デジタル大辞泉 「フィトクロム」の意味・読み・例文・類語
フィトクロム(phytochrome)
Sponserd by ![]()
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
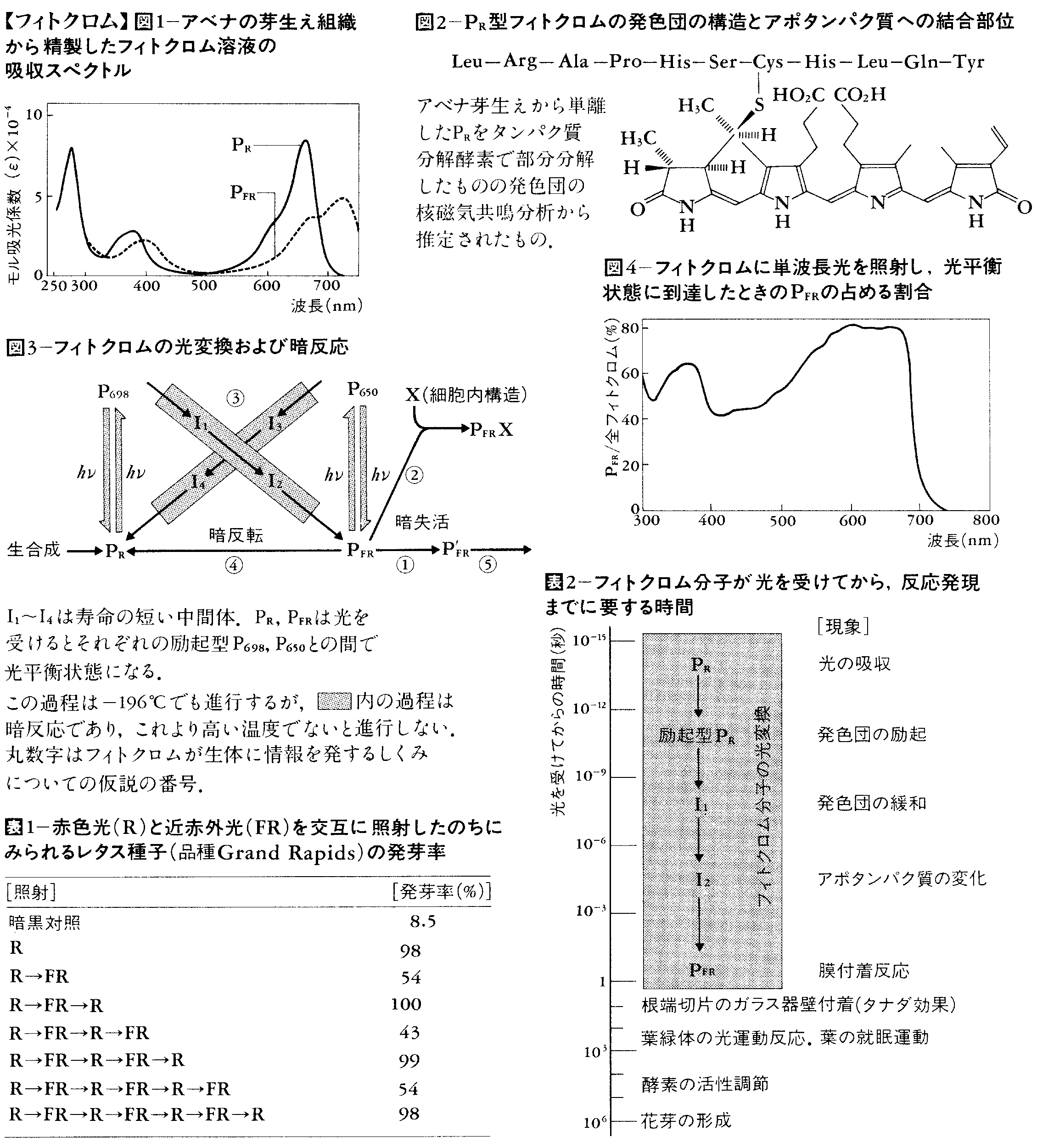
光を受けると,吸収スペクトルの相異なる型に可逆的に変わる性質をもつ色素タンパク質。菌類以外のすべての植物に存在し,環境の光条件を感知し,植物のさまざまな機能の制御に関与している。常温で280,380,666nmに吸収極大をもつ赤色光吸収型(PR)と280,400,725nmに吸収極大をもつ近赤外光吸収型(PFR)とがあり(図1),光を受けてPR⇄PFRの相互変換が起こる。
分子量12万4000。1種類のポリペプチド鎖から成る二量体で,沈降平衡法による分子量は約23万,沈降係数は約9sで,やや細長いタンパク質,アミノ酸組成はライムギとアベナ(カラスムギ)の黄化芽生えのものについて知られており,疎水性アミノ酸残基数は全残基数の約40%で,全体としては親水性のタンパク質である。二次構造は円偏光二色性より20%がα-らせん,30%がβ-構造,残りの50%がランダムコイルと推定されており,この点に関してはPRとPFRとの間に差がない。発色団は開環したテトラピロールで金属を含まず,C-フィコシアニンやアロフィコシアニンの発色団フィコシアノビリンとよく似た構造の胆汁色素で,常温では蛍光を発しない。単量体1個当り1個の発色団をもち,アポタンパク質のシステインとチオエーテル結合によって結ばれていると考えられている(図2)。
フィトクロムは抽出・精製の過程で,共存するタンパク質分解酵素による分解を受けやすく,発色団を含む分子量6万のポリペプチド断片(C)になる。Cは光可逆的吸収スペクトル変化を起こすしくみをもっているので,初期(1972年以前)の研究には専らCが用いられたが,これは親水性であり凝集や膜付着反応を起こさない。Cが除かれた残りの部分(B)はもともとCに比べ疎水性が高く,さらにPR→PFR変換による疎水性の高まり,高濃度PFRの2価金属イオンによる凝集のしくみをもつものと考えられている。
フィトクロム依存の生理学的反応は,PRとPFRの光可逆的変化が起こることによって引き起こされるのであるから,植物細胞はPRとPFRの違いを識別しているはずである。PFRはPRに比べ,より低い濃度の硫酸アンモニウムで塩析され,固定化されたヘキシル基やω-アミノオクチル基により強く吸着され,パルミチン酸に対するより高い親和性をもつこと,PFRでは分子の表面に単量体当り1個多くのヒスチジンとシステイン残基が現れていることから,PFRのほうが疎水性が高まっていると考えられ,また亜鉛や銅イオン添加による吸収スペクトルの変化および過マンガン酸による酸化についてのPRとPFRの比較からPFRでは発色団がより表面に露出していると考えられている。一方,PR⇄PFRにともなう発色団の構造の変化については,(1)光酸化還元,(2)光互変異性化,(3)シス-トランス光異性化の三つの説があるが,まだ確定はしていない。最近,精製したPRの溶液に赤色光を照射すると,pHが7.5より低い溶液ではフィトクロムによるプロトン吸収,pHが7.5より高い溶液ではプロトン放出が起こること,この変化は赤色・近赤外光可逆的であることが報告されている。
PR⇄PFR変換は,直接PR→PFRまたはPFR→PRへと変わるのではなく,寿命のごく短いいくつかの中間体を経て変換することが知られている(図3)。中間体を経る暗過程ではフィトクロム分子の水和が必要とされる。PRとPFRの間の光可逆的反応はどちらの方向に関しても一次反応である。ある光質の光を連続して照射したとき,PR⇄PFRは平衡に達して(光平衡photo equilibriumまたはphotostationary state),一定のPR:PFR比を示す。この比は各波長における吸光係数と量子収量とで定まる。単波長光を照射した場合の光平衡における全フィトクロムに対するPFRの比を図4に示す。カラシナ芽生えのアントシアン合成促進の場合のように近赤外光の連続照射が有効であることがしばしばある。このような場合は全フィトクロムのうちPFRの占める割合が大きいことが重要ではなく,少量のPFRが長時間にわたり存在することが必要と考えられる。
フィトクロムを含む試料に十分な(1)近赤外光または(2)赤色光を照射し光平衡に到達させ,それぞれの場合について一定波長間例えば660nmと730nmの間の吸光度の差⊿A=A660-A730を測り,次式のように(1)と(2)の場合の差を算出する。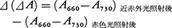
この値は試料のフィトクロム濃度に比例するので,溶液だけでなく組織中のフィトクロムの定量にも適用される。もう一つの定量法であるラジオイムノアッセーはPRとPFRを区別できないが,検出限界が分光的方法の1/200となっているほか,光化学的活性を失ったフィトクロムをも検出することができる利点がある。
生合成により生じるフィトクロムはPRのほうである。PRは暗所では長時間安定であるが,PFRは暗所においてPRへ戻ったり(PFR暗反転),徐々に光可逆性を失っていったりする(PFR暗失活)(図3)。カリフラワーの花序組織ではPFR暗反転のみが,イネやトウモロコシの子葉鞘(しようしよう)やアサガオ子葉ではPFR暗失活のみがみられるが,多くの植物では両反応が同時に起こる。PFR暗失活はCO,CH⁻,アジド,アンチマイシンAなどの呼吸阻害剤,嫌気条件やEDTAなどのキレート剤またはSH基をもつ試薬で阻害され,PFR暗反転はNA-DH,還元型フェレドキシンやNa2S2O4などの還元剤により促進される。PFR暗失活は単にフィトクロムが光化学活性を失うばかりでなく,タンパク質部分も変化を受けていることがラジオイムノアッセーの結果から示されている。白色光下で育てたアベナ芽生えに含まれるフィトクロムの量は黄化芽生えの1/100にすぎない。これは光によりPFRが生じ,それがPRより不安定であるということのほかにフィトクロムの生合成が抑制された結果と考えられる。黄化組織が赤色光を受けるとフィトクロムのmRNAの量が低下し,この変化には赤色・近赤外光可逆性がみられることから,フィトクロム自身の合成がフィトクロムで制御されていると考えられる。野外の植物では日暮れどきにはフィトクロムのかなりの部分はPFRになっているが,夜半にはほとんどがPRとして存在しており,このような変化が,日周的に起こる植物体内の種々の反応に意味をもっているものと考えられる。
1935年にスミソニアン研究所のフリントL.H.FlintとマッカリスターE.D.McAlisterがレタス品種グランドラピッドの種子の発芽が580~700nmの波長域の光で促進されることを見つけたのがフィトクロム発見への端緒となった。これを受けて52年にアメリカ農務省農学研究所のボースウィックH.A.Borthwickらは同研究所に大型スペクトログラフを建設し,光発芽の作用スペクトルを測定するとともに,発芽に最も有効な660nmの赤色光とその効果を打ち消す730nmの近赤外光を交互になん回か与えた場合,発芽率は最後に受けた光が赤色光であるか近赤外光であるかによって決まるという光可逆性現象を発見した(表1)。この反応の光受容色素が59年バトラーW.L.Butlerらにより発見されたフィトクロムであった。
フィトクロムは,外界から光を吸収することにより生じた色素分子内の変化を,信号として細胞に伝える光変換器の役割をしている。PFRが生理学的反応を引き起こす活性型であろうとの説があるが,PFRの機能はまだよくわかっていない。フィトクロムの光変換ののち多岐にわたる道筋を経てさまざまな現象が現れるが,現象の種類により数秒間で変化が認められるものから,多くの光形態形成反応の場合のように数時間から数日を要するものまである(表2)。切り取った根端を適当な溶液に浮かべ,赤色光を照射すると根端は正に帯電し,あらかじめ負電荷をもたせたガラス器壁に付着し,この変化には赤色・近赤外光可逆性がみられる(タナダ効果)。付着は光処理後20秒以内に起こり,フィトクロムによる最も速い生理学的反応の一つとされている。このほかフィトクロム依存の現象としては,赤色光によるNADキナーゼの活性化,フェニルアラニンアンモニアリアーゼの誘導,葉緑素,カロチノイド,アントシアンなどの色素,リブロース-1,5-ビスホスフェートカルボキシラーゼの小サブユニットのmRNAおよび集光性クロロフィルa/b-タンパク質複合体のアポタンパク質のmRNAの合成促進など転写,翻訳,酵素活性レベルでの調節から,オジギソウ小葉片の就眠運動促進のような運動現象の調節,コケ・シダ類の原糸体の光屈性反応,胞子・種子の発芽促進,胚軸鉤曲(こうきよく)部の伸展,オーキシンの極性移動,伸長生長,花芽分化の促進または抑制のような生長・発育現象の調節にいたるまで,きわめて多くのものが知られている。細胞小器官でみられるフィトクロム依存反応としては,黄化組織から単離したミトコンドリアのNADP還元反応,エチオプラストからのジベレリン様物質分泌の赤色光による促進,赤色光照射を受けた組織から単離した細胞小器官でみられる変化としては,ミクロソームのペルオキシダーゼ,アデノシントリホスファターゼやアデニル酸キナーゼ活性の阻害,ミトコンドリアの添加NADH酸化反応の促進,カルシウムイオンCa2⁺取込みの阻害などが知られている。フィトクロムの光変換後に起こる代謝的変化のうち,どれが光形態形成の原因となりどれが副次的なできごとにすぎないかは,今後の課題として残されている。
光変換したフィトクロムの作用機作としては(1)PFRが活性をもち他の系に働きかける(PFR活性説)。とくに遺伝子活性を調節する場合は遺伝子活性化説と呼ばれる,(2)PFRが生体膜へ付着し膜の機能を変化させる(膜付着説),(3)励起型のP698,P650がそれぞれPFR,PRへと変化する過程(緩和過程)を経る回数が作用を受ける他の系にとって重要(情報ポンプ作用説),(4)暗反転の速度が計時機構として働く(PFR暗反転説),(5)暗失活したPFRが他の系に働きかける(暗失活PFR説)などがあるが,どれもまだ確定的ではない(図3)。(4)に対しては,フィトクロムは単に光変換器として働き,光信号を受けて生体時計の位相設定をする役割を担うという説がある。
高濃度の単離PFRは他の生体成分がまったく存在しなくてもCa2⁺やマグネシウムイオンMg2⁺など2価の金属イオンが存在すると凝集し,近赤外光照射によりPRに戻すと解離する。凝集条件で生体膜を加えればPFRはそこへ付着する。黄化組織内のフィトクロムはその数%が膜画分中に得られるが,組織を赤色光照射すると可溶性画分のフィトクロムは減少し膜画分のフィトクロムは増加し,この変化は赤色・近赤外光可逆的である。同様の変化は組織抽出液を光照射した場合にも起こる。フィトクロムは膜画分中のタンパク質に付着するものと考えられている。免疫細胞化学的手法でフィトクロムの細胞内分布の変化が調べられ,黄化組織の細胞ではフィトクロムは細胞質基質に均一に分布しているが赤色光を照射すると細胞質のところどころにかたまって存在するようになること,近赤外光照射で元の状態へ戻ることが示された。ヒザオリの葉緑体の光運動反応やホウライシダの原糸体の光屈性反応では赤色光,近赤外光ともに偏光効果が認められることから,これらの場合にはフィトクロムが一定の配向で細胞膜に組み込まれているものと考えられている。
自然界には多くの色素が存在し,それぞれ固有の光受容反応を行っているが,代謝制御から形態形成に至る広範な現象を受け持つ制御専業分子としてのフィトクロムの出現は,色素の進化という面からきわめて興味深い課題を提供している。
執筆者:辻 英夫
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
植物が光(赤色光)の刺激に反応するとき、その光の受容体となるタンパク質色素のことで、Pr型とPfr型の二つの型が存在する。Pr型はおもな吸収極大が660ナノメートル付近にあり、Pfr型は720ナノメートル付近にある。Pr型フィトクロムは赤色光を吸収するとPfr型に転換し、Pfr型は近赤外光を吸収するとPr型に戻り、この二つの型は相互に可逆的に転換できる。ある種の植物では、Pfr型のフィトクロムは暗黒中で徐々にPr型に戻ることができる。フィトクロム分子の発色団は開環したテトラピロールで、藍藻(らんそう)類の色素フィコシアニンとよく似ている。フィトクロムはすべての植物に含まれ、光に関係するさまざまな形態形成や、生体の生理的諸機能の調節にあずかっている。
[勝見允行]
レタスの一品種であるグランド・ラピッズGrand Rapidsの種子は光発芽種子であるが、アメリカのフリントL. H. FlintとマカリスターE. D. McAlisterは1935年、その発芽には赤色光がもっとも有効である反面、近赤外光は逆に阻害的であることを報告した。やがて1946年になると、アメリカのパーカーM. W. Parkerらは、短日植物であるオナモミやビロキシダイズの花芽誘導暗期における光中断効果は、赤色光がもっとも有効であることを発見した。その後、アメリカ、メーン州ベルツビルにある農務省研究所のボースウィックH. A. BorthwickとヘンドリックスS. B. Hendricksの研究グループは、これらの赤色光の作用についての研究を進展させ、赤色光と近赤外光とが相互に可逆的効果をもつことをみいだした(1952)。前記のレタス種子を暗黒中で吸水させ、赤色光を短時間照射すると発芽が誘導されるが、赤色光照射直後に近赤外光を照射すると、赤色光効果は打ち消されてしまうというのもその一例である。この可逆的効果は何度も繰り返すことができる。このようなことから、赤色光および近赤外光に対して、それぞれ最大吸収をもつような相互に転換可能な一つの色素が想定され、これをフィトクロムとよんだわけである。1959年アメリカのバトラーW. L. Butlerらは、トウモロコシの黄化芽生えからフィトクロムの抽出に成功した。
[勝見允行]
フィトクロムは、Pfr型が生理的に活性であるが、容易に別の型(Pfr´)に不活性化される。フィトクロムがPfr型になることが、どのようにしてさまざまな形態形成作用に結び付くのかは、まだ明らかにされていない。しかし、植物ホルモンの量的変動、酵素の合成や活性化を伴う場合のあることが知られており、おそらく、なんらかの生化学的変化を介するものと考えられている。
[勝見允行]
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
植物の生長や形態形成の過程には,弱い赤色光と近赤外光の短時間照射によって可逆的に制御できる反応,すなわち,赤色光効果のみられる現象が多数ある.フィトクロムはこれらの可逆的反応の光エネルギー受容体として,1956年,S.B. Hendricks,W.L. Butler,S.B. Borthwickらによって発見され,抽出された色素タンパク質で,すべての植物に存在すると考えられている.フィトクロムが関与する反応としては,胞子,種子の光発芽,葉の展開,花芽の形成,クロロフィルおよびカロテノイドの生合成,細胞分裂の時期決定,生体時計の時刻合わせ,屈光性の感度変化などの多岐にわたっている.フィトクロムは分子量4×104 のサブユニットから構成されるタンパク質部と,開環したテトラピロール誘導体を発色団とする部分とからできており,赤外光吸収型フィトクロムと近赤外光吸収型フィトクロムがあり,これらは光を受けると相互に可逆的に変換する.
出典 森北出版「化学辞典(第2版)」化学辞典 第2版について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
…この際,内在リズムの位相そのものも光中断によって変化すると考えられる。光中断に有効な光は赤色光であり,この効果は近赤外光の照射によって打ち消されることから,光中断に関与する物質はフィトクロムであると推定されている。フィトクロムは赤色光あるいは近赤外光の照射によって,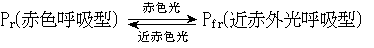 の可逆的変化を示すことがわかっているからである。…
の可逆的変化を示すことがわかっているからである。…
※「フィトクロム」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
3月から 5月頃に発生する雷。寒冷前線の通過時に発生する界雷で,この雷雨はよくひょう(雹)を伴う。春の到来を伝える雷ともいわれる。雷鳴に驚き冬眠していた地中の虫たちが目ざめるという理由で「虫出しの雷」...