
改訂新版 世界大百科事典 「筋収縮」の意味・わかりやすい解説
筋収縮 (きんしゅうしゅく)
muscle contraction
筋肉が神経のインパルスあるいは刺激によって収縮する現象をいう。筋肉は筋繊維(あるいは筋細胞ともいう)からなり,筋収縮は個々の筋繊維の収縮にもとづく。一般に筋繊維の収縮は細胞膜内外の電位差(膜電位)の変化によって調節されている。膜電位の減少を脱分極といい,脱分極がある値をこえると収縮がおこる。
筋収縮の神経による調節
筋肉は,身体の運動をおこす骨格筋,心臓の拍動をおこす心筋,消化管や血管の運動にあずかる平滑筋に大別される。骨格筋の収縮は,脊髄内の運動ニューロンからのびた運動神経によって調節されている。運動神経繊維のインパルスが神経筋接合部に達すると,神経末端から伝達物質(アセチルコリン)が分泌され,神経筋接合部の筋繊維の膜に作用して活動電位を発生させる。活動電位は,速やかに筋繊維の全長にわたって伝播し,筋繊維全体の収縮をひきおこす。単一の活動電位によっておこる短い収縮を単収縮という。活動電位は〈全か無かの法則〉(ある一定以上の強さの刺激が与えられたとき初めて反応がおこり,それ以下ではまったく反応がみられず,また逆にそれ以上はいかに強い刺激を与えても反応は変わらない,という法則)にしたがうので,単収縮も全か無かの法則にしたがう。活動電位が短期間にくり返し発生すると,単収縮は加重して大きな持続的収縮すなわち強縮がおこる。われわれの身体の運動はほとんどすべてが骨格筋の強縮によるものである。心筋の活動電位は,骨格筋よりもはるかに持続時間が長く,また絶対不応期も長い。このため心筋の収縮は加重がおこらず,全か無の法則にしたがって拍動する。平滑筋の収縮は交感神経と副交感神経によって調節されており,一方が収縮を他方が弛緩をおこすが,そのしくみには不明な点が多い。
脱分極と収縮との関係
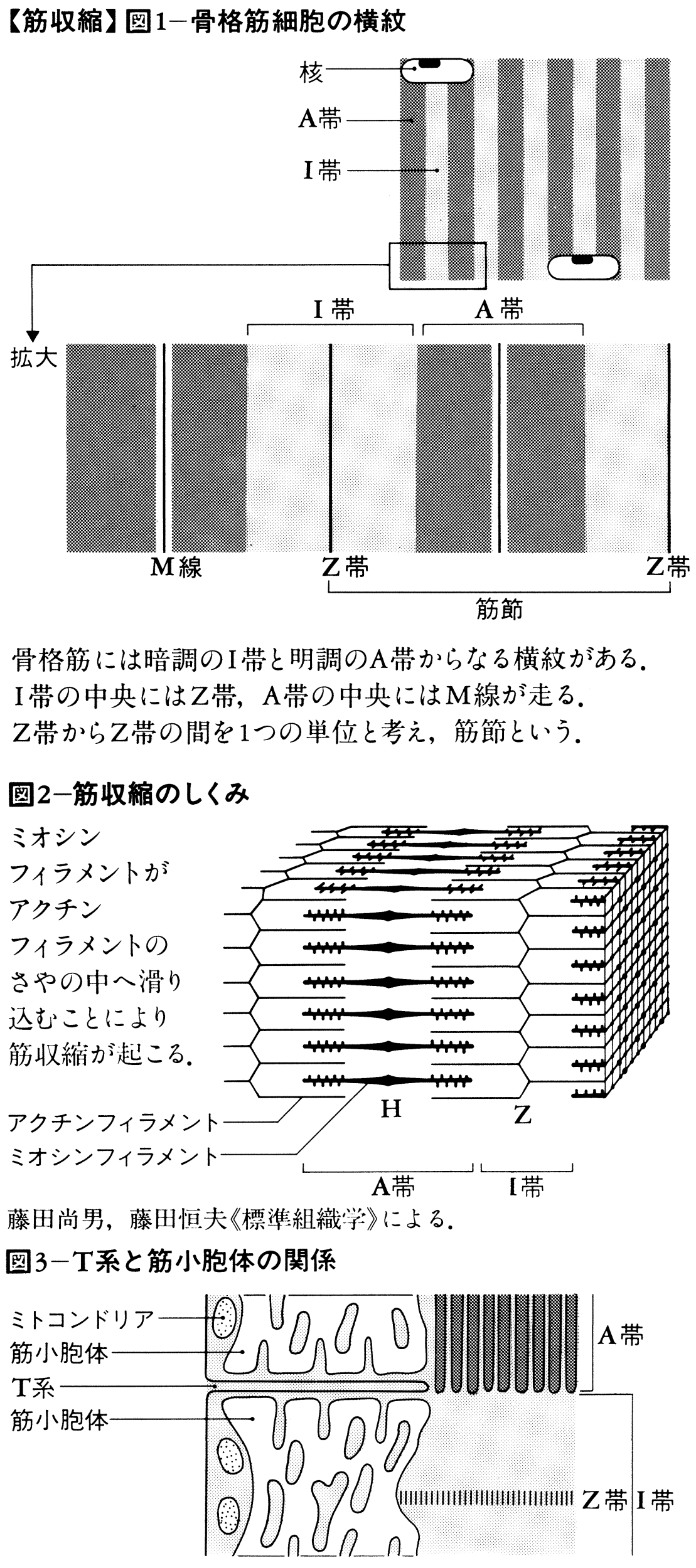
筋収縮は,筋フィラメントを形成するタンパク質アクチンとミオシン間の反応によるものであり,このエネルギー源はATPである。静止状態の筋肉では,アクチンとミオシン間の反応がトロポミオシンおよびトロポニンというタンパク質によって抑制されているので収縮はおこらない。この抑制は微量(10⁻6~10⁻5mol)のカルシウムイオンCa2⁺によってとり除かれ,アクチンとミオシン間の反応が開始されて収縮がおこる。つまり筋収縮は,筋フィラメントのまわりの微量のCa2⁺の増減によって調節される。骨格筋繊維の内部には筋小胞体sarcoplasmic reticulumという袋状の膜構造が発達しており,その内腔にCa2⁺をとり込む性質がある。このため静止状態の筋繊維内のCa2⁺は大部分筋小胞体内腔にとり込まれており,筋フィラメント周囲のCa2⁺濃度は10⁻7mol以下にすぎない。一方,筋繊維の細胞膜は多くの部位で管状に内部におちくぼみをつくっており,これを横行小管系(T系)transverse systemという。横行小管は筋小胞体と多くの部位で密接しているので,筋繊維細胞膜におこる活動電位あるいは脱分極の影響は筋小胞体にただちに伝えられ,その結果,筋小胞体内腔のCa2⁺が筋細胞質中に放出される。筋フィラメント周囲のCa2⁺が10⁻6molをこえると収縮がおこり10⁻5molで収縮は最大となる。活動電位がやむと筋小胞体からのCa2⁺放出も止まり,筋フィラメント周囲のCa2⁺はふたたび筋小胞体中にとり込まれて筋肉は弛緩する。筋小胞体のCa2⁺とり込みはATPのエネルギーを用いるポンプ作用(能動輸送)であるが,Ca2⁺放出のしくみはまだ明らかでない。心筋や平滑筋では収縮・弛緩の調節にあずかるCa2⁺は,筋小胞体から放出されるほか,筋肉周囲の外液からも流入すると考えられている。
筋収縮のしくみ,滑り説
筋収縮のしくみについてはこれまで多くの説が提出されてきた。筋肉の微細構造がまだ不明な時期に考えられた説は,すべて鎖状高分子の折りたたみによって収縮がおこるとしているので,折りたたみ説と総称される。1954年にH.E.ハクスリーらが骨格筋繊維が2種の筋フィラメントからなること,筋収縮の際,筋フィラメントの長さは変わらずたがいに滑り合うことを電子顕微鏡で示してからは折りたたみ説は消失し,筋フィラメント間の滑りをおこすしくみによって筋収縮を説明しようとする滑り説が登場した。2種の筋フィラメントのうち,細いフィラメントは主としてアクチンからなり,太いフィラメントはミオシンからなる。アクチンフィラメントは球状のアクチン分子が二重ラセンをなして重合したものである。ミオシン分子はふくらんだ2個の頭部と棒状の尾部をもつ。尾部は束になって太いフィラメントの軸を形成し,頭部は側方に突き出て側枝となっている。筋収縮のさい2種の筋フィラメントは能動的に筋繊維の長さを短くする方向にたがいに滑り込んでいく。ハクスリーによれば,この滑りをおこす原動力は,太いフィラメントの側枝がまず細いフィラメントのアクチンと結合し,ついで結合したままボートのオールのように運動して筋フィラメントを滑らせたのちアクチンから解離するという反応サイクルをくり返すことによるという。しかし太いフィラメント側枝の運動は現在にいたるまで確かめられていない。最近,筋フィラメント間の滑りはリニアモーターカーのような静電気的なしくみで行われるという説もあらわれている。
筋収縮の性質
筋収縮の研究法は,筋肉の両端を固定して張力の発生を記録する等尺性収縮と筋肉に一定の荷重をかけて短縮を記録する等張力性収縮の二つに大別される。骨格筋の等尺性収縮張力は,筋肉が生体長(関節を可動範囲の中間位置にしたときの筋肉の長さ)のとき最大となり,筋肉の長さがこれより長くなっても短くなっても減少する。ヒトの骨格筋の最大張力は5~6kg/cm2であり,ヒトの身体のすべての筋肉の発生する張力の総和は約20tにも達する。一方,筋肉の等張力性収縮の際の短縮速度は,荷重が小さいほど大きく,荷重ゼロのとき最大となる。骨格筋の最大短縮速度を比較するには,筋繊維の構造的・機能的単位である筋節の半分あたりの短縮速度を用いる。この値は,筋フィラメントの滑りの速度に対応しており,20℃で約10μm/sである。また平滑筋の等尺性収縮張力は骨格筋よりも大きく,二枚貝の貝柱の平滑筋では10kg/cm2以上である。平滑筋の短縮速度は一般に骨格筋よりもはるかに遅い。
特殊な筋収縮
昆虫のうちハチ,ブヨ,ハエなどの翅は非常な高頻度(毎秒200~1000回)ではばたく。しかし,高頻度ではばたいている昆虫の翅の筋肉からは低頻度の活動電位(毎秒数回)しか記録されない。これは翅の筋肉と昆虫の外骨格とが高頻度の共振をおこして翅をはばたかせているためであり,低頻度の活動電位はこの共振状態の維持に必要である。したがって個々のはばたきは単収縮によるものではなく,昆虫ははばたきの頻度をみずから調節することはできない。セミの発音器官の筋肉も鼓膜と共振をおこす。なお,他のすべての昆虫では,翅の筋肉の活動電位とはばたきの頻度とは等しいので,個々のはばたきは翅の筋肉の単収縮によるものである。以上の例とは逆に,二枚貝類の殻の開閉筋(貝柱の筋肉)は活動電位の発生なしに長時間等尺性張力発生をつづける。このとき筋肉の代謝速度は静止状態とほとんど同じである。このような持続的収縮中の筋肉は等張性短縮を行うことができず,この状態から静止状態にもどるには弛緩神経のインパルスが必要である。
→筋肉
執筆者:杉 晴夫
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponsored by ![]()