
最新 心理学事典 「視覚領野」の解説
しかくりょうや
視覚領野
visual area
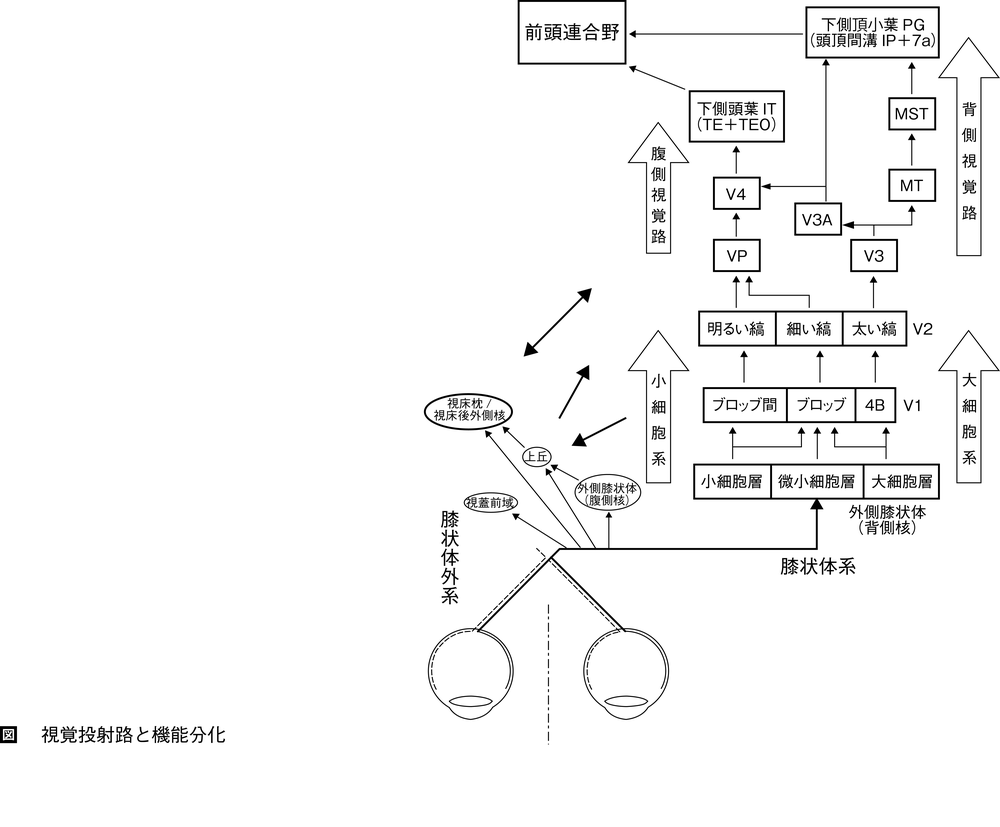
【視覚系の概要】 視覚情報の処理は,網膜の視細胞において可視光(波長400~700nmの電磁波)が視物質に吸収され,細胞膜を介するイオン流の変化に変換されることに始まる。網膜神経節細胞retinal ganglion cellに生じた活動電位は,視床の外側膝状体背側核dorsal lateral geniculate nucleus(以下,外側膝状体lateral geniculate nucleusとしてLGNと略記する)を経て,大脳皮質の一次視覚野に伝わる。一次視覚野では,視野の構造や物体の図形特徴,奥行きや動きなどの情報を抽出し,後頭連合野の視覚前野(後述)に出力する。大脳皮質における視覚情報処理は,2系の並列投射路による。すなわち,物体の位置や動きなど運動・空間視に関する情報は,視覚前野から背側視覚路を経て頭頂連合野へ,形や色など形態視情報は視覚前野から腹側視覚路を経て側頭連合野(下側頭葉)へと伝えられる。この2系の視覚情報は前頭連合野prefrontal association areaで統合されて,視野空間の構成,そこに存する物体,自己との関係性などについての認識が成立する(図)。
【網膜retina】 光は眼球の角膜および水晶体で屈折し,眼底の網膜上に焦点を結ぶ。網膜は厚さ0.3~0.4mmの神経組織で,投影された光の分布や強度,波長の情報を活動電位に変換して出力する。網膜には5種類の神経細胞(ニューロン),すなわち視細胞photoreceptor,水平細胞horizontal cell,双極細胞bipolar cell,アマクリン細胞amacrine cell,神経節細胞ganglion cellがある。
視細胞は桿体rodと錐体coneに区別される。視細胞において視物質visual pigmentが光を吸収すると,一連の化学反応を経て,視細胞膜の電位応答が生じ,それが信号として双極細胞以後の網膜内神経回路に伝わる。桿体視物質は吸光感度のピークを波長約500nmとする1種類のみであるが,錐体視物質は560nm(赤),530nm(緑),420nm(青)に最大吸光感度を示す3種類があり,それぞれの視物質をもつ錐体を赤錐体,緑錐体,青錐体とよぶ。桿体は錐体に比べて光に対する感度が1000倍も高く,暗所視scotopic visionを担う。桿体は網膜で中心視に対応する中心窩fovea を避けて分布する。これに対して,錐体は明所視photopic vision条件で働き,3種類の錐体が波長の異なる光を選択的に吸収し,色覚の情報を伝える。中心窩には錐体のみが高密度に分布し,空間分解能が最も高い。錐体の電気信号は,錐体→錐体双極細胞→神経節細胞と送られ,神経節細胞に生じた活動電位が視神経からLGNへと出力される。桿体の信号は,桿体→桿体双極細胞→アマクリン細胞→錐体双極細胞→神経節細胞という経路で出力される。水平細胞およびアマクリン細胞は,周辺受容野,桿体経路,神経節細胞の運動方向選択性など受容野特性の形成にかかわる。
視覚系の各神経細胞は網膜上の一定範囲の光情報を処理し,その範囲を受容野receptive fieldとよぶが,より一般的には網膜上の受容野に対応する視野空間内の位置と範囲として定義される。網膜の出力細胞である神経節細胞の受容野は中心部と周辺部からなる同心円状であり,たとえば中心部に光が当たったとき(オン),周辺部の光が消えたとき(オフ)というように,中心部と周辺部が逆の明るさ変化に反応する。これを中心-周辺拮抗型受容野antagonistic center-surround receptive fieldという。このような受容野は明るさのコントラスト,すなわち物体の輪郭や境界の検出に利点がある。
霊長類の神経節細胞は,大型のM神経節細胞M ganglion cell(約10%),小型のP神経節細胞P ganglion cell(約80%),K神経節細胞K ganglion cell(約10%)に分類される。M細胞は受容野が大きく,コントラスト感度が最も高く,粗い形状検出,また物体の運動やそのタイミングを検出するが,色選択性を示さない。これに対してP細胞は受容野が小さく,高い空間解像力で詳細な形の情報を検出する。またP細胞とK細胞の受容野は色選択性を示し,P型は赤-緑,K型は青-黄の中心-周辺拮抗型である。これらの細胞の活動により色を含む形態視情報がLGNに送られる。M型神経節細胞はLGNの大細胞層,P型は小細胞層,K型は各層の間(層間層)の細胞に入力する。このLGNを経て一次視覚野に向かう経路を膝状体系geniculate systemという。これに対し,M細胞およびK細胞の一部は,視線制御や眼球運動にかかわる上丘superior colliculusにも投射し,さらに視床枕pulvinar,視床後外側核posterior lateral thalamic nucleusを経て視覚前野と連絡する経路があり,膝状体外系extrageniculate systemという。
【外側膝状体lateral geniculate nucleus(LGN)】 網膜神経節細胞の軸索は視神経optic nerveとなって視神経円盤optic diskより眼球を出た後,脳底部で視交叉optic chiasmを形成する。視交叉以後の視神経束は視索optic tractとよばれるが,耳側網膜由来の視神経は交叉せず同側へ,鼻側網膜由来の視神経は交叉して対側のLGNへと向かう。これにより右LGNには左半視野,左LGNには右半視野の情報が入力する。
LGNは6層構造で,腹側の第1,2層は大細胞層magnocellular layer,背側の第3~6層は小細胞層parvocellular layerとよばれる。これらの層の間にも小型のニューロン群が存在し,微小細胞層koniocellular layer(層間層intercalated layer)とよばれる。大細胞層にはM,小細胞層にはP,微小細胞層にはK神経節細胞に由来する神経線維が入力する。また各層は単眼性入力を受ける。LGN細胞の受容野は中心-周辺拮抗同心円型であり,各層は入力神経節細胞の特性を反映し,機能的な並列情報処理経路の構造的基礎となる。大細胞層ニューロンは,コントラスト感度が高く,明るさの変化に対して一過性の応答を示し,時間分解能が高い。これは物体の出現や動きの検出に必要な性質である。また空間分解能は低く,色選択性を示さない。大細胞層ニューロンは,背側視覚路dorsal pathway(運動視・空間視経路)のもとになる大細胞系magnocellular systemを構成する。これに対して小細胞層ニューロンは,空間分解能が高く,赤や緑など色に選択的で持続的な応答を示すが,大細胞系に比べて時間分解能,およびコントラスト感度は低い。小細胞層ニューロンは,形態視に必要な情報を処理する腹側視覚路ventral pathway(形態視経路)のもとになる小細胞系parvocellular systemを構成する。微小細胞層ニューロンに始まる微小細胞系koniocellular systemは形態視経路に合流する。
LGN内のシナプスのうち神経節細胞由来のものは約10%にすぎず,大脳皮質由来が約30%,脳幹由来が約30%,残りは抑制性シナプスである。大脳皮質の活動に依存してLGNの出力がコントロールされていると考えられる。また,脳幹からのアセチルコリンなど脳の活動性を全般的に制御する投射系により,覚醒レベルや注意を反映して大脳皮質へのゲート機能を担うことも,並列投射路の形成と並んで,LGNの重要な役割である。
【大脳皮質視覚野visual cortex】 大脳皮質視覚野は,後頭葉から側頭葉にかけて,V1すなわちvisual area 1(一次視覚野),V2(二次視覚野),V3(三次視覚野),V4(四次視覚野),MT(中側頭野middle temporal)あるいはV5(五次視覚野),MST(中上側頭野medial superior temporal)などとよばれる機能分化した多くの領野で構成されている。
【一次視覚野primary visual cortex】
1.位置 一次視覚野(17野area 17または V1)は後頭葉のブロードマン17野に位置する。マカクザルにおいては月状溝と下後頭溝にはさまれた位置で,後頭葉皮質表面に広く露出する。ヒトでは大脳半球内側面の鳥距溝周囲である。右半球のV1では左視野,左半球のV1では右視野の情報が処理されている。
網膜像は視野の2次元投影であるが,この連続表現はV1においても脳表面上の視野地図visual field mapとして保持されており,これを網膜部位再現retinotopic representation,retinotopyという。V1では視野の中心部が周辺視野に比べて拡大的に表現・処理されている(皮質拡大cortical magnification)。
2.層分化と機能分化 V1灰白質は6層構造で,厚さは約2mmである。V1は,視床からの入力を受ける小型の顆粒細胞が多い第Ⅳ層が発達している。第Ⅳ層はさらに4A,4B,4Cα,4Cβの4亜層に分かれ,LGNからの大細胞系および小細胞系の求心性線維は第Ⅵ層で一部分枝するが,その後,大細胞系は4Cα層に,小細胞系は4Cβ層に入力する。微小細胞系は直接に第Ⅱ・Ⅲ層に入力する。4B層には皮質表面に対して平行な有髄神経線維のバンド(ジェナリGennariの線条)が明瞭であるため,V1は有線野(線条野)ともよばれる。第Ⅳ層以外には皮質表面に向かって伸びる太い尖端樹状突起apical dendriteをもつ錐体細胞pyramidal cellが多い。脳表面から見ると,第Ⅱ・Ⅲ層にはチトクローム酸化酵素染色によって濃染される直径約200μm程度のブロッブblobとよばれる構造が斑点状に多数分布する。ブロッブは皮質の垂直断面で見ると全層を貫く柱状構造であり,色に選択的な応答を示すニューロンが多い。
各層は機能的に分化しており,4C層より浅い第Ⅱ・Ⅲ層,4B層は大細胞系,小細胞系,微小細胞系の情報を統合再編し,視覚前野に出力する。V1から視覚前野への投射は主にV2に向かうが,V2にはチトクローム酸化酵素染色により太い縞thick stripe,細い縞thin stripe,明るい縞pale stripeとよばれる三つのストライプ構造が見られ,機能的にも分化している。すなわち,V1の4Cα層からは主に4B層,一部ブロッブに投射し,4B層からはV2の太い縞,V3,MT野に出力され,頭頂連合野に向かう背側視覚路を形成する。4Cβ層からは第Ⅱ・Ⅲ層ブロッブおよびブロッブ間に投射し,V2の細い縞および明るい縞に出力され,以後V4を経て下側頭葉に向かう腹側視覚路を形成する。V1の第Ⅴ・Ⅵ層は浅層から入力を受け,それぞれ皮質下の上丘およびLGNに投射する。上丘への投射は,眼球運動の制御,LGNへの投射はV1への出力のフィードバック制御にかかわる。
3.受容野および特徴抽出性 LGNニューロンは円形受容野をもち,刺激の傾きに対して弱い選択性しか示さない。しかし,多数の求心性線維が収束するV1ニューロンでは受容野サイズが大きく,また特定の軸方向に伸びた形であり,その軸の傾きに沿った線分に選択的に応答する。これを方位選択性orientation selectivityという。求心性線維からの入力を受け,V1内の階層的情報処理の初段階にある単純型細胞simple cellは,光の照射に反応する領域(オン野)と消滅に反応する領域(オフ野)が隣り合う受容野をもち,この受容野の長軸の傾きがニューロンの最適刺激方位と一致する。単純型細胞は星状細胞stellate cell(顆粒細胞granule cell)であることが多い。複数の単純型細胞からの入力が収束する複雑型細胞complex cellは主に錐体細胞であり,その受容野はオン野とオフ野が重なり合い,光刺激の明暗の位相に依存しない反応を示す。
V1ニューロンは視覚刺激の方位,運動方向,大きさ,空間周波数,時間周波数,色(波長),両眼視差,明るさのコントラストなどさまざまの図形的特徴に選択的な反応を示し,これを刺激選択性stimulus selectivityあるいは刺激特異性stimulus specificityという。V1ニューロンは受容野に呈示された視覚刺激の特徴に応じて反応するが,受容野周囲の広い範囲に存在する刺激の特徴にも依存して主に抑制性の影響を受ける。これを文脈依存的反応修飾contextual modulationまたは受容野周囲抑制surround suppressionという。この性質は,複雑な視野を分節化し,知覚的な図と地の分離figure-ground segregationを成立させるために重要である。
4.機能コラム V1では,皮質表面方向から見て近傍同士のニューロンは類似の反応特性を示し,層間を縦方向に連絡する神経投射によって,浅層から深層までを貫く機能的柱状構造(機能コラムfunctional column)とよばれる特定の刺激特徴の処理単位とみなされる構造ができる。ヒューベルHubel,D.H.とウィーゼルWiesel,T.N.は,サルのV1でどちらの眼に由来する入力で活動するかという眼優位性コラムocular dominance columnと共通の刺激方位に選択的に応答するニューロンの方位コラムorientation columnについて構造的モデル(1977)を提案した。サルのV1では一つの眼優位性コラムの幅が約0.4mm,左右一対で約1mmである。また各眼優位性コラムが延びている方向に沿って50μm離れた場所では,最適方位が約10°ずれることから約1mmで180°の全方位をカバーすることになる。ヒューベルとウィーゼルは,この皮質上の約1mm四方の範囲に,特定の視野範囲についてのすべての情報を処理するコラムが統合されているとし,これをハイパーコラム(超円柱)hyper columnとよんだ。
現在ではV1の機能コラムとして,眼優位性コラム,方位コラム,ピンウィールpinwheel(方位コラムの風車状の配列),ブロッブ(色コラム),空間周波数コラムspatial frequency columnなどが知られる。これらの各種機能コラムは視野地図と重畳し,全視野の刺激特徴を組み合わせ処理するマクロな神経回路の構造的基礎となっている。
【視覚前野prestriate area】 後頭葉連合野occipital association cortexともいう。マカクザル大脳皮質の視覚関連領野は30以上に細分化している。後頭葉の最後部にあるV1の浅層(第Ⅱ・Ⅲ層および4B層)からは,視覚前野に出力する。
視覚前野は,もともとブロードマンによって,細胞構築学的に18野・19野とされている。しかし近年の研究は,視覚皮質がさらに多数の領野に細分化していることを明らかにし,後頭葉の最後部に広がるV1に対する位置関係や,脳葉あるいは主要な脳溝・脳回における位置などで,これらの領野を区別する命名が行なわれるようになった。この結果,18野は主としてV2,V3,V3A,19野はV4,MT(V5)から成る。V2はV1の前に隣接し,サルでは月状溝および下後頭溝の後壁に細長く広がる。V2のチトクローム酸化酵素染色による太い縞には背側路,細い縞と明るい縞には腹側路へと向かう情報が伝えられる。V2の太い縞および明るい縞領域には,明暗や色の境界による線が存在しないにもかかわらず端点や角,弧などの配置によって遮蔽物体を知覚する主観的輪郭subjective contour(または幻の輪郭illusory contour)に反応するニューロンが多数観察される。V3は,ニューロンの多くが方向選択性direction selectivityや両眼視差選択性binocular disparity selectivityを示すのに対して,色選択性color selectivityを示すものは少ないことから,運動・空間視情報を分析・統合してV3A,MTに送る背側路に含まれると考えられる。ただしV3Aは腹側路にも連絡すると見られる。これに対して,腹側V2の前方に帯状に広がるVP(ventral posterior)とよばれる領野は,方向選択性細胞が少なく,色選択性細胞が大半を占めることからV4とともに形態視経路(腹側路)に含まれると考えられる。
MT(中側頭野middle temporal,すなわちV5)は上側頭溝superior temporal sulcus後壁の小さな領野であり,V1の4B層,V2の太い縞,V3から主たる入力を受ける。MTには運動方向選択性を示すニューロンが集まって,同じ方向に選択性を示すニューロンがコラム状に配列している。MTニューロンの方向選択的な活動は,物体の運動方向の知覚と相関し,MTの刺激や部分的破壊は,動きの判断やその閾値を変化させる。MTは大細胞系入力に依存しており,等輝度色から成る運動刺激に反応しない。MTからは頭頂連合野のMST(中上側頭野medial superior temporal)とよばれる領野に情報が送られ,MSTでは広視野の動きと物体との相対運動,図形の拡大・縮小,等距離平面上での物体の接近・離反,追跡性眼球運動関連活動など,自身と視野の相対的な動きとその関係性についての処理を行なう。
V4は背側部でV3,V3Aと接し,腹側部でVPと接し,前方では下側頭葉inferior temporal cortex(ITすなわちinferior temporal,inferotemporal)後部に接する。V4に主たる入力を与えているのはV2の細い縞および明るい縞であり,出力先はITである。V4は均質な領域ではなく色選択的な応答を示す場所と,色選択性のない形態特徴に選択性を示すニューロンや渦巻き状図形,放射状図形など極図形・双極幾何図形など複雑なパターンに反応する場所がモザイク状に分布している。V4の色処理は,特定の物体とその周囲から反射されてくる光のスペクトルを比較して色の恒常性color constancyのもとになる性質を作り出すと考えられている。V4の損傷は主観的輪郭図形を含む図形弁別の障害や,色弁別,色の恒常性知覚の障害を生じる。
【背側視覚路と腹側視覚路】 V1からは,MT野medial temporal areaに直接投射する経路と,V1からV2,V3を経てMT野に至る経路がある。これらは頭頂連合野parietal association areaに向かい,MSTを経て頭頂間溝intraparietal sulcus(IPS)に至り,背側視覚路dorsal visual pathwayとよばれる。この経路は運動・空間視に関する大細胞系の性質を反映しており,頭頂間溝において細分化された領野の活動は,視空間の構造の認識,自身と視空間の定位,視覚情報の運動制御情報への変換,立体視,物体の動きの認識,多種感覚情報の統合など,実空間で動きまわり,働きかけるために重要である。
またV1は,V2,V3,V4を経て側頭連合野temporal association areaのIT野inferotemporal areaに至る腹側視覚路ventral visual pathwayの起点にもなっている。この経路が扱う情報は,小細胞系・微小細胞系に一部大細胞系の情報を統合した性質があり,物体の詳細な形態や質感・肌理などを分析して,その物体がどのようなカテゴリーに属し,自身との関係を知り,その視覚的記憶を保持・参照して認知的判断を行なうために重要である。 →側頭連合野 →頭頂連合野
〔佐藤 宏道〕
出典 最新 心理学事典最新 心理学事典について 情報
Sponserd by ![]()