デジタル大辞泉 「霊長類」の意味・読み・例文・類語
れいちょう‐るい〔レイチヤウ‐〕【霊長類】
Sponserd by ![]()
Sponserd by ![]()
更新日:
出典 共同通信社 共同通信ニュース用語解説共同通信ニュース用語解説について 情報
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
霊長目(サル目)Primatesに属する種の総称。霊長とは,万物の霊長の意である。食虫目に類似の動物から分化し,樹上生活を通じて適応放散した一群と考えられている。現生霊長目は,原猿亜目Prosimiiと真猿亜目Anthropoideaの2亜目に分けられる。原猿亜目は,さらにメガネザル下目Tarsiiformes,ロリス下目Lorisiformes,キツネザル下目Lemuriformesの3下目に分けられる。ただし,メガネザルは真猿亜目の姉妹群であるため,分岐分類では,メガネザルを除く原猿類を曲鼻猿亜目Strepsirrhini,メガネザルと真猿類を合わせて直鼻猿亜目Haplorhiniとする。真猿亜目は,中南米に棲息する広鼻下目と,アジア,アフリカに棲息する狭鼻下目に分けられる。前者は,オマキザル上科Ceboidea,後者はオナガザル上科Cercopithecoideaとヒト上科Hominoideaからなる。科のレベルで見ると,原猿は,メガネザル科Tarsiidae(メガネザル下目),ロリス科Lorisidae,ガラゴ科Galagidae(ロリス下目),コビトキツネザル科Cheirogaleidae,キツネザル科Lemuridae,インドリ科lndrildae,アイアイ科Daubentoniidae(キツネザル下目),また真猿はクモザル科Atelidae,オマキザル科Cebidae(広鼻下目),オナガザル科Cercopithecidae,テナガザル科Hylobatidae,オランウータン科Pongidae,ヒト科Hominidae(狭鼻下目)に分けられる。なお,広鼻下目の分類については見解の相違があり,ここに示したものはそれらの一つである。この13科には,約220種の現生種が含まれている。ヒトHomo sapiens(ヒト科)はその中の1種で,直立二足性,際だって大きな脳,言語によるコミュニケーション,家族を基本単位とした高度の社会性など,著しい特殊化を遂げた種であるが,原猿類の中には第三紀初頭の初期霊長類の姿をほうふつとさせるような原始的な種も含まれており,一つの目の中にきわめて原始的な種と,著しい特殊化をとげた種を同時に含むという点も,この目の大きな特色といえる。
霊長類を他の哺乳類から分ける重要な特性は,いずれも樹上生活と強く結びついており,それらは手足と目に集中している。まず,樹上,とくに細い枝先での体の安定を保つために,その手足は母指を他の4指に対向させて,物を把握することが可能になっている。他の多くの哺乳類では,手は単なる歩行器官でしかないが,霊長類では歩行器官であると同時に保持器官になっている。鈎爪(かぎづめ)から平爪(ひらづめ)への変化,指掌紋(ししようもん)の発達なども霊長類に見られる大きな特色であるが,上記と一連の機能的意味をもった形質といってよい。次に,両眼視,つまり立体視の可能な目をあげることができる。樹上という三次元空間で活動しなければならない霊長類にとって,遠近の目測の必要性は容易に推測しうるところである。両眼視が可能な目は,頭部側方から正中に寄り,視野の重複域を拡大する。現生霊長類では,すべての種において眼輪(がんりん)(眼窩(がんか)のふち)が閉じ,真猿類ではさらに眼窩後壁が形成されている。さらに,口顎部の短縮などの特徴も目の変化と関連している。そして,嗅覚(きゆうかく)依存から視覚依存への移行傾向が認められ,このことは脳の発達を促す基盤となるとともに,霊長類の社会性進化において重要な鍵となった。
しかしながら,霊長類の諸特性の起源が樹上生活と結びついていることは間違いないものの,一連の特徴が霊長類以外の樹上性哺乳類に必ずしも見られないことにも注意しなければならない。この点を補うため,単に樹上で運動することではなく,樹上で昆虫を捕獲して食べることが淘汰圧になったとする意見が出されている。また,一連の特徴が同時に進化したのではなく,まず,把握性の手足が進化し,その後に両眼視の発達があったとする仮説も出されている。
こうした諸特性は,霊長類のすべての種が共通してもちあわせているわけではない。むしろこれらは,霊長類全体を眺めたときの進化の方向性としてとらえたほうがよいかもしれない。つめを例 とってみても,原猿は,少なくとも後肢の第2指には鈎爪を残しているし(化粧爪,あるいはtoilet claw),マーモセット亜科では足の親指以外に鈎爪が二次的に進化している。進化の方向性ということであれば,必ずしも霊長類の定義の項目に加えることはできなくても,さらに多くの特性の抽出が可能だろう。妊娠期間や育児期間の長期化,産子数の減少(真猿類ではマーモセット類を除いて1産1子),夜行性から昼行性への変化,単独生活から集団生活という移行なども,霊長類がたどった特殊化の方向性を示している。そして脳の発達もその例外ではない。
霊長類の歯数は,特殊化と共に少なくなっている。原始的な哺乳類の歯式は,片側の上顎で切歯3本・犬歯1本・小臼歯4本・大臼歯3本,片側の下顎で切歯3本・犬歯1本・小臼歯4本・大臼歯3本の計44本(以降,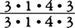 =44本と略記する)であるが,霊長類ではこれと同構造の歯式をもつものは,化石種にも現生種にも存在しない。すべてはこれよりも少ないのである。始新世のオモミス上科Omomyoidea,アダピス上科Adapoideaの初期のメンバーは,
=44本と略記する)であるが,霊長類ではこれと同構造の歯式をもつものは,化石種にも現生種にも存在しない。すべてはこれよりも少ないのである。始新世のオモミス上科Omomyoidea,アダピス上科Adapoideaの初期のメンバーは,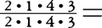 40本をもつ。より新しい時代になると,小臼歯を1本減らした種も現れる。現生原猿類では,メガネザル類が
40本をもつ。より新しい時代になると,小臼歯を1本減らした種も現れる。現生原猿類では,メガネザル類が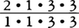 =34本,ロリス下目が
=34本,ロリス下目が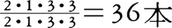 ,キツネザル科もロリス類と同じだが,イタチキツネザル属Lepilemurだけが上顎切歯を完全に欠いている。また,インドリ科は下顎犬歯を,アイアイDaubentoniaは上下犬歯と下顎小臼歯を欠き,上下切歯と上顎小臼歯はいずれも1対だけ
,キツネザル科もロリス類と同じだが,イタチキツネザル属Lepilemurだけが上顎切歯を完全に欠いている。また,インドリ科は下顎犬歯を,アイアイDaubentoniaは上下犬歯と下顎小臼歯を欠き,上下切歯と上顎小臼歯はいずれも1対だけ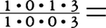 18本という歯式をもつ。広鼻猿類は多くがであるが,マーモセット亜科では
18本という歯式をもつ。広鼻猿類は多くがであるが,マーモセット亜科では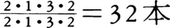 ,オナガザル上科とヒト上科は
,オナガザル上科とヒト上科は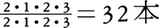 である。最初期の霊長類はおそらく40本の歯式をもっていたと考えられており,アイアイなどに見られる欠落は,系統群固有の特殊化とみなしてよい。
である。最初期の霊長類はおそらく40本の歯式をもっていたと考えられており,アイアイなどに見られる欠落は,系統群固有の特殊化とみなしてよい。
化石霊長類は新旧両大陸にまたがって広範な分布を示すが,ヒトを除く現生種の分布域は,人為的移入を除き,ヨーロッパと北アメリカを含まない。またオーストラリアとニュージーランドは,霊長類の歴史とは無縁の土地であった。原猿類の分布は旧世界(旧大陸)に限られる。メガネザルTarsiusは東南アジアに,ロリスLorisはインド,スリランカ,インドシナ半島とアフリカ大陸に,ガラゴはアフリカ大陸に分布する。キツネザル類の分布はマダガスカルとその周辺の小島に限られる。オマキザル上科Ceboideaのサルたちは,メキシコ中部以南の中央アメリカと南アメリカに広く分布するが,南限はアルゼンチン中部である。オナガザル上科Cercopithecoideaは,日本,東南アジアの島々,中国からインドシナ半島とインド亜大陸を経てインド・パキスタン国境付近まで,そしてアフリカ大陸とアラビア半島の一部に分布する。類人猿は,テナガザル科Hylobatidaeが中国南部とインドシナ半島,そしてスマトラとボルネオに, オランウータンPongo pygmaeusはスマトラとボルネオに分布する。アフリカの3種の大型類人猿は,チンパンジーPan troglodytesが西アフリカのセネガルからタンザニア西部まで赤道をたすきがけにしたような分布を見せ,ザイール川左岸(=南側)の森林にはボノボPan paniscusが分布する。ゴリラGorilla gorillaは,アフリカ中央部の低地熱帯林帯のカメルーンやガボンにニシローランドゴリラ,コンゴ東部にヒガシローランドゴリラが,西部大地構帯沿いの山地帯にはマウンテンゴリラが分布している。ヒトを除く現生霊長類の分布域中,その北限を占めるのはニホンザルMacaca fuscataで青森県下北半島の北端に近い北緯41°30′が北限地,南限はチャクマヒヒPapio ursinusの分布域であるアフリカ大陸南端の南緯35°である。
霊長類が森林ときってもきれない関係にあることは,新旧両大陸の熱帯多雨林が霊長類の分布の中心になっていることからも明らかであろう。原猿とオマキザル上科Ceboideaのサルのほとんどは,熱帯林の中の樹上生活者である。原猿での例外といえば,乾燥サバンナに分布するショウガラゴGalago senegalensis,疎林帯に分布を広げているオオガラゴOtolemur crassicaudatusやベローシファカPropithecus verreauxiなどであろう。オナガザル上科に至って,地上性の種,森林から脱出した種が見られるようになる。パタスモンキーErythrocebus patas,ヒヒ属Papio,ゲラダヒヒTheropithecus gelada,マカク属Macacaなどで地上性はとくに顕著であり,パタスモンキーは草原に,5種のサバンナ性のヒヒはサバンナと半砂漠に,ゲラダヒヒは高地草原に生息している。ニホンザルのように寒冷な落葉広葉樹林帯にまで分布を広げている種もある。また,マウンテンゴリラは森林限界を越えたアフロ・アルパイン帯にまで分布を広げているし,チンパンジーの分布域は乾燥疎開林帯にまでのびている。
現生霊長類の多くは雑食性である。しかし,栄養価は低いが,労せずして大量に採食できる植物食に偏食化した種が,各分類群中に見られる。原猿では,イタチキツネザル属Lepilemur,インドリ属Indri,シファカ属Propithecusなどがその例であるし,クモザル科ではホエザル属Alouattaが菜食,クモザル属Atelesは果実食の傾向が強い。オナガザル科のオナガザル亜科Cercopithecinaeは雑食性だが,コロブス亜科Colobinaeに属する種は一般に菜食で,アジアの森林に住むラングールlangurはリーフモンキーleaf monkeyとも呼ばれる。類人猿では,オランウータンが果実食への偏りを見せ,ゴリラは繊維質の植物を多く利用する。このような偏食主義は,一般に雑食性を保っている近縁種に比して体が大型化している。
霊長類の食性について,いくつかの注目すべき現象をあげておこう。樹脂は,夜行性の原猿ポットーPerodicticusなどの小型の原猿類にとって,見逃すことのできない重要な食物に違いない。ニホンザルの食物メニューは群れごとに異なっており,それらは各群れで伝承されていることが明らかにされている。深雪地帯のニホンザルは,冬の間は広葉樹の樹皮の形成層に食生活を依存している。チンパンジーは,アリやシロアリのほか,小型・中型の哺乳類を捕食する。採食行動にも多くの興味深い現象が記録されており,ニホンザルのイモ洗い行動,チンパンジーの道具を用いてのアリ釣りやシロアリ釣り,石で堅果を割る行動などはいずれも文化的な伝統として注目された。また,ボノボとチンパンジーに見られる食物を分配する行動は,人類における分業の発生と研究する見地からも,きわめて重要な意義をもっている。
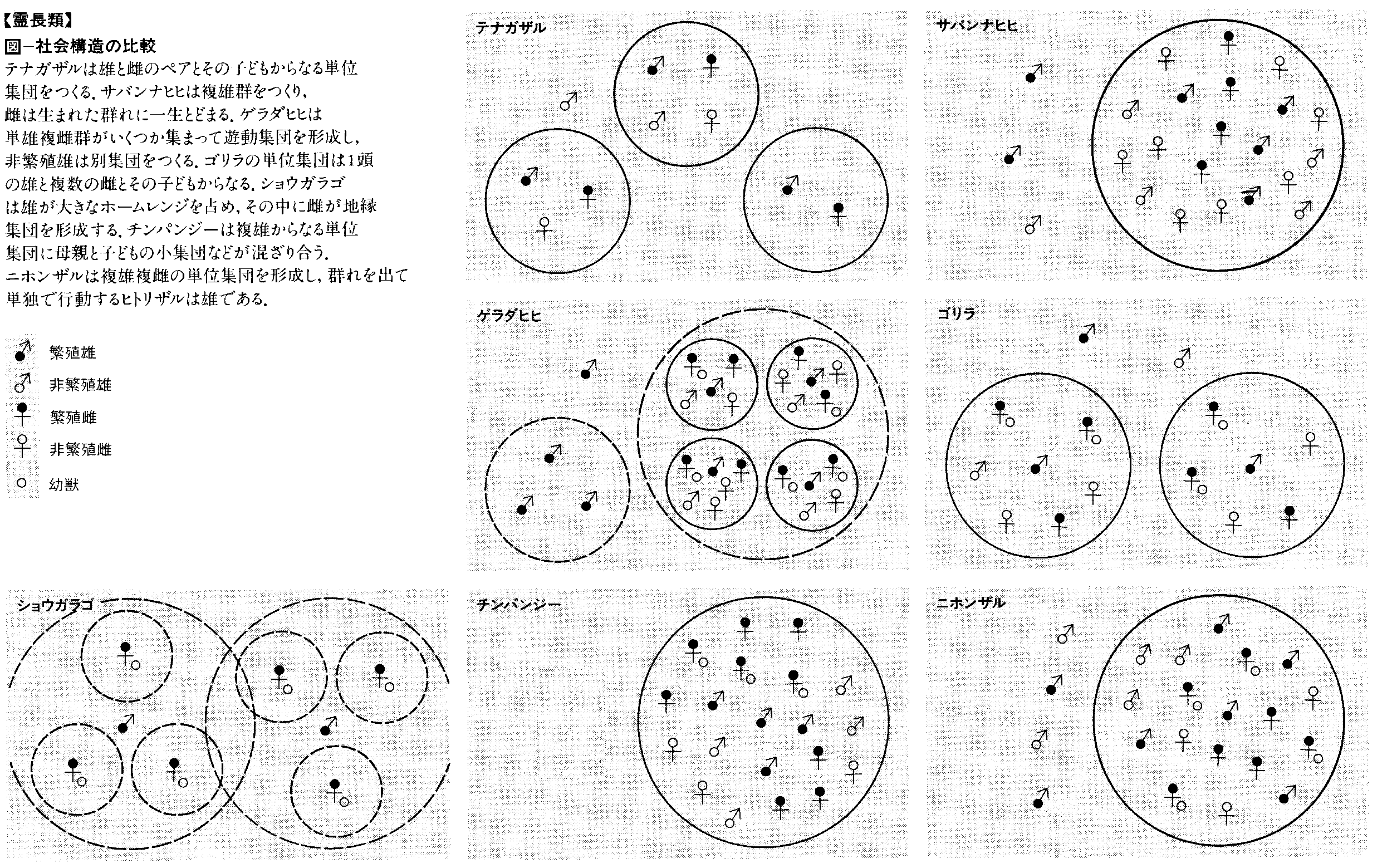
原猿の約半数は夜行性で,ガラゴGalago,コビトキツネザルCheirogaleus,アイアイDaubentoniaなどは巣をつくって,日中はその中で眠っている。現生霊長類中,〈巣〉をつくるのは,これらの原猿と大型類人猿のみである。しかし,後者の〈巣〉は毎日夕刻に新たにつくられ,巣というよりもベッドと呼ぶほうが適切である。夜行性の原猿の多くは単独行動者である。わずかにメガネザルTarsiusとアバヒAvahiが集団をもつが,これらはいずれも雌雄各1頭の結びついたものであるから,このようなペアの構成が霊長類の単位集団の原型的なものと考えることができる。ヨザルAotusは真猿中唯一の夜行性の種であるが,これもペアの集団をもっている。昼行性の種は,オランウータンという唯一の例外を除いて,すべて安定した単位集団(社会)をもっている。霊長類の単位集団は両性からなり,種に特異な構成とサイズをみずから保つ半閉鎖的な集団で,特定の構成要素(メンバー)を放出し,また外部から受けいれる。したがって霊長類の一つの種社会は,ある社会的な距離を保って散在する単位集団と,その空隙を彷徨(ほうこう)する単独行動個体によって模式化することができる。単位集団は構成によって,単雄単雌,単雄複雌,複雄複雌に分けることができる。単雄単雌の集団は上述の夜行性霊長類のほか,インドリ,テナガザルなどに見られる。これらは,母系によっても父系によっても継承されることのない集団だといってよい。単雄複雌と複雄複雌の集団には母系と父系の2型があり,クモザル亜科とアフリカ類人猿以外は,ほとんどすべてが母系的である。すなわち,集団内で生まれた雄は集団を離脱し,他集団に加入する。単雄の場合にも,そのただ1頭の雄は数年に一度の周期で外部の雄との入れ替えが行われる。雌は原則として出自集団を離れることはなく,一つの集団は母系血縁個体によって継承される。ゴリラの単位集団は単雄複雌で非母系,チンパンジーおよびボノボの単位集団は複雄複雌で父系的である。これらに共通するのは,雌が性成熟に達するまでに出自集団を離れる点で,これは母系的な集団とは対照的である。ゴリラでは雄の子も集団を離れて集団内の単雄の構成を保つが,チンパンジーでは雄が集団を離れることはなく,出自を同じくする雄たちの強いきずなが集団の核をなしている。霊長類社会のこれらの社会構造は,近親交配の回避と深い関係をもっている。オナガザル類の中には,マントヒヒPapio hamadryasやゲラダヒヒTheropithecus geladaなどのように,重層の社会構造をもつものが知られており,後者はときに数百頭にのぼるハード(群れ)herdをつくって遊動することがあるが,その中で単位集団の輪郭は厳重に守られている。しかしこのような例はまれで,一般に霊長類の集団間関係には厳しい対立が見られ,地域社会としての構造化の達成されていない例が多い。チンパンジーでは,集団を異にする雄間での殺戟(さつりく)を伴う集団間関係の事例が報告されている。
ニホンザルは明確な交尾期をもつが,熱帯地方に生息する高等霊長類には一定の交尾期をもたないものが多い。また,ニホンザルなどでは出産は毎年あるいは隔年であるが,チンパンジーやボノボなどでは出産間隔は約5年となる。寿命も,ニホンザルなどでは約20年といわれるが,チンパンジーやゴリラでは40年以上と推定されている。複雄複雌の構成をもつオナガザル類の集団内では,よく発達した優位劣位関係が知られているが,類人猿では食物の分配などの社会関係も見られる。これらは,集団内でのコミュニケーションの発達,成長に要する期間の長期化とも無関係ではないと考えられる。
→化石類人猿 →化石霊長類 →類人猿
執筆者:伊谷 純一郎+中務 真人
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 最新 心理学事典最新 心理学事典について 情報
Sponserd by ![]()
哺乳(ほにゅう)綱霊長目に属する動物の総称。この目Primatesは、動物界でもっとも進化を遂げたヒトや類人猿を含むが、同時に原始的な原猿類をも含み、このこと自体が霊長目の大きな特徴の一つである。その系統を追うことは、原始哺乳類からヒトまでの進化のあとをたどることに通ずる。
[伊谷純一郎]
霊長類は約7000万年前に食虫類に類似の哺乳類から分岐したといわれており、最初の化石は北アメリカ・ロッキー山脈沿いの暁新世中期の地層から出土する。フェナコレムール科Phenacolemuridaeなど3科6属が知られている。これらは始新世以降の原猿類とはつながらない原始性を残しているが、食虫類とははっきり区別でき、両目の分岐は暁新世初期にさかのぼると考えられている。暁新世の原猿は、前方に突出する大きい切歯をもち、閉じていない眼輪をもつものもあった。暁新世後期にも北アメリカとヨーロッパでさらに7属の化石が出土している。原猿類は始新世には、北アメリカ、ユーラシアに分布を広げ、アダピス科Adapidae、オモミス科Omomydaeなど5科四十数種の化石が知られ、繁栄期を迎える。これらは暁新世の原初的原猿とは異なり、より大きな脳と目を備え、鼻口部は短縮し、すでに高等霊長類に向かう進化のスタートが切られたことを物語っている。メガネザル科、そして最初の真猿アンフィピテクスAmphipithecusなどがこの時期に出現している。始新世に全盛を誇った原猿は漸新世には姿を消し、ヒトニザル上科が姿を現す。それらはエジプトのファイユームから出土した化石群で、原猿から真猿への移行を物語るアピィディウムApidiumやパラピテクスParapithecus、ショウジョウ科Pongidaeの祖先と考えられるエオロピテクスAeolopithecus、エジプトピテクスAegyptopithecusなどである。エオロピテクスはテナガザルへの分岐を示すし、エジプトピテクスは中新世の類人猿につながるものと考えられている。このほか、旧世界ザルの先祖と推定される化石も出ている。中新世と鮮新世にはふたたび多くの霊長類の化石がみられ、その多くは現生種と深いつながりをもつもので、分布もユーラシア、アフリカ、南アメリカに及ぶ。そしてヒトニザル上科の繁栄期を迎え、プロコンスルProconsul、ケニアピテクスKenyapithecus、さらに鮮新世末にはアウストラロピテクスAustralopithecus類が出現し、ヒトの時代である第四紀を迎える。
[伊谷純一郎]
霊長目は、原猿亜目Prosimiiと真猿亜目Anthropoideaに、さらに前者をメガネザル下目Tarsiiformes、ロリス下目Lorisiformes、キツネザル下目Lemuriformesの3下目に、また後者をオマキザル上科Ceboidea、オナガザル上科Cercopithecoidea、ヒトニザル(あるいはヒト)上科Hominoideaの3上科に分ける。科のレベルでは、原猿はメガネザル科Tarsiidae、ロリス科Lorisidae、キツネザル科Lemuridae、インドリ科Indriidae、アイアイ科Daubentoniidaeの5科に、真猿はオマキザル科Cebidae、マーモセット科Callithricidae、オナガザル科Cercopithecidae、ショウジョウ科Pongidae、ヒト科Hominidaeの5科、目全体で10科に分かれる。属以下のレベルについては諸説あるが、ほぼ55属約180種とみてよい。
[伊谷純一郎]
上述のように原始的な種ときわめて高等な種を含む霊長目を定義することはかならずしも容易ではないが、すべての種は次の特性を共有している。平づめ、発達した鎖骨、母指対向により物を握ることのできる手と足、閉じた眼輪(暁新世の原猿のあるものを除く)をもつ。他の哺乳類との大きな相違は、霊長類が樹上への適応を通じて独自の進化を遂げてきたということであり、上記の特徴も四肢と目に集中しているのであるが、それは樹上での活動を保証する手足と、遠近の目測を可能にする目を意味している。さらに目の変化と並行して、鼻口部の短縮、歯数の減少の傾向がみられる。原始的な哺乳類の歯式の基本型は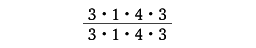
の44本であるが、すべての霊長類の歯はこれより少なく、現生種には32本を越えるものはなく、最少はアイアイの18本である。そして、大脳とくに皮質の発達、単なる歩行器官からの手の分化、産子数の減少、妊娠期間の長期化、寿命の延長、コミュニケーションの発達、社会構造の複雑化などが、系統的な段階があがるにつれて顕著に現れてくる。
[伊谷純一郎]
霊長類を対象にする学問、すなわち霊長類学primatologyは、自然人類学の基礎分野の一つに位置づけられ、人類進化史探究の科学的接近を目的とする。また、霊長類はヒトに近く発達した脳神経系をもつがゆえに、医学・薬学・神経生理学・心理学などの重要な実験動物とされる。20世紀後半に至って、野外研究・実験的研究ともに盛んになり、各国に霊長類研究所がつくられた。国際霊長類学会International Primatological Societyは隔年に開催されている。国際学術誌として『Primates』(日本)、『Folia Primatologica』(スイス)、『International Journal of Primatology』『American Journal of Primatology』(アメリカ合衆国)がある。
[伊谷純一郎]
『伊谷純一郎編『人類学講座2 霊長類』(1977・雄山閣出版)』▽『杉山幸丸著『サルの百科』(1996・データハウス)』▽『西田利貞・上原重男著『霊長類学を学ぶ人のために』(1999・世界思想社)』
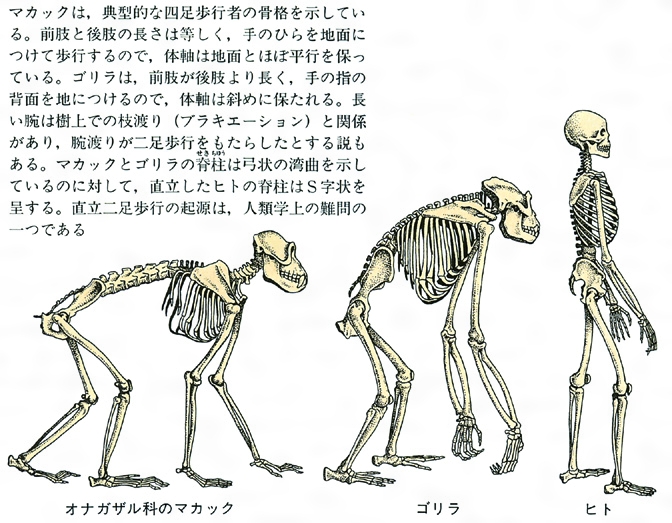
霊長類の骨格
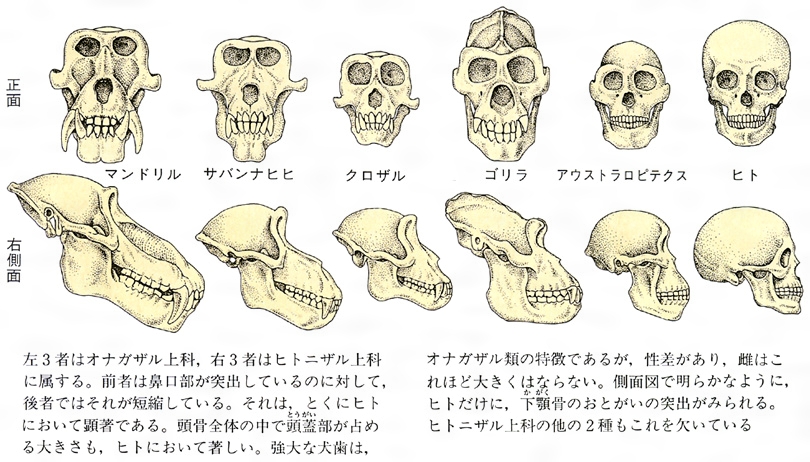
真猿類の頭骨の比較
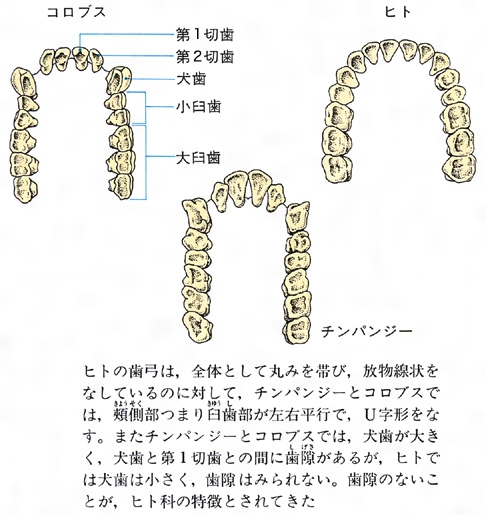
真猿類の上顎歯弓の比較
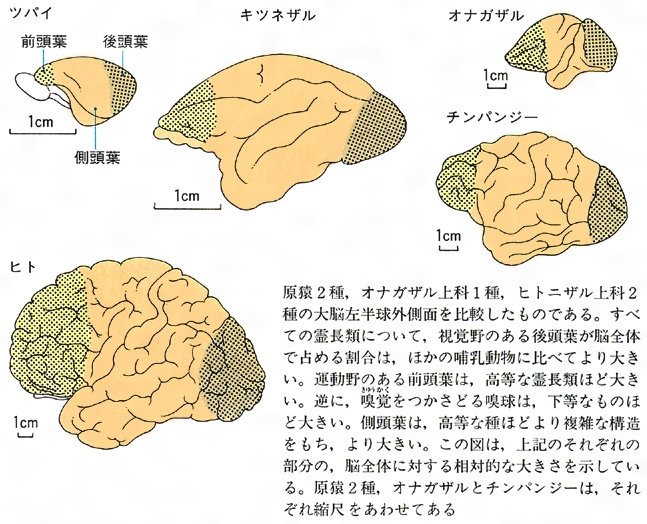
霊長類の脳の比較
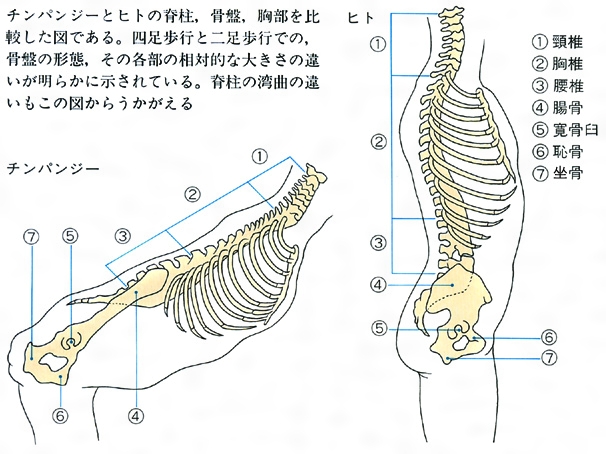
チンパンジーとヒトの躯幹
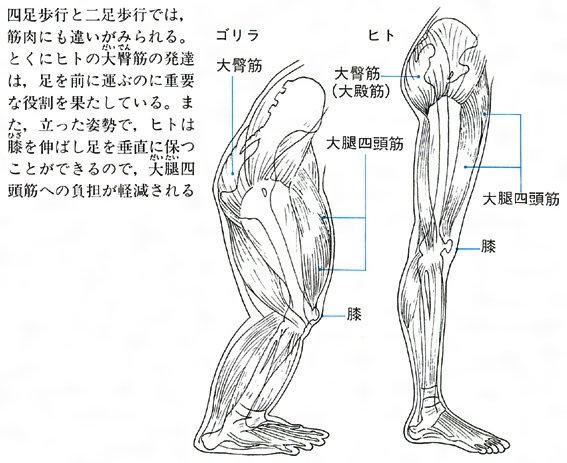
ゴリラとヒトの下肢の筋肉の比較
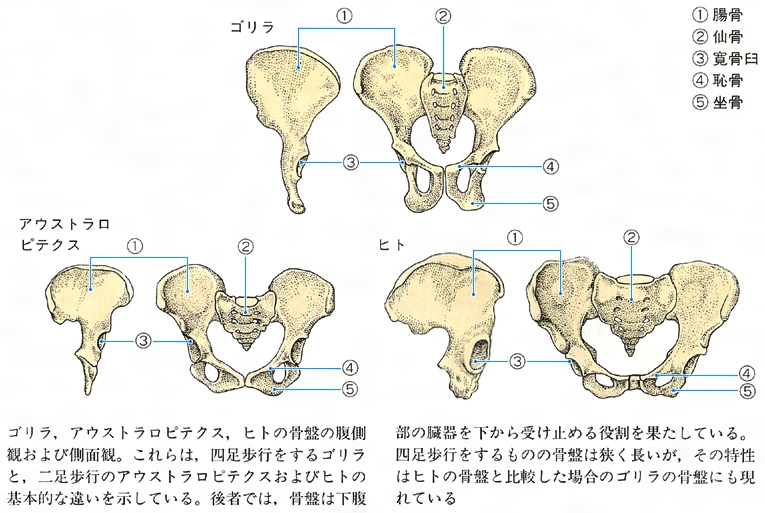
霊長類の腰(骨盤)
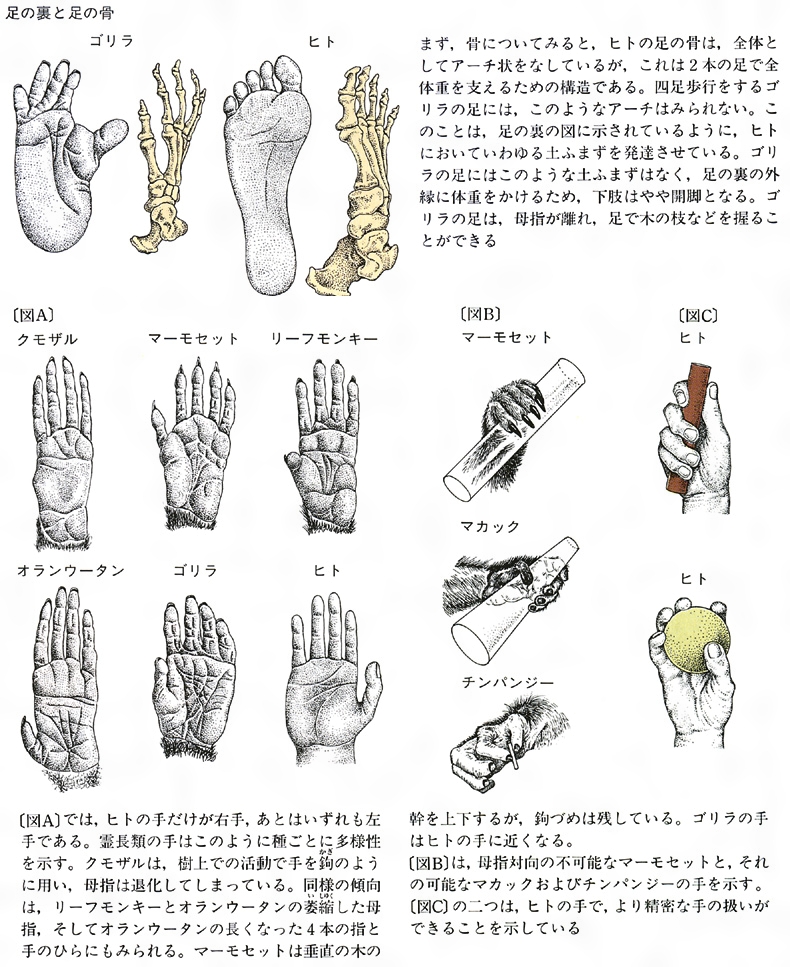
霊長類の足と手
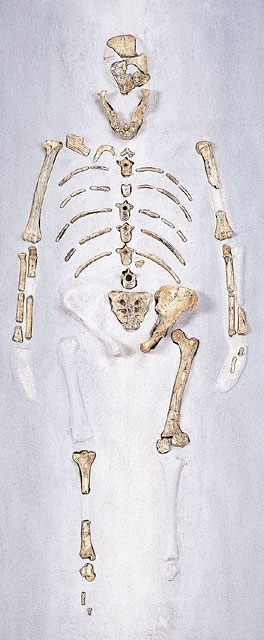
アウストラロピテクスの化石骨

サルのおもな種類(オナガザル類)〔標本…

サルのおもな種類(オマキザル類)〔標本…

サルのおもな種類(原猿類)〔標本画〕

サルのおもな種類(ヒトニザル類)〔標本…

アカゲザル

オランウータン

コビトガラゴ

シロガオサキ

チンパンジー

テイチゴリラ(ローランドゴリラ)

ドグエラヒヒ

ニホンザル

パタスモンキー

ピグミーマーモセット

フクロテナガザル

ボリビアリスザル

ワオキツネザル

ワタボウシタマリン
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
学◆Primates 英◆primates
哺乳綱獣亜綱正獣下綱の一目。かつてプレシアダピス類(Plesiadapiformes)も霊長類としていたが,現在では,はずす傾向にある。霊長類には,アダピス類(Adapiformes)・キツネザル類(Lemuriformes)・オモミス類(Omomyiforrnes)・メガネザル類(Tarsiiformes)・広鼻類(Platyrrhini)・狭鼻類(Catarrhini)が含まれる。系統的に,アダピス類・キツネザル類,そしてオモミス類・メガネザル類・広鼻類・狭鼻類がそれぞれ単系統と考えられる。前者を曲鼻類(Stepsirhini),後者を直鼻類(Haplorhini)とすることがある。霊長類は樹上が基本的な生活の場。第一指が対向し,握ることができる手・足,そしてよく発達した視覚が特徴。大脳は,体重と比較すると海生哺乳類を除いて最大。霊長類の骨学的な特徴は,鼓室を包む骨が耳周骨(岩骨)であり,一般的な哺乳類の内鼓室骨と異なる。
執筆者:亀井 節夫・名取 真人
出典 平凡社「最新 地学事典」最新 地学事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
Sponserd by ![]()
…地質時代に生息し,化石として残されている霊長類の総称。中生代も終わる頃になると,大陸の地形に大きな変化がみられ,ロッキー山系の隆起もこの頃に生じた。…
…ヒトにもっとも近縁な動物で,ヒトとともに哺乳綱霊長目をなす。もともとニホンザルを指すことばであったが,現在ではヒト以外の霊長類の総称として用いられ,狭義には,真猿類のオマキザル科とオナガザル科の種を指す。英語では,尾の長いサルをmonkey,尾のないサルをape,原猿類をlemur,またはprosimianといっている。…
…人種を考える場合には人類の系統的・発生的側面に,また民族を考える場合には生活面を重視しその地縁的・文化史的側面に着目することになろう。
【社会的文化的特徴とその進化】
かつてヒトを定義するための多くの特性があげられたが,近年の霊長類を対象とした研究によってそれらの中のいくつかはヒトの定義に耐えうるものではないことが明らかにされるにいたった。ヒトは道具を使用し道具を作る動物であるといわれたが,野生チンパンジーについての多くの観察がそれを無効にしたというのもその一例であろう。…
※「霊長類」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
春になって暖かくなりかけた頃、急に寒さが戻って、地面などがまた凍りつく。《 季語・春 》[初出の実例]「七瀬御秡 同晦日也。〈略〉雪汁いてかへる」(出典:俳諧・誹諧初学抄(1641)初春)...