
最新 心理学事典 「記憶の進化」の解説
きおくのしんか
記憶の進化
evolution of memory system
【動物の記憶研究】 動物の記憶研究は,シカゴ大学のハンターHunter,W.S.によって開始された。彼は博士学位論文(1913)の中で,ラット,イヌ,アライグマ,および2.5~8歳のヒトの幼児を対象に,遅延反応課題delayed reaction taskという方法を用いて,情報の保持可能時間を比較している。被験体を出発箱に閉じ込め,三つのうちの1部屋を点灯して,遅延時間ののち点灯していた部屋に入ることを訓練し,遅延を徐々に延長して限界を求めた。最大値は,ラットで10秒,イヌで5分,アライグマで25秒,子どもでは25分であった。しかしラットとイヌおよび1頭のアライグマでは,遅延終了時の体や頭部の向きが選択の最大の決定因で,遅延時間中正しい向きを維持できれば正解とした。しかし他のアライグマや子どもの反応は,そうした身体的手がかりに依存しないもので,何か個体内部にある手がかりを使っていたとしか考えられないという。このことからハンターは,この手法が,動物における表象的要因representative factorの研究として適切なものであるとした。
【動物の記憶研究の手法】 ハンターの研究は,記憶過程を非言語的に研究できることを示したものとしてきわめて重要である。しかし,空間的表象以外の記憶を扱えないなどの難点があるため,その後の研究では,種々の他の手続きが利用されるようになった。
1.遅延見本合わせdelayed matching to sample 1930年代に霊長類の知的行動の研究が盛んになると,概念や抽象能力などの分析のための高次の知的課題として,見本合わせmatching to sample課題がしばしば用いられるようになる(Weinstein,B.,1941ほか)。この課題に遅延を組み合わせたものが遅延見本合わせで,この課題では,見本刺激の提示後,見本が消失し,遅延時間をおいて二つ以上の刺激が提示される。その中から,見本と同じ刺激を選択することが正解である。ハーローHarlow,H.F.ら(1943)は,両側の前頭前皮質を切除したアカゲザルを対象に,遅延見本合わせを行なった。同じ場面を用いて行なった空間的遅延反応課題と比較すると,最大遅延可能時間こそ短いものの,サルは5~10秒遅延の見本合わせが可能であったと報告されている。
ブラウBlough,D.S.(1959)は,スキナー箱を使って,ハトの遅延見本合わせを分析した。ハトの正答率は遅延5秒で偶然レベル近くまで低下した。しかし1羽のハトは,遅延期間中,見本刺激の種別に対応する分化した行動を示すようになり,この反応をしたときには,遅延が10秒でも正答率はほとんど低下しなかった。この行動は,符号化を伴う一種の行動的リハーサルといえる。
遅延見本合わせは,記憶に影響する種々の要因を独立に操作しやすく,さまざまな変形課題を含め,今日の動物の記憶研究でよく利用される。たとえば,見本の呈示時間や見本に触れる反応の要求回数を変えたりして,記銘の容易さを操作することができる。赤ならば縦棒,青ならば横棒を選ぶというような象徴見本合わせsymbolic matching to sample課題にすれば,記憶がどのように符号化されて保持されているかを調べることもできる。
2.放射状迷路radial arm maze オルトンOlton,D.S.ら(1976)は,装置の中央の部屋から,放射状に多数のアームが伸び,各アームの入口には開閉可能なドアがある,放射状迷路とよばれる空間記憶を検討する新装置を考案した。この装置のすべてのアームの先端に報酬を置き,ラットを中央に入れてドアを開放すると,ラットはアームの先端に行って報酬を食べ,中央に戻って,別のアームに行くことを繰り返す。彼らの研究では,最初の8選択のうち,未訪問のアームを選択した平均本数は7.4本であった。つまりラットは,ほとんど同じアームを訪れることなく,きわめて効率よく採食する。17アームの迷路では,平均15本であったという(Olton,et al.,1977)。
放射状迷路課題の遂行は,短期記憶に基づくものではなく,きわめて保持可能時間の長い記憶に基づくもののようである。たとえば,8本中4本を選択した時点でいったんラットを装置から取り出し,遅延後に再び迷路に戻すと,4時間後までは遅延の効果はなく,24時間後にようやく成績が偶然レベルまで低下した(Beatty,W.W.,& Shavalia,D.A.,1980)。
同じ考え方に基づく手法として,空間内に複数設けられた採食場所を次々に訪問させる課題がある。種々の霊長類(MacDonald,S.E.,& Wilkie,D.M.,1990ほか)や鳥類(Kamil,A.C.,Balda,R.P.,& Olson,D.J.,1994ほか)が,放射状迷路同様に容易に学習することが知られている。ハトでは,五つのキーを使った類似課題でその学習が確認されている(Zentall,T.R.et al.,1990)。イヌはラットと同様に放射状迷路遂行を学習するが,ラットの成績には及ばない(Macpherson,K.,& Roberts,W.A.,2010)。
3.水迷路water maze モリスMorris,R.G.M.(1981)は,円形のタンクに白濁した水を入れ,ラットを泳がせて,見えない浅瀬にたどり着くことを学習させた。その後,浅瀬を取り去ってテストすると,ラットは,もともと浅瀬のあった4分円部分に圧倒的に長く滞在した。直接的に目標を示す手がかりはなくとも,水槽内外にあるなんらかの手がかりを使って,長期的に目標位置を学んだのである。モリス水迷路Morris water mazeともよばれるこの装置は,マウスやラットのナビゲーションにおける空間情報の利用を調べるために,しばしば用いられる。毎日同じ場所に浅瀬を設置すれば長期記憶,毎日異なる場所に設置すれば短期記憶を調べることができる。行動薬理学や行動遺伝学,神経科学などでは,特定の薬物や遺伝子や脳部位などが記憶能力に及ぼす影響を調べるための標準的な方法として,広く用いられている(D'Hooge,R.,& De Deyn,P.P.,2001)。
【記憶機能の比較研究】 ヒトとヒト以外の動物の記憶系は相同関係にあるのだろうか。これを考察するために,ヒトの記憶機能に見られる種々の特徴が動物で検討されている。
1.干渉interference 類似した記憶課題を続けて行なうと,単独で同じ記憶課題を行なった場合より成績が低下する。先行課題が後続課題の遂行を妨害する現象は順向干渉proactive interference(順向抑制proactive inhibition),その逆は逆向干渉retroactive interference(逆向抑制retroactive inhibition)とよばれる。グラントGrant,D.S.(1975)は,ハトに色の見本合わせを訓練すると,直前の試行と当該試行で見本刺激の色が異なるとき正答率が低くなることを示した。これは前試行からの順向干渉によると考えられる。実森正子らは,花の写真と霊長類の写真を用いて系列項目再認課題(後述)をアカゲザルに訓練した。セッションの前半は一方の写真,後半は他方だけを提示した。サルの成績は同じタイプの写真が続くにつれて低下し,切り替えとともに突然回復した(Jitsumori,et al.,1989)。これは順向干渉とそれからの回復を示すものと思われる。
逆向干渉は,たとえばハトでは,色の遅延見本合わせの遅延時間中に色刺激を挿入すると,図形を挿入するよりも正答率が大きく低下する(Zentall,1973)。他方,視覚刺激の遅延見本合わせの遅延時間に聴覚刺激を提示しても,フサオマキザルの成績は大きな影響を受けない(Worsham,R.W.,& D'Amato,M.R.,1973)。つまり動物においても逆向干渉が生じ,それは記憶素材に類似したもので強くなると思われる。しかし最近の研究では,感覚様相の異なる刺激でも,逆向干渉が見られる場合があることも示されている。たとえば足立幾磨らは,アカゲザルに,既知のサル個体の動画をその個体の写真に合わせる遅延見本合わせを訓練した。習得後,写真を出す前に動画に一致したあるいは一致しない個体の音声を流してテストすると,不一致の場合には成績が低下した(Adachi,I.,& Hampton,R.R.,2011)。同様の結果はリスザルのヒト弁別課題でも見られている(Adachi & Fujita,K.,2007)。これらは,聴覚刺激が視覚像を想起させ,それが視覚的記憶に干渉したものと解釈できる。
2.系列位置効果serial position effect ライトWright,A.A.ら(1980)は,系列項目再認serial probe recognition課題をアカゲザルに訓練した。2ヵ所の刺激呈示場所の上段に,種々の写真を10~20枚次々に呈示した。最終項目呈示後,下段に1枚の写真を呈示した。サルはこの写真がリスト中にあったかどうかを,レバーを左右に動かして答えた。系列位置による正答率の変化は,同じ課題を行なったヒトとサルで酷似していた。また彼らはこの課題を4項目に簡略化し,最終項目呈示後の遅延時間の操作による系列位置効果の変化の様相が,ハト,アカゲザル,フサオマキザル,ヒトでほぼ同じであることを示している(Wright,1999;Wright,et al.,1985)。系列位置効果は多くの種に見られる現象であり,そのメカニズムも共通しているのではないかと推測される。聴覚刺激でも確認されているが,視覚と聴覚では,異なった性質が示されるという報告もある(Wright,1998)。
3.符号化coding ヒトは与えられた情報をさまざまな形に変換して利用する。こうした情報の変換を符号化とよんでいる。情報を元の形で記憶し,必要な時に必要な情報に変換するやり方を回顧的符号化retrospective coding,あらかじめ将来必要な情報に変換しておくやり方を予見的符号化prospective codingとよぶ。ヒトはしばしば後者の戦術を取る。ギャファンGaffan,D.(1977)は,アカゲザルに色をキーの位置に符号化する課題を与えた。サルの誤反応は,見本が類似した赤と琥珀ではなく,正解キーの位置が近接した青と赤の間で多かった。つまりサルはこの事態で色から位置への予見的符号化をしたことが示唆された。同様の行動はハトでも見られる(Roitblat,H.L.,1980)。またラットでは,放射状迷路遂行において,既訪問アーム数が多くなると,回顧的符号化から予見的符号化に方略を切り替えることも示唆されている(Cook,R.G.et al.,1985)。
4.指示された忘却directed forgetting ヒトは不要な情報をあえて記憶せず,認知的負荷を減らす場合がある。マキMaki,W.S.ら(1980)は,食物の有無を,色キーをつつき分けて答える象徴見本合わせをハトに訓練した。このとき,数秒の遅延の間の室内灯の点灯-消灯によって,色キーをつつく場面があるかそのまま試行が終わるかが決まっていた。つまり室内灯の状態によっては,見本を忘れてもよかった。ハトの正答率は,「忘れろ」手がかりの後思いがけず色キーをつつかせると,甚だしく低下した。ハトは忘れてよい時には憶えていようとしなかったのだと思われる。指示された忘却はラットでもサルでも示されている(Grant,D.S.,1982; Roberts,W.A.et al.,1984ほか)。
5.記憶再生memory recall 動物を用いた記憶再生の研究は希少である。メディンMedin,D.L.(1969)は,4×4の小扉の行列のうち,1,2,4個を点灯したパターンを見本刺激として1秒間呈示し,0~20秒の遅延時間ののちに,点灯した扉すべてを開けることをアカゲザルに求めた。正解の扉を開けて報酬を取るとそこが点灯した。サルは順次扉を開けて,見本と同じパターンを作ることを学習した。マッケイMackay,H.A.ら(1971)は,アカゲザルに,横に並んだ8個のキーのうち2個が順次点灯するパターンを見本として呈示した。点灯したキーをサルが押すとライトは消えた。直後サルは,同じ反応系列の再生が求められた。サルはこれを学習した。
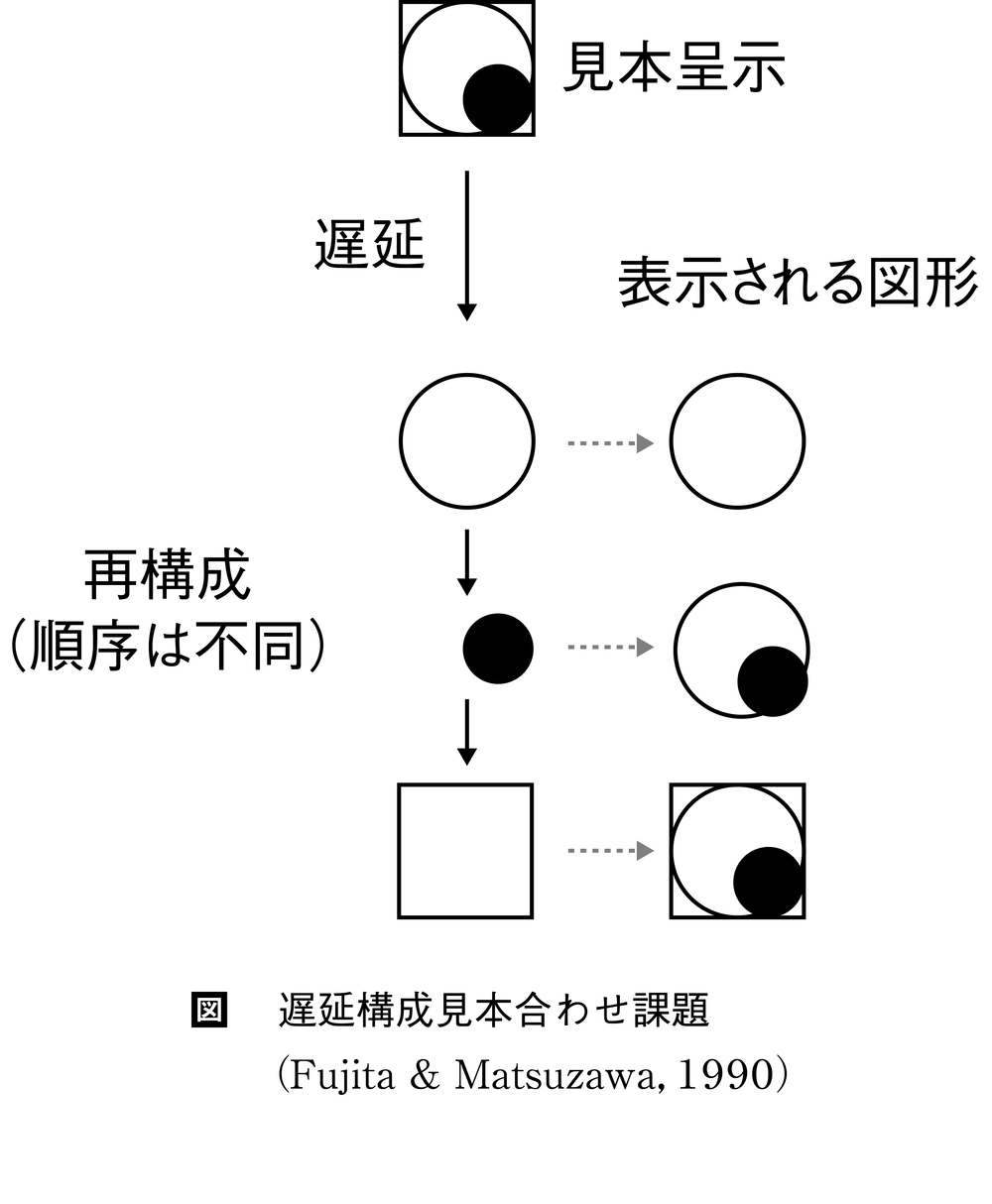
上述の2研究は,位置パターンの再構成と見ることができるが,刺激パターン自体が手がかりであったか否かには疑問が残る。藤田和生らは,1頭のチンパンジーに図形の再構成を訓練した。○,□などの9種類の要素図形を三つ重ね合わせてできる84種の図形を1秒間だけ見本として呈示し,遅延後,要素図形のキーを選んで図形を重ね合わせ,再構成させた(図)。最大遅延(32秒)におけるチンパンジーの正再構成率は約80%と高く,同じ課題を行なったヒト成人の成績に遜色がなかった(Fujita & Matsuzawa,T.,1990)。
ハンプトンHampton,R.R.ら(2011)は,アカゲザルに,5×5の行列上に置かれた赤青2刺激の配置を見本として呈示し,遅延時間後,青だけを別の場所に呈示して,マークされた2ヵ所のうちどちらかに触れて,見本パターンと同じ配置を再構成する課題を訓練した。サルはこれを習得し,その学習は,3刺激の配置を含む新奇パターンに般化した。同じ2刺激パターンの再認課題に比べると成績は遅延時間によらず低く,ヒトと同傾向を示した。これはヒトとサルの記憶系の機能の共通起源を示すものではないかと著者らは述べている。
6.短期記憶容量short-term memory span ヒトの短期記憶容量は7±2項目であるといわれている。動物ではどうだろうか。川合伸幸らは,1頭のチンパンジーに,画面に複数呈示されるアラビア数字を小さいものから順に触れていくよう求めた。正答率は5項目の場合に65%であった。つまりチンパンジーは,少なくとも5個までは情報を保持できると考えられる。これは就学前児と同等である(Kawai & Matsuzawa,2000)。井上紗奈らは,5歳のチンパンジーの子どもは,9個の数字でも高い正答率を示すことを報告している。しかし,数字の提示時間を210ミリ秒ときわめて短くしても成績は低下しないことなどから,直観像のようなものの可能性も示唆している(Inoue & Matsuzawa,2007)。
7.意味記憶容量semantic memory span 分散貯蔵scattered hoarding(あるいは貯食food caching)する鳥類には,ハイイロホシガラスのように秋に数万個の種子を地面の穴に隠し,翌夏まで利用するものもいる。しかしこうした適応的特殊化と思われる行動に関与する長期記憶が,ヒトのそれと同じものと考えて良いのかは判断が難しい。ファゴFagot,J.ら(2006)は,ギニアヒヒとハトに,既知と未知のカラー写真を左右のキーに振り分けることを求め,これを3~5年続けた。枚数が増えるとともに,全体の正答率はほぼ直線的に低下していったが,2頭のヒヒは約6000枚の写真を約80%の正答率で分類した。2羽のハトは2000~3000枚を約60~70%で正しく分類した。いずれの種も,驚くほど多数の写真と反応場所の関係を記憶し保持できることが示された。この研究では,呈示された刺激セットの順序に対して,長期的リスト学習に見られるヒト同様の系列位置効果も生じた。ヒヒとハトで質的な違いはなく,これらは意味記憶の相同性を示すものと思われる。ファゴらは,鳥類の祖先である爬虫類と哺乳類の分岐後,記憶容量は変化したが,記憶過程の基本部分や作動様式は変化しなかったのではないかと論じている。
8.メタ記憶meta-memory メタ記憶とよばれる自身の記憶に関する認知は,動物にもあるのだろうか。ハンプトン(2001)は,アカゲザルに遅延見本合わせを訓練し,遅延時間後,比較刺激を呈示する直前に,課題選択場面を挿入した。課題選択場面では,遅延見本合わせを行なうか,回避して簡単なキー押し課題に行くかを選択できた。強制的に見本合わせに行く試行もあった。見本合わせに正解すると価値の高い報酬がもらえた。キー押し課題では低価値の報酬が必ずもらえた。こうすると,見本合わせを強制された試行より,それを自ら選択した試行の方が,サルの正答率は高くなった。見本合わせの回避率は,遅延が長い場合や見本を出さない場合には高くなった。これらから,アカゲザルは自身の記憶痕跡の強さをモニターできること,すなわちメタ記憶をもつことが示唆された。同様の課題でフサオマキザルはメタ記憶を示すが(Fujita,2009),鳥類では否定的である(ハト:Inman,A.,& Shettleworth,S.J.,1999; Sutton,J.E.,& Shettleworth,2008,ハシブトガラス:Goto,K.,& Watanabe,S.,2011)。
9.エピソード記憶episodic memory エピソード記憶とは個人的体験の記憶で,いつ,どこで,何が,という情報が統合されたものであること,偶発的事象の内省的想起として,事後に必要に応じて意識的に取り出されることの二つの主要な特徴をもつ。what-where-whenからwww記憶ともよばれる前者の性質は,鳥類,げっ歯類および類人猿で例証されている。
クレイトンClayton,N.S.ら(1998)は,貯食行動を使ってフロリダカケスのwww記憶を検討した。まず2種類の餌の保存性を教えた。カケスに,砂入りの皿にハチミツガの幼虫とピーナッツを隠させ,ピーナッツは124時間後でも食べられるが幼虫は腐って食べられないことを数度経験させた。次に,2連の皿の片側に一方の餌を隠させたあと,120時間後に反対側に他方の餌を隠させ,その4時間後に取り出させた。カケスは,幼虫を後で隠した場合には,まずそちらを探った。しかし幼虫を先に隠した場合には,逆にピーナッツを隠したほうの皿を探った。つまりカケスは,いつ,どこに,何を隠したかをすべて手がかりとして利用した反応をした。
マルティン・オルダスMartin-Ordas,G.ら(2010)は,ボノボとチンパンジーとオランウータンで,よく似た簡便なwww課題を試している。トレイの3穴の一つに時間が経つと溶けて流れて消える大好物の氷菓,もう一つにブドウを置いた。不透明のカップをかぶせて,5分または1時間後にカップを選ばせた。その結果,全体として類人猿は,1時間後よりも5分後に,有意に多く氷菓の方を選択した。しかし,選択が逆転するまでには至らず,個体差も大きいので,カケスほど明瞭な結果ではない。
バッブBabb,S.J.ら(2005)は,8アームの放射状迷路でラットのwww記憶を検討し,同様にラットも,いつ,どこに,何が得られたかをすべて手がかりとして利用している可能性をもつことを報告している。全体としてこれらの結果は,比較的広範な動物種が,www記憶をもちうることを示している。
10.心的時間旅行mental time travel エピソードの想起は,過去の事象を現在の心的表象内に再現することだと考えることができる。エピソード記憶のこの働きは,近年では心的時間旅行とよばれ,このシステムを未来に向けて機能させ,将来の計画を立てることがエピソード記憶の適応的意義だと考える研究者もいる(Tulving,E.,2005ほか)。
ラビーRaby,C.R.ら(2007)は,アメリカカケスが過去経験を将来計画に生かす能力をもつことを示す実験を行なったが,カケスの行動には適応的特殊化の要素が含まれており,ヒトの将来計画と相同なものかについては議論が分かれる。学習性の行動を用いて同様の事実が示されればその問題はない。ムルカイMulcahy,N.J.ら(2006)は,ボノボとオランウータンでこれを検討した。筒に細い棒を差し込んで食物を落下させる道具使用を教えたのち,実験室にいろいろな棒を並べ,類人猿を導き入れた。道具課題は準備されていたが,穴が透明の板で塞がれていた。5分後,類人猿を隣の待合室に出し,残った棒を片付けて透明の板を取り外し,1時間後に類人猿を実験室に入れた。これを繰り返すと,類人猿は自発的に適切な道具を待合室から持ち出し,実験室に持ち込んでくるようになった。ボノボとオランウータン各1頭について,遅延時間を14時間に延長し,12試行テストしたところ,オランウータンは第2試行以降全試行で成功した。ボノボも第2試行以降8試行で成功した。こうした行動は,特定の道具を持ち帰り持ち込んだときに実験者が報酬を与える手続きでは,ほとんど生じなかった。つまり類人猿たちは,将来道具を使うために,準備的行動として,適切な道具を持ち帰ったのだと思われる。
将来を見越して現在の行動を調節する能力は,ヒトに限られたものではなく,複数の動物種に備わった能力なのかもしれない。動物たちの記憶の諸過程は,ヒトのそれに質的にはよく似ており,相同な起源を見て取ることができる。 →記憶 →動物実験法 →符号化 →見本合わせ
〔藤田 和生〕
出典 最新 心理学事典最新 心理学事典について 情報
Sponsored by ![]()