入量が増加すれば左室拡張末期容積は増大し,心筋線維が引き延ばされて発生張力は増加し,その結果,後負荷が同じならばより多くの駆出量が得られる.
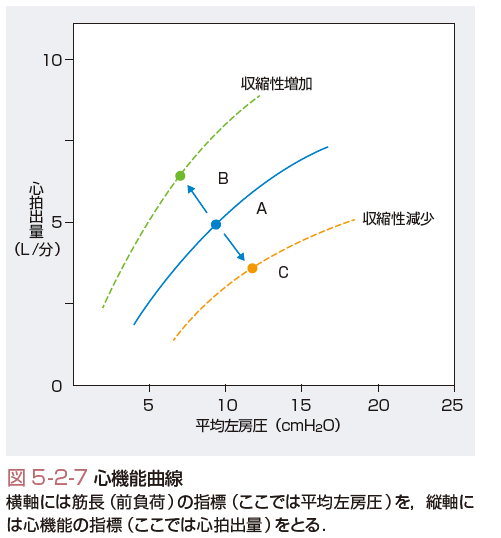
心機能曲線は,横軸に左室拡張末期容積などの前負荷指標をとり,縦軸に心機能の指標をとって描いた曲線である(図5-2-7).心機能曲線が左下から右上に向かうのは,Starlingの心臓法則を示している.心収縮性が亢進すると曲線は左上方に移動して,心収縮性が低下すると右下方に移動する.心機能曲線のシフトから心機能を判定することが可能になる.また,後負荷の減少は心機能曲線を左上方へ移動させ,増大は右下方へ移動させる.心機能曲線では心収縮性の変化と後負荷の変化が区別できないので,注意が必要である.
(6)強心薬の作用機序
強心薬は心筋に作用して収縮性を増強させる薬物であるが,その作用機序は一定でない. 交感神経β受容体刺激薬は交感神経β受容体を刺激して細胞内サイクリックAMP(cAMP)濃度を増し,cAMPの分解酵素であるホスホジエステラーゼを阻害することにより細胞内cAMP濃度を増す.cAMPの増加はAキナーゼを活性化し,電位依存性L型Ca2+チャンネルをリン酸化し,開口率を高めCa2+の流入を増加させる結果,筋小胞体からのCa2+放出を促進し強い心筋収縮力を生じる.一方,Aキナーゼの活性化はホスホランバンをリン酸化させ,その結果筋小胞体上のSERCA2aの抑制が解除され筋小胞体への細胞質から筋小胞体へのCa2+の汲み上げが促進され心筋の弛緩速度を高める.その他,トロポニンIもAキナーゼによってリン酸化され,トロポニンCとCa2+の親和性を低下させその結果アクチンとミオシンの架橋形成が抑制され心筋の弛緩が促進される.このように,カテコールアミンでは心筋の収縮・弛緩ともに亢進する. ピモベンダンなどのCa感受性増強薬は,収縮機構のCa2+に対する感受性を増加させることにより,Caイオン濃度の増加を介さずに収縮性の増加をもたらす.しかし,カルシウム感受性増強薬は,原理的には収縮期の張力のみならず,拡張期の張力も増加させることになるので,拡張機能を低下させる可能性がある.[斎藤能彦]
■文献
Ganong WF: Ganong’s Review of Medical Physiology, 2th ed, Mc Graw, Hill, 2012.
Opie LH: The Heart: Physiology, from Cell to Circulation, 3rd ed, Lippincott-Raven, Philadelphia,
1998.吉川純一,他編:心機能を知る(心臓病診療プラクティス3).文光堂,東京,1994.
出典 内科学 第10版内科学 第10版について 情報
Sponserd by ![]()