改訂新版 世界大百科事典 「遺伝的組換え」の意味・わかりやすい解説
遺伝的組換え (いでんてきくみかえ)
genetic recombination
親から子に,または個体から個体に遺伝子が伝達される時,遺伝子の組合せが変わること。例えば,G.J.メンデルは,エンドウで丸くて黄色の豆をつける植物としわが寄った緑色の豆をつける植物との交配の結果,雑種2代目に親と同じ形質の植物だけではなく,丸くて緑色の豆をつける植物やしわが寄った黄色の豆をつける植物が生じることを見いだした(1865)。つまり,豆の形と豆の色という形質,ひいてはそれぞれの形質を決定している遺伝子の組合せが変わっていることを発見した。これを基にして,いわゆるメンデルの〈独立分離の法則〉が提唱された。
連鎖linkage
〈独立分離の法則〉の例外を見いだし,連鎖という概念を最初に出したのはスタートバントA.H.Sturtevant(1913)である。この現象は,複数の遺伝子が〈独立分離の法則〉に従わず組として親から子に(または個体から個体に)伝わるもので,その程度は遺伝子の組合せによって異なるが,通常は50%を越えることはない。そして,連鎖の頻度を基準にして,遺伝子を直鎖状に配列できることも明らかになった。これは,飼育が容易で,子孫の数が大きく,しかも変異個体が多く得られるショウジョウバエDrosophilaを使ったT.H.モーガンのグループの研究の成果である。その後,さらに多くの遺伝子についての連鎖の解析から,詳細な連鎖地図が作られた。ショウジョウバエでは遺伝子が4組の連鎖群に分かれるが,これは顕微鏡で観察できる相同染色体対の数と一致した。他の生物でも連鎖群の数と相同染色体対の数とが一致することが明らかになるにつれ,連鎖群が一つの染色体上にある遺伝子群にほかならないことが明らかになり,連鎖地図は染色体上での遺伝子の並び方を示していると考えられるようになった。現在では,連鎖地図と染色体地図とは同じ意味に使われている。
メンデルが実験に使った7対の形質に対応する遺伝子がそれぞれ別の染色体上にあることが後になって明らかになった。つまり,メンデルの〈独立分離の法則〉は,減数分裂(細胞分裂)の時に対合していた相同染色体どうしはかならず分離するが,異なる染色体の分離は互いに独立であるということで説明できる。ところで,エンドウには7対しか相同染色体がないのだから,もしもメンデルが8番目の形質を交配実験に加えていたなら連鎖を発見していたかもしれない。なぜなら,8番目の遺伝子は他の七つの遺伝子のどれかと同じ染色体上にあるはずであり,その場合,なんらかの程度の連鎖が見られると考えられるからである。逆に言えば,メンデルは連鎖のない遺伝子の組合せを選んで実験を行い,遺伝子の粒子性を明確に示そうと意図したとも考えられなくはない。
交叉crossing-over
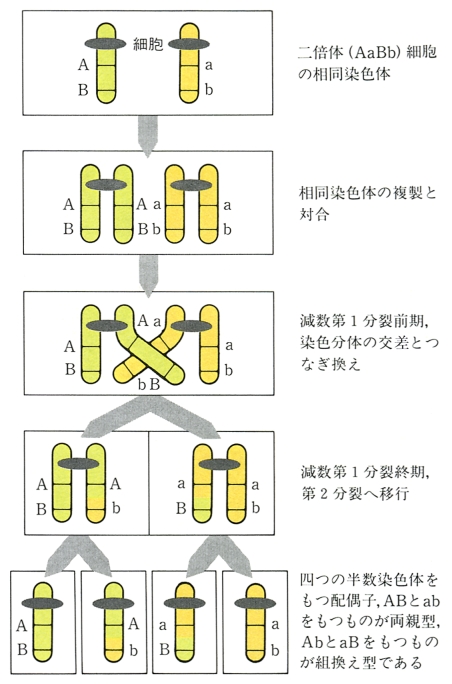
染色体の独立分離による遺伝的組換えを染色体間組換えという。これに対して,同じ連鎖群つまり同じ染色体上の遺伝子間の組換えを染色体内組換えという。特にその機構を問題にする場合,染色体内組換えに対して交叉(または乗換え)という語を使うことがある。染色体内組換えの機構としては,高等生物の減数第1分裂前期から後期にかけてみられるキアズマとの関連が考えられた。かつてはキアズマの点で染色分体の切断と融合(つまり乗換え)が起こると考えられたが,現在ではキアズマは乗換えの結果ではあるかもしれないが,染色体の乗換えそのものを示しているのではないと考えられている。しかし,キアズマと組換えとの関係は完全に解明されたわけではなく,(1)相同染色体が対合する機構,(2)姉妹染色体間の相互作用,(3)主要なDNAの複製よりも遅れて起こるDNA合成の意義,などと関連して今後解決されるべき課題である。
染色体内組換えを大腸菌に感染するT4ファージを使ってきわめて詳細に調べたのはベンザーS.Benzer(1957)である。ファージはショウジョウバエよりもはるかに早く増殖するし,飼育(培養)も容易である。この研究によって,組換えが遺伝子間だけでなく,遺伝子内でも起こることが明らかになった。これは,遺伝物質の中で遺伝情報を担う部分とそうでない部分とが基本的に違っていないことを示した点で重要な研究である(遺伝物質であるDNAが直鎖状であり,遺伝子である部分もそうでない部分もDNAとしては基本的に差がないことと合致する)。この研究以後,細菌や高等生物においても遺伝子内組換えを利用して,遺伝子の内部構造の解析が行われるようになった。
遺伝的組換えの分子機構
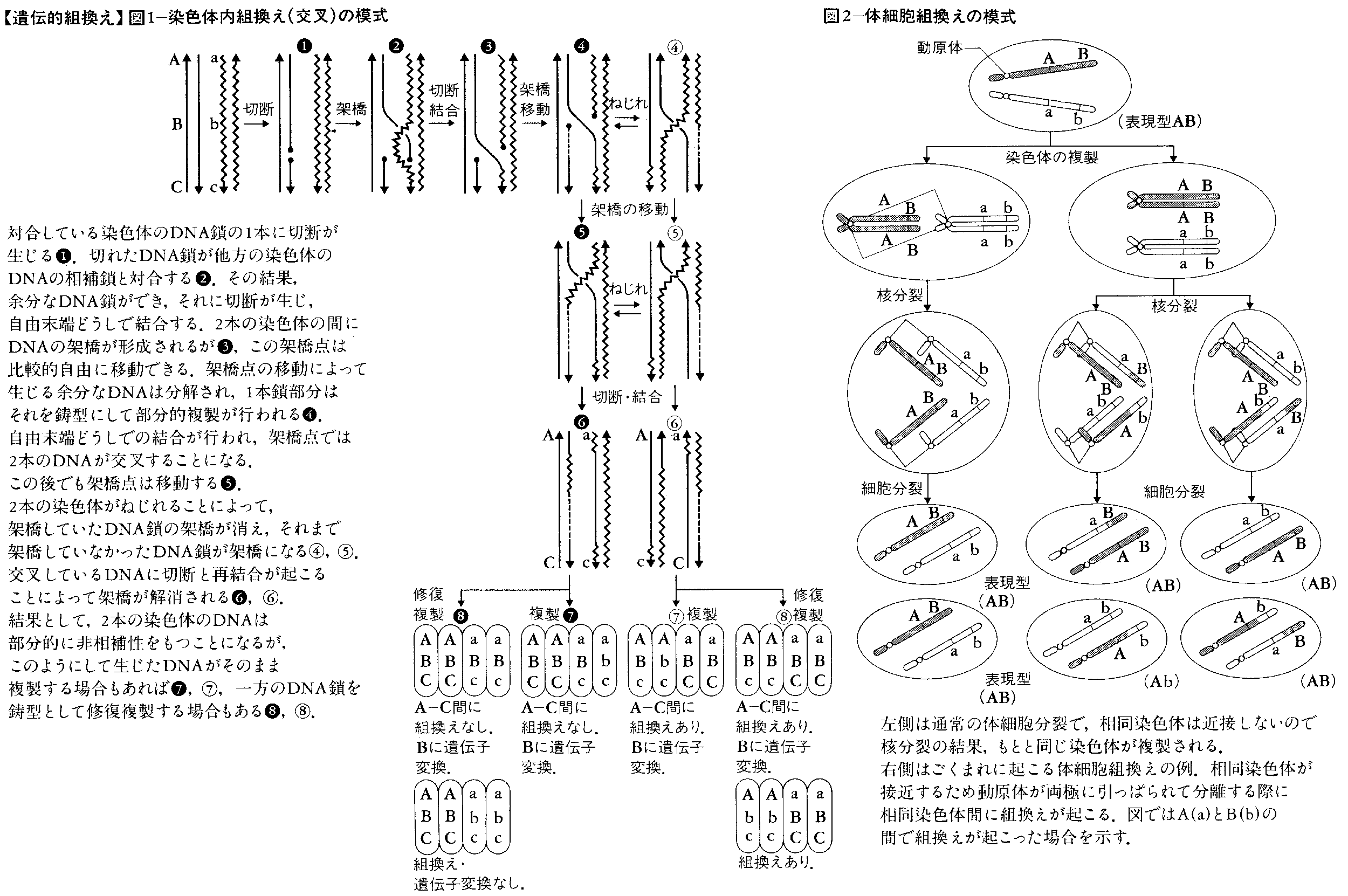
メセルソンF.MeselsonとワイグルJ.Weigleは,目で見て区別できる変異をもつλファージを用い,さらに,非放射性同位体(12Cに対して13C,14Nに対して15N)によってDNAの密度を変えられることを利用して,遺伝的組換え(この場合は染色体内組換えに相当する)がDNAの切断と再結合によって起こることを示した(1961)。メセルソンとラディングC.M.Raddingはこの研究結果を基盤にして,現在のところ最も広く受け入れられている染色体内組換えの分子機構(図1)を提唱した(1975)。メセルソンとラディングの模式は,染色体内組換えだけでなく,遺伝子変換gene conversionをも統一的に説明できるので注目されている。なお遺伝子内組換えや遺伝子変換の遺伝学的研究には,減数分裂の産物を組として分析できる子囊菌類(アカパンカビやパン酵母)が適している。これらの生物を使った分子遺伝学と古典遺伝学とを統合する方向での研究が今後重要になろう。図1にはメセルソンとラディングの模式に基づいて期待される4分子の遺伝子型を付け加えてある。
染色体内組換えは減数分裂の時だけでなく体細胞分裂の時にも低い頻度で起こる。これを体細胞組換えというが,機構としては減数分裂時の組換えと同じであると考えられている。つまり,体細胞分裂においては相同染色体は対合しないのであるが,まれに近接することがあり,その時に組換えが起こる(図2)のであろう。体細胞組換えの結果は体細胞分裂分離として観察される。なお現在では,染色体の特殊な染色によって姉妹染色分体間にも,切断-再結合による組換え(交叉)が起こっていることが明らかになっている。
遺伝的組換えの生物学的意味
生物体の表現型は形質という単位に分けられる。そして,各形質は遺伝子と環境要因との相互作用で決定される。つまり,遺伝的組換えは,遺伝子の組合せを多様にすることによって生物の表現型を多様にする機構であるとも言える(遺伝子そのものを変える突然変異も表現型を多様化する機構である)。世代交代をする生物(真核生物)では主として親から子に遺伝子が伝えられる時に,遺伝的組換えによって遺伝子の組合せが変化する。一方,世代交代のない生物(主として原核生物)では,個体から個体に遺伝子を伝達することによって遺伝子の組合せが変わる。後者の場合,伝達の方法としては,接合,形質導入,形質転換が知られている。生物種によってどの伝達方法を主にしているかは異なるが,いずれもなんらかの形でDNAの組換えが関与している。メセルソンとラディングの模式における組換えでは,組換えが起こるDNA間には塩基配列の相同性が想定されているが,塩基配列の相同性がなくても起こる組換え(illegitimate recombination)が細菌やファージやプラスミドや挿入配列などでみられることが明らかになっている。原核生物におけるこれらの遺伝子の伝達方法が,自然界でどの程度の重要性をもつかはまだ不明である。しかし実験室内での遺伝解析にはきわめて重要で,分子遺伝学の研究に,原核生物である細菌,特にその中でも大腸菌がよく使われ,生物学的に重要なさまざまな発見が行われているのは,上に述べた遺伝子の伝達方法を利用して遺伝子を比較的自由に操ることができるからである。近年,急速に実用化が進められている遺伝子工学というのも,組換えDNA実験という別名が示すように,人為的に試験管内でDNAの組換え(遺伝子組換え)を行う手法が基礎になっている。
組換え研究の将来
ショウジョウバエの精母細胞やクロユリの花粉母細胞では染色体内組換えがない(キアズマもみられない)ことが知られている。また細菌やパン酵母では,染色体内組換えが起こらない,ないしはきわめて頻度が低い変異株が得られており,その性質が遺伝学的および生化学的に調べられている。これらの研究によって,染色体内組換えに関与するタンパク質(酵素)も明らかになりつつあり,DNA分解酵素やDNA合成酵素などの関与も解明されつつある。今後は交叉の機構が分子のレベルでより詳細に明らかになるにつれ,組換え機構の進化も明らかになり,組換えが生物の進化に及ぼした効果も物質レベルで理解されるようになることが期待される。
遺伝子伝達における染色体内組換えとは別に,細胞分化における染色体内(この場合は単一染色体内)組換えの関与が近年実験的に示されるようになった。利根川進らはγ-グロブリン遺伝子の研究から,細胞分化と遺伝子の再編成とが対応していることを示し,その再編成には組換えによる欠失が起因しているという考えを提唱した(1976)。この場合は,遺伝子の調節部分の付着する構造遺伝子が欠失によって変わるのであるが,特定の遺伝子の欠失が細胞分化につながることもありうる。この種の研究は細胞分化を分子レベルで理解するという面で今後の進展が期待される。
執筆者:小野 文一郎
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponsored by ![]()