
内科学 第10版 の解説
心筋イオンの動態と心電図波形の成立(心血管代謝と機能)
(1)ペースメーカと非ペースメーカ細胞
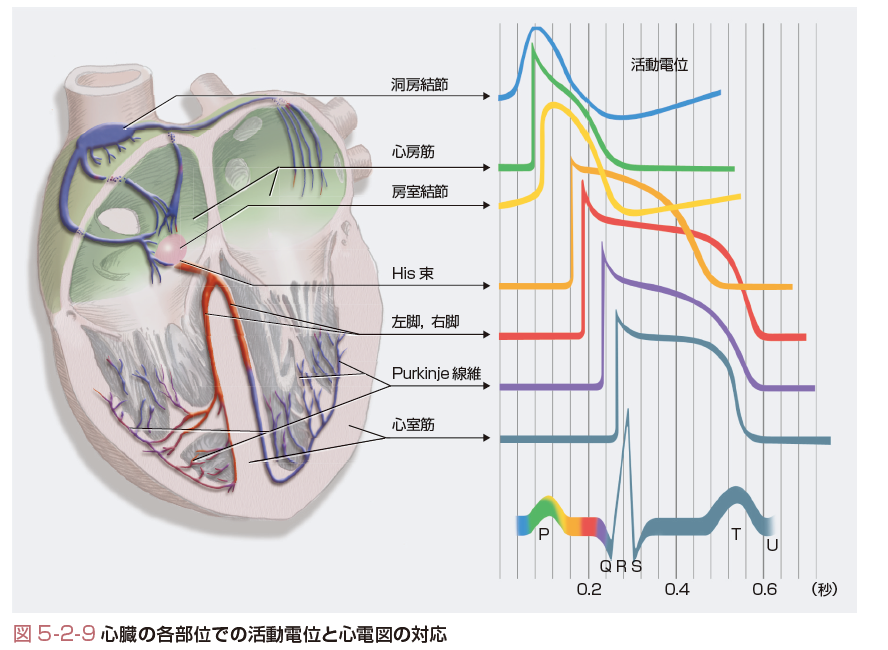
心臓には大きく分けて2種類の細胞があり,活動電位の形態も大きく異なる(図5-2-9).自動能を有するペースメーカ細胞とそれ以外の細胞である.ペースメーカ細胞は,洞房結節や房室結節,心室内の刺激伝導系に認められ,共同して,心臓刺激伝導系を構成している.一方,非ペースメーカ細胞は心房や心室の作業筋を構成しており,自動能をもった細胞群からの刺激があると活動電位を発生し収縮に寄与する.病的状態では,これらの非ペースメーカ細胞も自動能をもつようになり,したがって病的なペースメーカ細胞として働く.
(2)Nernst式により求められるイオンの平衡電位
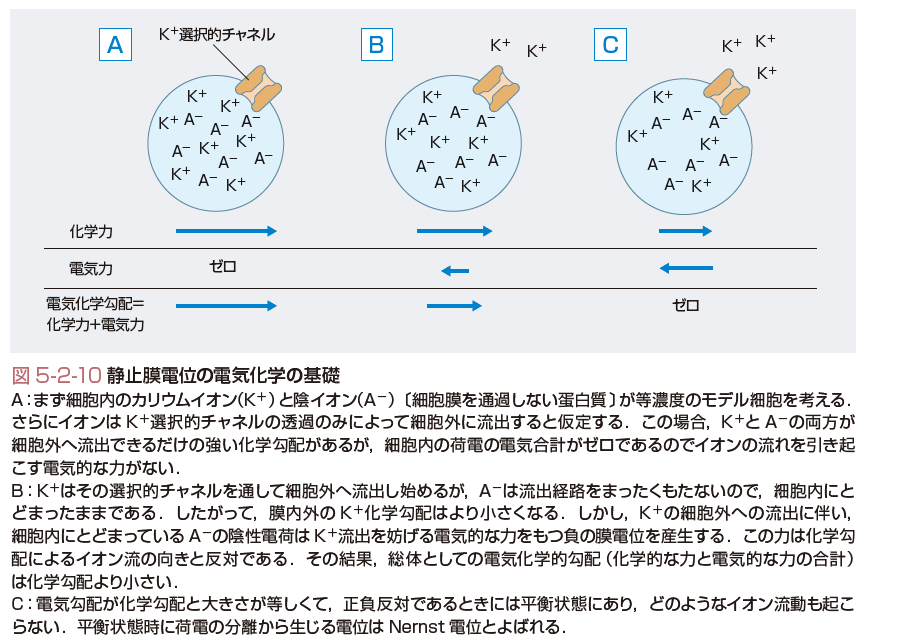
活動電位を決定するNa,K,Caイオンなどの分布は細胞内外で均等ではなく,膜に発現するトランスポータやポンプ(Na/K ATPase)などが協調して働き,細胞内のNaやCaを細胞外に汲み出して,Kイオンを細胞内に取り込むので,細胞膜を介したイオン濃度の生理的な勾配がつくられている.この差が膜電位を決定している.たとえば,Kイオン濃度は細胞内が140 mM,外が4 mM程度なので,もし膜が脂質二重層ではなくてイオン透過が100%可能であるとすれば,Kイオンは等モルになるまで細胞外に向けて流れ出ることになる.これに拮抗するように電気的な印加をすることにより,Kイオンの膜を介する動きを止める(平衡状態とよばれる)ことができる電位が,Nernst式で求められる平衡電位の定義である(図5-2-10).逆にいうと,細胞膜に発現している親水性でイオンを通すことのできるKチャネル蛋白が開口すると,Kイオンは自分のK平衡電位(EK=−95 mV)に膜電位が到達するように働くことになる.ちなみに,で与えられる.RT/zFの値は定数で,37℃,+1の電荷(Kイオンなど)では,23.7 mVである.[Kin]と[Kout]は,細胞内外のKイオンの濃度( mM)である.
(3)活動電位の形成
心筋細胞の膜電位は,どの程度の数のチャネルが,その時点で活性化するか,そのコンダクタンス(チャネルのイオンの通しやすさ),また,どのくらいの時間,活性化しているかによって決まってくる.興奮していない(静止時の)細胞は相対的にKイオンに対する透過性が高く,NaやCaイオンに対しては低いため,静止膜電位はK平衡電位に近い値となる.この電位付近では背景(background)K電流(IK1)が,内向き整流Kチャネルにより運ばれており,このチャネルはKCNJ2という遺伝子によりコードされている.最近,このKCNJ2の変異がチャネル機能を変化させるために,いくつかの遺伝性不整脈を起こすことが知られている(Hille, 2001;Zipesら,2009;Gussakら,2008).
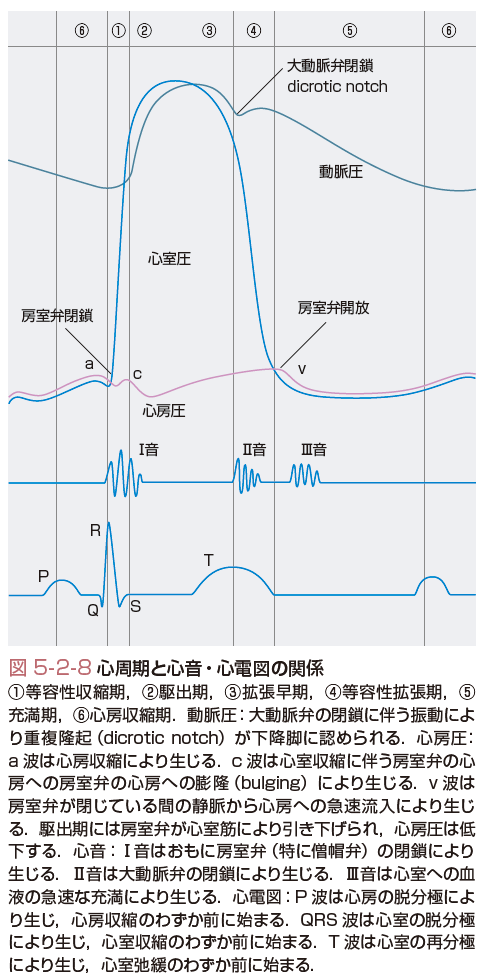
膜を介して動くイオンは少量でも容易に膜電位は変化するため,細胞膜内外のイオン濃度勾配は比較的一定しており,したがって,心周期の間に平衡電位に大きな変化はない.心筋活動電位は,非常に持続時間が長く,500 msec近くある.この結果,心臓はポンプとして働くことのできる長い収縮時間を得ることができる.一方,洞房結節は毎分60~100の間で規則正しく脱分極して作業心筋に興奮を送り,心房と心室は順次に興奮して全身に血液を送り出すポンプとして働く(図5-2-8,5-2-9).図5-2-9は心臓の各部位での活動電位と心電図の対応を示しているが,心房と心室筋の興奮が,それぞれ心電図のP波とQRS波に対応し,QT時間は心室筋活動電位の長い脱分極時間を反映している.
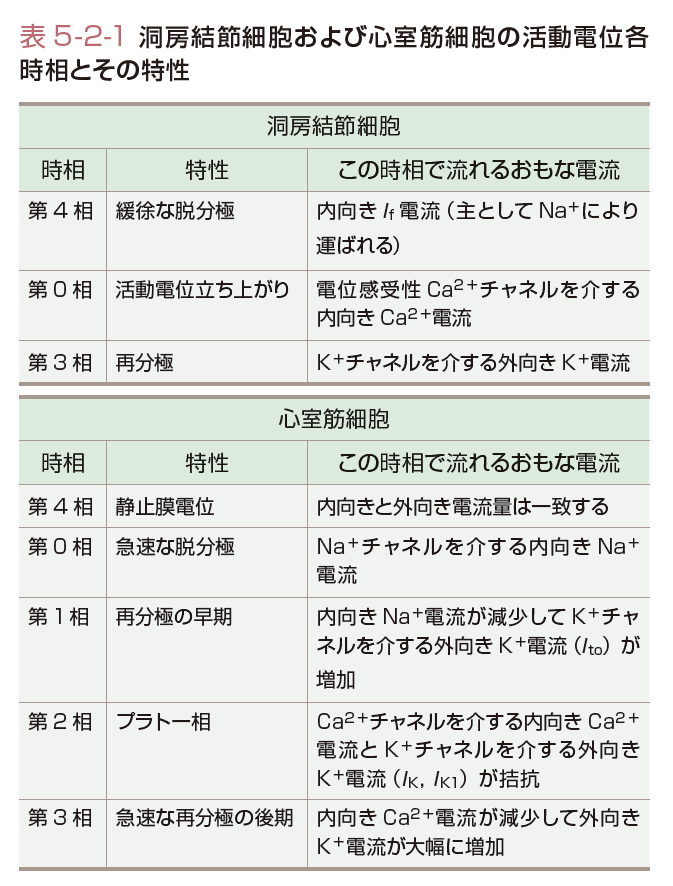
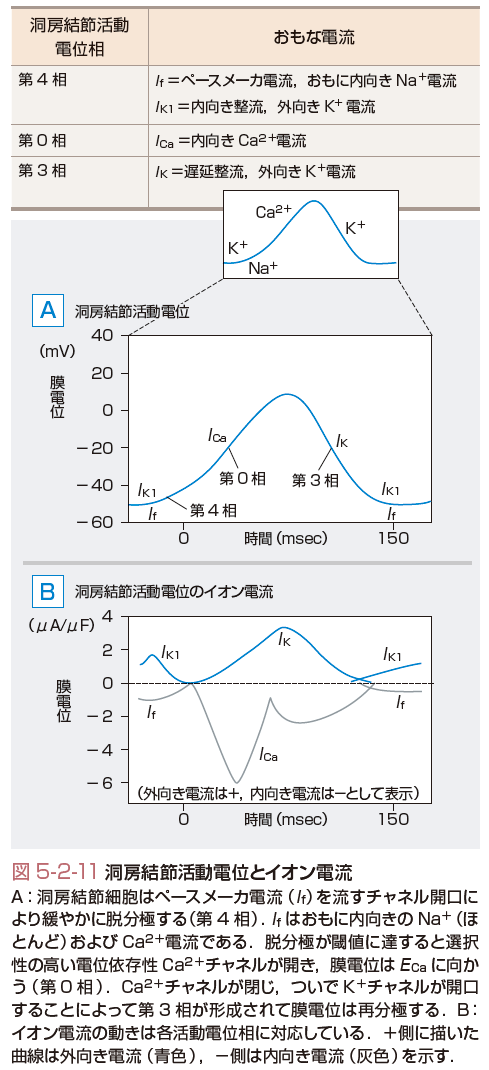
さて,洞房結節の活動電位は常に振動しており,第0,3,4相がある(図5-2-11,表5-2-1).つまり,心室筋のプラトー相(2相と3相)がない.さらに,第4相は,緩徐な脱分極を指し,この脱分極は,主としてNaイオンにより運ばれるIf電流により惹起される.一方,0相はL型Caチャネルの開口により形成され,Caの平衡電流(+159 mV)に近づくように膜電位は変化する.3相では,Caチャネルはゆっくりと閉じてKチャネルが活性化することにより再分極が起こる.そして膜電位が−60 mV前後になるとIfチャネルが活性化して再び脱分極が始まる.洞房あるいは房室結節の活動電位は細胞数が少ないため心電図で検出することはできない.
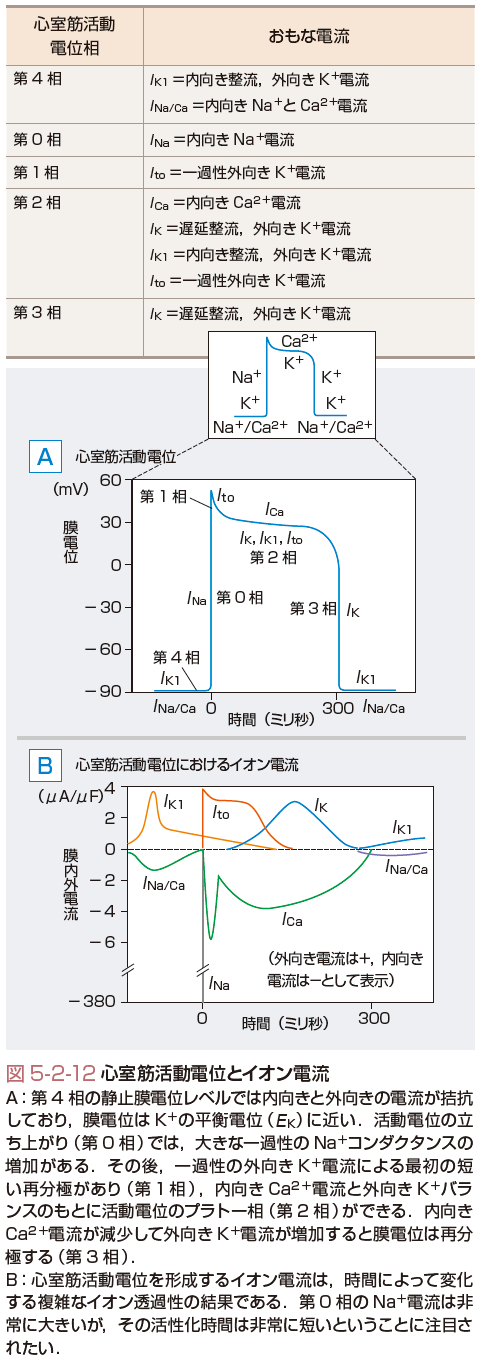
一方,作業心筋細胞の活動電位は第0から4相まで有している(図5-2-12,表5-2-1).図5-2-12に心室筋の活動電位に関与するイオン電流をまとめる.その静止膜電位はKの平衡電位付近であり,近傍の刺激伝導系からの脱分極刺激を受けて活動電位を発生する.まず,膜電位依存性Naチャネルを介する内向きのNa電流が一過性に増加するため心室筋活動電位0相の急速な脱分極が起こる.一方,洞房結節の0相は異なるイオンすなわちCaイオンにより形成される.Naチャネル(INa)の開口は急速なNaイオンの流入を起こし,これにより細胞の膜電位は,Naイオン平衡電位(ENa)である+60 mVに膜電位は近づくように脱分極を始める.しかし,Naチャネルは速やかに不活性化するため,通常Naコンダクタンスの増加は1~2 msecしか持続しない.心電図では一番大きな波であるQRS波の始まりに対応している.
さて,Naチャネルが膜電位あるいは時間依存性の不活性化状態から回復するまでの時間を心筋細胞の不応期(refractory time)とよぶ.この不応期の間はほかの細胞からの刺激があっても脱分極できない.この機序により心臓は収縮のための十分な時間を確保している.不応期は活動電位の立ち上がりから再分極過程の終わりまでで,Na電流は興奮伝導の速度を決定する重要な因子である.心電図(図5-2-9)では,QT時間に対応している.
活動電位の急速な立ち上がりは,いったん+20 mV付近に急速に再分極する.これが第1相で,心電図ではQRS波の終わり部分に対応する.近年,特発性心室細動との関連がいわれているJ波の出現もこの時間に相当しており,第1相の形成には,2つの機序,すなわち①急速なINaの不活性化と②一過性K電流(Ito)の活性化が関与する.
第2相は活動電位のプラトー相(図5-2-12)で,2種類のCaチャネル電流(ICaT,ICaL)により運ばれる内向きCa電流と数種類のKチャネルにより運ばれる外向きK電流(IK,IK1,Ito)との微妙なバランスで決定される.このバランスを保つのに数百個のチャネルしか活動していない.プラトー相においては,このように少ない数のチャネルしか開口していない,逆にいうと膜抵抗が非常に高い(低いコンダクタンス).実は,これは心臓にとって大切なことである.つまりプラトー相の心筋を電気的に保護するとともに,効率よく(少しの電気的興奮で)活動電位を心室全体に伝えることができる.心電図でみると,この時間はやはり前述のQT時間に対応する.
最後にプラトー相でKチャネルが運ぶ外向き電流は,内向きCaチャネル電流に対抗し,再分極に働く.Ca電流が不活性化し,外向きK電流が活性化して,膜電位をEKに引き戻すことにより,再分極相すなわち第3相のはじめが形成される.IKチャネルは,しかしながら−40 mV付近で脱活性化するため,EKまでは再分極することができない.第4相の深い静止膜電位は,実際上,時間非依存性K電流(IK1)により再び形成され,1心周期が完結する.これらの個々の心筋細胞の興奮は全体の電気活動として心電図(図5-2-9)に記録される.このように活動電位の裏側にあるイオンチャネルやイオン電流の働きを理解しておくことは,心電図や不整脈を学習するために大変役立つものである.[堀江 稔]
■文献
Gussak I, Antzelevitch C: Electrical Diseases of The Heart, Springer-Verlag, London, 2008.
Hille B: Ionic Channels of Extable Membranes, 3rd ed, Sinauer Associates, Sunderland, 2001.
Zipes DP, Jalife J: Cardiac Electrophysiology: From Cell to Bedside, Saunders, Philadelphia, 2009.
出典 内科学 第10版内科学 第10版について 情報
Sponserd by ![]()