デジタル大辞泉 「DNA」の意味・読み・例文・類語
ディー‐エヌ‐エー【DNA】[deoxyribonucleic acid]
1 ⇒デオキシリボ核酸
2 俗に、遺伝子のこと。また、先祖から子孫へ連綿と伝わるもの。「父の
[類語]細胞
ディー‐エヌ‐エー【DNA】[Defense Nuclear Agency]
Sponserd by ![]()
Sponserd by ![]()
更新日:
出典 共同通信社 共同通信ニュース用語解説共同通信ニュース用語解説について 情報
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
デオキシリボ核酸の通称。デオキシリボ核酸の英語表記DeoxyriboNucleic Acidの頭文字を並べてDNAとした。
2-デオキシリボヌクレオチド(2'deoxyribonucleotide)が、3'-5'間のフォスフォジエステル結合で、直鎖状に重合した高分子化合物。核酸には、DNAとRNA(リボ核酸RiboNucleic Acid)とが含まれる。核酸は、タンパク質、糖質、脂質などとともに生物体を構成している高分子化合物の一つである。ほとんどすべての生物の遺伝子は、DNAでつくられているが、ウイルスのなかにはRNAを遺伝子としているものが若干存在する。
[菊池韶彦]
1860年代の後半、スイスのバーゼルに在任していた有機化学者ミーシャーFriedrich Miescher(1844―1895)は、細胞核内にある化合物を重要と考え、病院から、膿(うみ)で汚れたガーゼや包帯を集め、白血球(とくに核が大きい)の死骸である膿から、新しい強酸性の化合物を発見し、これにヌクレインnucleinと名前をつけ、1867年に発表した。ヌクレインは、C(炭素)、H(水素)、N(窒素)、O(酸素)の元素を含むほかに大量のP(リン)を含んでいた。同様の物質はサケ(マス)の精巣(精子もまた核の占める割合が大きい)からも検出することができた。サケは、ライン川を遡(さかのぼ)ってくる間に、筋肉が減り、精子が増えていることから、ミーシャーは筋肉のタンパク質が変化したものと考え、おそらく核にある特有のタンパク質を意図して「Nuclein」と名づけたと考えられる。1889年に、ドイツの有機化学者アルトマンRichard Altmann(1852―1900)は、これはタンパク質とは異なる新奇な高分子化合物だとし、新たに「核酸Nucleic Acid」と命名し直した。
[菊池韶彦]
ほかの生体高分子化合物(タンパク質、糖質、脂質など)に比べて、核酸の構成成分はきわめて単純であり、4種の2'-デオキシリボヌクレオチドすなわち、アデニル酸、グアニル酸、シチジル酸、チミジル酸が重合したものである。4種の2'-デオキシリボヌクレオチドは、リン酸基1分子と2'-デオキシリボースにそれぞれ異なる塩基、アデニン(A)、グアニン(G)、シトシン(C)、チミン(T)のいずれかが1分子ついている。DNAは2'-デオキシリボースの3'-OHと5'-OH間がリン酸によりフォスフォジエステル結合で直鎖状に重合している。アデニンとグアニンはプリン環をもつ塩基であり、シトシンとチミンはピリミジン環をもつ塩基である。こうした構成の単純さから、生物学的な機能の解析は遅れ、単に核内のタンパク質の支持体と考えられていた。とくに、P・A・T・レビン(フェーブス・レビーン)が1912年に発表した四つのヌクレオチドを単位とした構造、テトラヌクレオチドTetranucleotide仮説が有力となり、生物学的な機能の研究は遅れることになった。
[菊池韶彦]
1928年にグリフィスFrederick Griffith(1879―1941)は連鎖状球菌(肺炎双球菌)の形質(遺伝的な性質)を変えることのできる物質があることを発見し、これを形質転換物質transforming principleとよんだ。1944年にO・T・エーブリー(アベリー)、マクラウドColin Munro MacLeod(1909―1972)、マッカーティーMaclyn MacCarty(1911―2005)の3人により、細菌に精製したDNAを直接混ぜると、細菌の形質が変わること、さらにこの形質転換の活性は、タンパク分解酵素によっては影響されず、DNA分解酵素で処理すると、完全に失われることから、形質転換を行っている物質はDNAであろうと示唆された。しかし、当時この発見も広くは受け入れられずに、最終的に遺伝子はDNAであるということが一般的に認められるようになったのは、1952年に発表されたA・D・ハーシェイとM・チェイスによる実験である。彼らは、遺伝子がDNAかタンパク質かという議論に決着をつけるために、大腸菌に感染するT2バクテリオファージ(ウイルスの一種、T2ファージと略称する)を使用した。T2ファージは、タンパク質とDNAのみから構成されており、大腸菌に感染させると、十数分のうちに100倍にも増殖し、子どものファージをつくる。このファージのタンパク質を放射性同位元素S35、DNAをP32で標識し、大腸菌に感染後それぞれの標識された化合物がどうなるかを測定した。ファージが吸着した直後の大腸菌をミキサーにかけたのち、遠心分離器で大腸菌を集めると、S35の標識の80%は大腸菌とは共存せず、逆にP32の80%は大腸菌にとどまっており、この集めた菌体を新しい培地で培養すると子どものT2ファージが生産され、そこには30%のP32が回収され、S35は1%以下であった。この結果から、ファージをつくるための遺伝子はDNAであろうと推論した。この実験から、子どものファージをつくっている遺伝情報はタンパク質ではなくDNAにあるという考え方が広まるようになった。
[菊池韶彦]
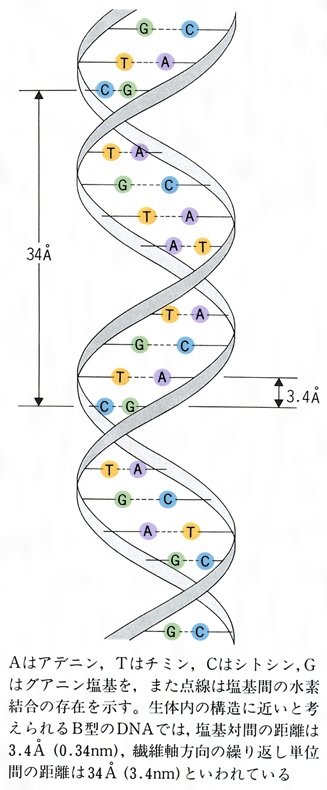
DNAが遺伝子であることを示すには、どのようにコピー(複製)がつくられるかということを示す必要がある。1951年にL・C・ポーリングはX線結晶解析の結果をもとにタンパク質の基本的な構造として、α(アルファ)-へリックスを提案し、これにより、タンパク質の機能が理解できることを示した。おそらくDNAも遺伝物質としての機能のためには何らかの高次構造がかかわるものと考え、J・D・ワトソンとF・H・C・クリック、それにDNAのX線結晶解析を進めていたM・H・F・ウィルキンズ(とR・E・フランクリン)により、1953年にDNAの二重螺旋モデル(にじゅうらせんもでる)が発表された。このモデルは、X線結晶解析から、(1)DNAの構造には周期性のある螺旋構造がある、(2)螺旋の直径は2ナノメートル(20オングストローム)で長軸方向に互いに逆向きになる2本の螺旋が並行している、(3)塩基は0.34ナノメートル(3.4オングストローム)の間隔にある、(4)螺旋の周期は3.4ナノメートル(34オングストローム)で塩基10個に相当する、と推定した。また、DNAの化学的性質から、リン酸基は分子の外側にあり、2本の螺旋は塩基間の水素結合により安定に維持されていると考えられた。螺旋の直径が2ナノメートルになることから、ピリミジンとプリンを組み合わせるとこの大きさになり、AとT、GとCの相補的な塩基対により最適な水素結合がつくられることが示された。ちなみに、E・シャルガフはさまざまなDNAの塩基組成を解析し、その結果からDNAのアデニン(A)とチミン(T)、グアニン(G)とシトシン(C)の量はほぼ等しいとする「シャルガフの通則」を提唱していたが、二重螺旋モデルは「シャルガフの通則」をも説明していた。
遺伝子としての、コピーされるためのもっとも基本的な方法も、このDNAの二重螺旋モデルで説明される。2本の螺旋が二つに分かれ、それぞれに対して相補的な塩基対をつくってDNA合成が起これば、もとのDNAとまったく同一のものがつくられることになるからである。この複製機構が正しいことは、1958年にメセルソンMatthew Stanley Meselson(1930― )とスタールFranklin William Stahl(1929― )による実験で証明され、半保存的なDNA複製のモデルが提案された。
[菊池韶彦]
DNAが遺伝子としての役割を果たすためには、正確にコピー(複製)されなければならない。1956年にA・コーンバーグは大腸菌の抽出液から、DNAを鋳型として同じ配列をもったDNA複製をする酵素DNAポリメラーゼを精製した。この酵素は、(1)DNAを鋳型として相補性をもったDNA鎖を合成する、(2)4種の2-デオキシリボヌクレオシド三リン酸(dATP、dGTP、dCTP、TTP)を基質とし、ピロリン酸を遊離する、(3)ATPとMg2+が必須である、(4)DNA合成開始には3'-OHをもったプライマーprimer(最初のヌクレオチド合成が起こるために3'-OHを提供する短いヌクレオチド)が必要である、(5)合成の方向は5'から3'の方向にしか起こらない、という特徴をもっていた。このことから、1967年(昭和42)には分子生物学者の岡崎令治(おかざきれいじ)(1930―1975)らは、DNA複製は不連続にしか起こらず、複製中間体として岡崎フラグメント(岡崎令治が発見したDNA断片)を経由するというモデルを発表した。さらに1975年に岡崎令治らは、プライマーは短いRNAであることを発見した。なお、ウイルスやバクテリオファージのDNA複製では、DNA複製のプライマーとしてはタンパク質中のアミノ酸にあるヒドロキシ基が使われることもある。
[菊池韶彦]
遺伝子であるDNAは、遺伝情報を子孫に伝えるだけでなく、自分自身の細胞を構成するすべてのRNAとタンパク質をつくりだす使命を担っている。とくにF・ジャコブとJ・L・モノーは、1960年に発表したオペロン説(遺伝子が発現してタンパク質の合成が調節される仕組みを統一的に示した)の当然の帰結として、タンパク質の合成にはDNAの情報をコピー(複製)した寿命の短いRNAが中間体として存在することを予測し、この中間体をメッセンジャーRNA(mRNA)と命名した。こうしてDNAの遺伝情報は、DNAからRNAに転写され、タンパク質に翻訳されるというセントラルドグマcentral dogmaがクリックにより提唱された。
[菊池韶彦]
細胞内のDNAの大きさは、生物種によってさまざまであるが、独立して生活をしている最小のマイコプラズマでも、そのDNAは5.8×107塩基対もある。これは、ワトソン-クリックによるB型のDNAの長さは直鎖状として20ミリメートルになり、菌体がせいぜい1マイクロメートル程度であることから考えると、非常に長い。
一方、真核生物では、ヒトの場合、3×109塩基対で、全部つなぎ合わせると約1メートルに達する。核の大きさはせいぜい10マイクロメートルで、DNAはこのなかに23本の染色体に分かれそれぞれが2本ずつ23対詰め込まれている。その長さを小さくまとめるため、DNAは非常に規則正しく折りたたまれている。もっとも基本的な最小単位はヌクレオソームで、これは4種のヒストン(H2A、H2B、H3、H4)が、それぞれ二つずつ8量体となり、それに200塩基対のDNAが2巻きに巻きついている。ヌクレオソームは、さらにコイル状に折りたたまれ、染色体を構成している。細胞分裂期になると、染色体の凝縮の度合いはさらに増し、顕微鏡下で特徴的な形態が観察できる分裂中期染色体metaphase chromosomeとなり、それぞれ二つの娘細胞に分配される。
[菊池韶彦]
ワトソンとクリックが示したDNAの構造はB型とよばれる構造で、ほかにもA型という若干異なる右巻きの螺旋構造の存在も明らかにされている。このA型の構造は、RNAが二重螺旋をつくるときにとる典型的な構造である。1979年にリッチAlexander Rich(1924―2015)らは、DNAが左巻きの構造をとる可能性に注目した。この構造はZ型とよばれているが、細胞内での存在や、その役割は明らかにされていない。
多くの原核生物(細菌、古細菌)の染色体DNAやプラスミド、ウイルスのDNAは直鎖状ではなく、その両端が共有結合をした環状構造をとっている。1963年にワイルRoger WeilとアメリカのビノグラドJerome Vinograd(1913―1976)が細胞から抽出したポリオーマウイルスのDNAは環状で、DNAの二重螺旋がさらに螺旋構造をとった超螺旋構造をしていることを示唆した。1971年にはワンJames C. Wang(1936― )により超螺旋構造を解消するDNAトポアイソメレース(DNAtopoisomerase)、1976年にはゲラートMartin Gellert(1929― )により、超螺旋を導入するDNAジャイレース(DNAgyrase)が発見された。これらの酵素は、真核生物、原核生物を問わず、広く分布しており、DNAのコピー(複製)、転写に際しては必須であるため、これを標的とした抗菌剤や抗癌剤(こうがんざい)が開発されている。
[菊池韶彦]
DNAは遺伝子の本体であることから、細胞に直接DNAを加えることにより、遺伝情報を変える試みが多くなされてきた。DNAによる連鎖状球菌の形質転換法の発見以来、さまざまな細胞に若干の処理を施して(たとえば、細胞膜の透過性を高めたり、直接穴をあけたりする)DNAを取り込ませて、細胞の遺伝情報を変える操作(遺伝子工学)が可能になっている。しかし、通常の食事の一部としてDNAを摂取しても、DNAが個体に取り込まれて、遺伝情報にかかわるという報告はない。
核酸は生物にとって重要な成分であるが、ほかの栄養素、タンパク質、糖質、脂質のように、食料として摂取しなければならないことはない。生物にとっては、核酸は生命の維持にとって重要だからこそ、ほかの栄養素(アミノ酸や糖)から核酸の構成成分を自ら合成できるようになっており、核酸欠乏症はきわめて稀(まれ)である。ましてや、核酸を食事として摂取したからといって健康状態や能力が改善することは考えられず、逆に、多量の核酸を摂取すると、その成分であるプリンの分解産物として尿酸が体内に蓄積して痛風(つうふう)の原因となることも指摘されている。
[菊池韶彦]
『菊池韶彦著『DNAの冒険――二重らせんから超らせんへ』(1993・岩波書店)』▽『大島泰郎他編、今本文男著『未来の生物科学シリーズ35 DNAはどのように折れ畳まれて働くか』(1996・共立出版)』▽『C・R・カラダイン、ホレース・R・ドゥルー著、ワット富士子・西村善文訳『なぜ遺伝子はらせんを巻くのか――DNAの立体構造と機能』(1996・共立出版)』▽『渡辺政隆著『DNAの謎に挑む――遺伝子探求の一世紀』(1998・朝日新聞社)』▽『ニコラス・ウェイド編、中村桂子監修、翻訳工房ことだま訳『DNAのらせんはなぜ絡まらないのか』(2000・翔泳社)』▽『ショーン・B・キャロル、ジェニファー・K・グルニエ著、上野直人・野地澄晴監訳『DNAから解き明かされる形づくりと進化の不思議』(2003・羊土社)』▽『フィリップ・R・レイリー著、高野利也訳『リンカーンのDNAと遺伝学の冒険』1~2(2003・岩波書店)』▽『石井信一著『DNAとタンパク質――生物の特異性を決める分子たち』(2006・裳華房)』▽『J・D・ワトソン他著、中村桂子他訳『遺伝子の分子生物学』第5版(2006・東京電機大学出版局)』▽『B・ルーウィン著、菊池韶彦他訳『エッセンシャル遺伝子』(2007・東京化学同人)』▽『ジェームズ・D・ワトソン他著、松橋通生・山田正夫・兵頭昌雄・鮎沢大監訳『ワトソン組換えDNAの分子生物学――遺伝子とゲノム』第3版(2009・丸善)』▽『L・ハートウェル他著、菊池韶彦監訳『ハートウェル遺伝学』第3版(2010・MEDSI)』▽『柳田充弘著『DNA学のすすめ――躍動する生命の二重らせん』』▽『宮田隆著『分子進化学への招待――DNAに秘められた生物の歴史』』▽『工藤光子・中村桂子著『DVD&図解 見てわかるDNAのしくみ』(以上講談社ブルーバックス)』▽『三浦謹一郎著『DNAと遺伝情報』(岩波新書)』▽『木村陽二郎著『原典による生命科学入門』(講談社学術文庫)』▽『J・D・ワトソン著、江上不二夫・中村桂子訳『二重らせん』(講談社文庫)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
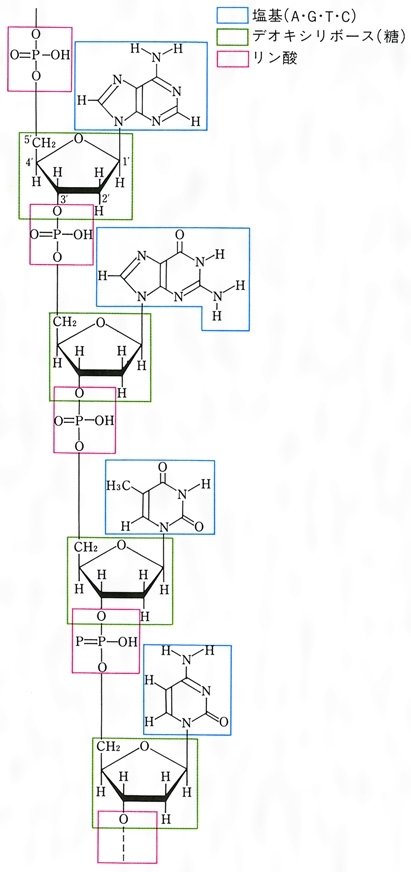
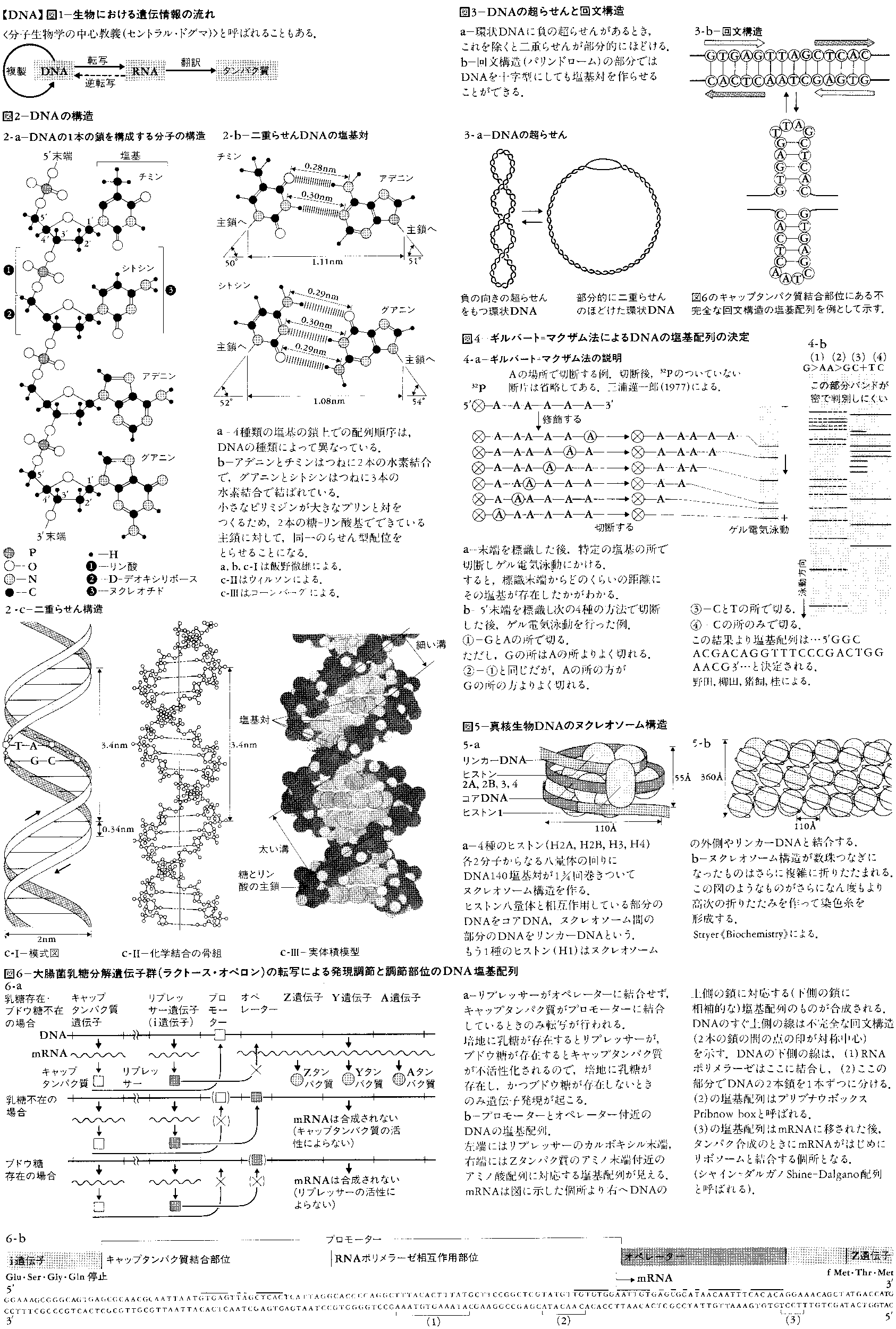
デオキシリボ核酸deoxyribonucleic acidの略称。核酸のうち糖成分がD-デオキシリボースであるもので,遺伝子の本体をなす。教育や体験によらずに親から子へと自動的に伝わる性質を遺伝形質という。この遺伝形質のもととなり,またそれを親から子へと伝える物質の単位を遺伝子と呼ぶのであるが,遺伝子とは,実は細長い糸のような物質DNAにほかならない。DNAはどの生物のどの細胞にも,その生物の全遺伝情報分だけある。細胞が分裂する前にDNAは複製され,分裂後の二つの娘細胞は同じDNAを含む。また,DNAの遺伝情報は,メッセンジャーRNA(mRNAと略記)を通じて,タンパク質へと移されて発現する(図1)。生物の体を形づくっているのも,生物が生きてゆくための働きを担っているのも主としてタンパク質であるから,個々の生物の性質はどのようなタンパク質ができるかで決まる。そういう意味で,DNAは生物の設計図ということもできる。
DNAの鎖は,D-デオキシリボースという糖とリン酸が交互につながってできている。そして,おのおのの糖には,必ず一つの塩基が結合している(図2-a)。リン酸・糖・塩基が一つずつ結合したものをヌクレオチドというので,この鎖はポリヌクレオチド鎖と呼ばれることもある。塩基にはアデニン(A),グアニン(G),シトシン(C),チミン(T)の4種があり,DNAの鎖の上での塩基の並び方が遺伝情報を与える。つまり,生命の記号は4文字のアルファベットで書かれているのである。DNAの鎖には向きがあり,鎖の両端はふつう糖の炭素原子の番号づけに従って,5′末端と3′末端と呼ばれる。大多数のDNA分子では,2本の鎖が塩基を内側にして互いに逆向きに並び,10塩基で1回転する右巻きらせんを作っている(図2-b)。この二重らせん構造は,発見者の名により,ワトソン=クリックWatson-Crickのモデルともいわれる。2本の鎖の塩基間は水素結合という弱い結合で結ばれているが,この塩基対は必ずAとTまたはGとCという組合せのどちらかである。このことを,2本の鎖の間では塩基配列が相補的になっているという。このため,どの生物のDNAをとってみても,AとT,GとCの含量はつねに等しく(シャルガフの法則),変化するのは全塩基に対するG+C(またはA+T)の含量のみである。DNAの二重らせんモデルは,DNAの機能である情報の貯蔵,複製,転写(mRNAの合成),突然変異,DNA修復がどのようにして行われるかを,その構造自体が暗示している。そのため,このモデルが1953年に発表されるやいなや,分子生物学の爆発的発展を引き起こした。
化学物質としてのDNAは,リン酸基をもつため強い酸性の物質であり,通常は塩の形で存在する。また,そのため塩基性タンパク質,2価金属イオン,ポリアミン類などを強く結合する。RNAと異なりアルカリに対しては比較的強いが,低いpHではAやGが糖から外れやすい。DNAの定量は,デオキシリボース部分をインドールまたはジフェニルアミンと反応させ,比色法により行う。加水分解してリンの定量をしたり,塩基による260nm付近の紫外吸収で定量することもできる。精製したDNAは,水溶液中では太さ約20Åの屈曲性のある鎖であり,ランダムに曲がって熱運動をしている。糸も短い部分をとると棒と見なせるように,DNAも,長さ1000Å以下では1本の棒のようにふるまう。DNAの浮遊密度はおよそ1.7g/mlであり,G+C含量が高いほど密度は大きい。したがって,塩化セシウムなどの濃厚溶液の密度こう配中で遠心分離すると,G+C含量によりDNAを分別することができる。
DNAを熱していくと,ある温度で急に塩基間の水素結合が切れ,2本鎖は分かれて1本ずつになる。これをDNAが融解するという。融解温度は溶媒のpHやイオン強度のほか,DNAの塩基組成にも依存し,G+C含量が多いほど融解温度は高い。また,融解により260nmの紫外吸収は約4割増大する。熱して分かれたDNAを冷却し,融解温度よりわずかに低い温度に長時間保つと,DNAは相補的な鎖を見つけて再び2本鎖を形成する。この反応は,DNAと相補的なRNAとの間や,完全に相補的ではないがそれに近いDNA間でも起こる。したがって,この反応により,mRNAを用いてその遺伝子を捕らえたり,異なるDNA間の塩基配列の類似性を調べたりすることができる。完全に相補的ではない2本鎖から成るDNAは,ヘテロデュプレックスheteroduplexと呼ばれるが,その相補的でない部分を電子顕微鏡などで検出することができる。
糸を強く撚(よ)ると,糸は自分自身の周りに巻きついて超らせん構造を形成するが,DNAでも同様のことが起こり,現に細胞から抽出した環状DNAはほとんどが超らせん構造をとっている。この場合,撚りの方向は通常,二重らせんをほどく向きである。したがって,超らせんによるゆがみを取ろうとすると,二重らせんがほどけてDNAは部分的に1本鎖に分かれる(図3-a)。複製や転写の際にDNAは部分的に1本鎖に分かれる必要があるので,DNAを撚ったり撚りを戻したりすることにより,これらの機能の制御をしている可能性が大きい。実際,そのような働きをする酵素DNAトポイソメラーゼが発見されている。またDNA上でタンパク質が結合する個所は,塩基配列が対称性をもち,他方の鎖を逆向きに読むと,元と同じ配列になっていることが多い。このような配列を回文構造palindromeという。これは,二量体や四量体のタンパク質と対称性を適合させるためと思われる。また,このような配列ではDNAを十字型に変えても塩基対ができるので,特別な立体構造が目印になっていることも考えられる(図3-b)。
1970年代の中ごろになって,DNAの塩基配列を直接決定する方法が開発された。これにより,適当な長さのDNA断片を十分量精製できれば,遺伝情報を読むことができるようになった。この方法の一つのギルバート=マクザムGilbert-Maxam法(図4)では,DNA断片の末端を放射性同位体32Pで標識する。そして,特定の塩基の所で切断した後,ゲル電気泳動法により標識をもったDNA断片の長さを調べる。これにより標識端からいくつ目に切断を受けた塩基があるかわかるという原理である。
生物によって,DNAはいろいろな存在様式をとる。多くのウイルスでは,DNA分子は1g/ml近い最高に凝縮した状態で存在する。また,ウイルスの中には例外的に,1本鎖のDNAをもつものや,DNAの代りにRNAを遺伝子としてもつものもある。細菌DNAは,両端がつながった環状をしている。そして,種々のタンパク質やRNAを結合し,中程度に凝縮した状態で,細胞内のある領域に偏在する。しかし,それを包む核膜は存在しない。細菌細胞内には,ほかにプラスミドと呼ばれる小さな環状DNAの存在する場合がある。細菌の接合に関係するF因子や薬剤耐性を与えるR因子がその例である。真核生物のDNAは,核膜に包まれた核に存在し,ヒストンという塩基性タンパク質の周りに巻きついたヌクレオソームnucleosome構造のつながりからできている(図5)。この構造はさらに複雑に折りたたまれて,いわゆる染色糸を形成するが,高次の折りたたまれ方はまだ十分に解明されてはいない。真核生物細胞には,核外にもミトコンドリアや葉緑体中に小さな環状DNAが存在する。また細菌のDNA中には,IS(insertion sequence,挿入断片)や二つのISで挟まれたトランスポゾンtransposonという特殊な塩基配列があって,これらは低頻度でDNAの上を飛び移り,挿入や欠失などの突然変異を起こしている。類似のものは真核細胞DNA中にも存在し,進化や遺伝子発現の調節に占める役割が注目されている。
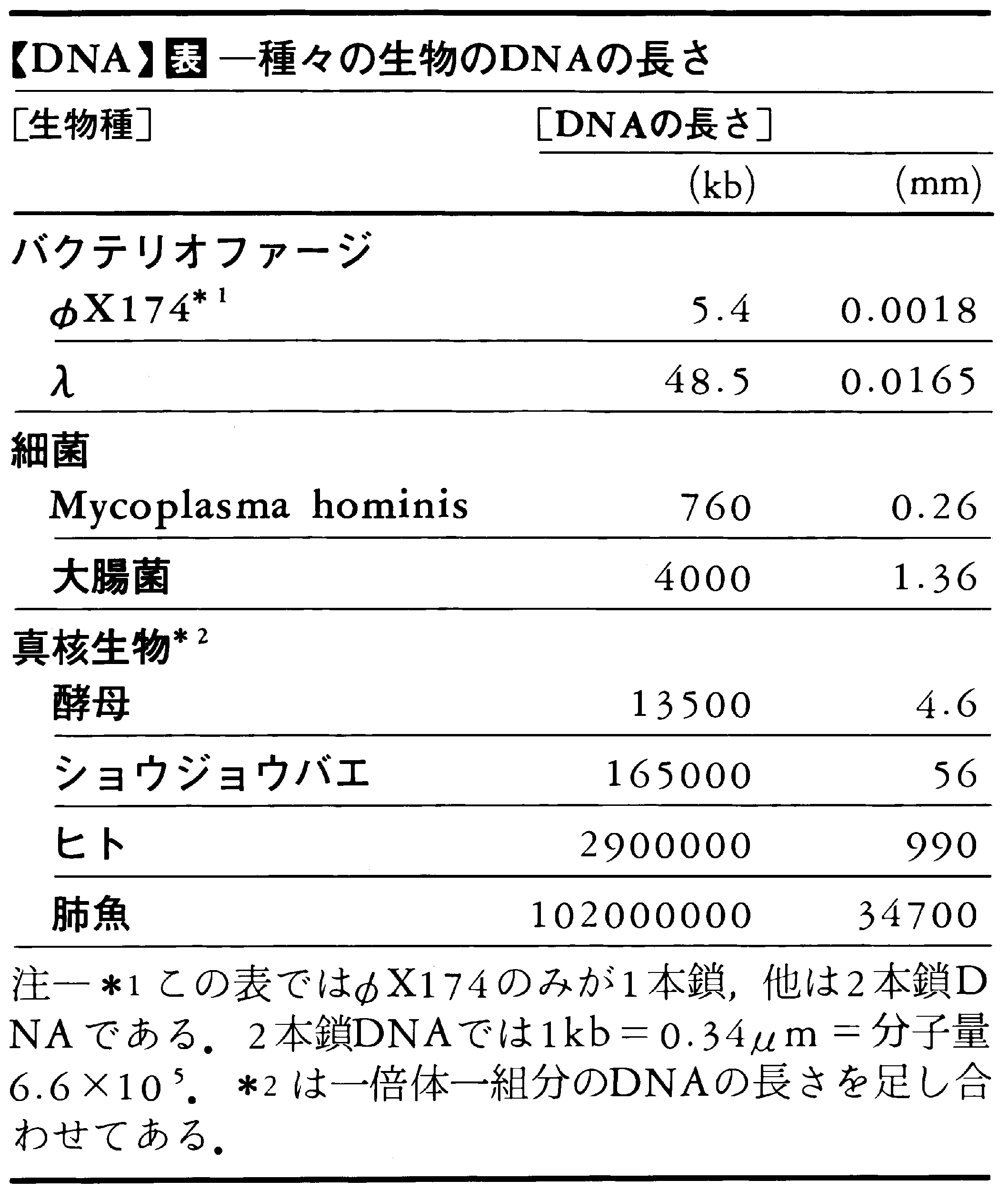
各生物の遺伝情報ひとそろいに対応するDNAの長さを表に示す。遺伝子一つは,平均して300~400アミノ酸残基から成るタンパク質を作るので,約1kb(キロベース=1000塩基対)の長さである。したがって,大腸菌DNAは,約4000ベースあるから,約4000の遺伝子から成るといえる。しかし高等生物では,高度反復配列やイントロン(後出)などアミノ酸残基と対応していない部分が多いので,遺伝子数はこのような計算値よりかなり小さいと考えられる。高等生物の生殖細胞はおのおのこの表の量のDNAを含むが,体細胞は通常二倍体なので,この倍量のDNAを含む。
遺伝情報としての塩基配列はどのように読みとられるのであろうか。実際の遺伝子とその転写調節の例として,大腸菌の乳糖分解遺伝子群(ラクトース・オペロン)を図6に示す。その特徴は以下の通りである。(1)関連ある機能の遺伝子がいくつかDNA上に隣接して並んでおり,その情報発現は同時に起こり,1本のmRNAとして転写される。この単位をオペロンoperonという。(2)タンパク質のアミノ酸配列に対応する塩基配列より5′末端側に,その発現を調節する塩基配列がある。(3)転写酵素(RNAポリメラーゼ)は,調節部位のうちプロモーターpromoterと呼ばれる部位を認識し,その下流(3′末端側)のある地点より,DNAと対応する塩基配列をもつmRNAを5′末端から3′末端の向きに合成する。(4)しかし転写が行われるためには,ほかにDNAのプロモーター部位にキャップ(CAP)タンパク質が結合しており,オペレーター部位にリプレッサーrepressorというタンパク質が結合していないことが条件である。培地にブドウ糖(グルコース)が存在するとキャップタンパク質は失活し,乳糖(ラクトース)が存在するとリプレッサーが失活するので,ブドウ糖が存在せず乳糖が存在するときのみ転写は行われる。ラクトース・オペロン以外でも,転写の活性化(キャップタンパク質に相当)や抑制(リプレッサーに相当)をする制御タンパク質により(片方だけのことも多い),遺伝子発現が調節されていると思われる。
これに対し真核生物DNAの転写では,ふつう一つの遺伝子に対して1本のmRNAが合成される。また,一つのタンパク質のアミノ酸配列に対応する塩基配列(エクソンexonと呼ばれる)の間に別の塩基配列(イントロンintronと呼ばれる)が挿入されている。イントロンはエクソンとともにmRNAに写されるが,その後,mRNAが熟成するときに切り捨てられる。転写調節の詳しい機構はいまだ研究中である。
DNAの複製は半保存的に行われる。すなわち,DNAの2本鎖が引き離されて,そのおのおのに対して新しく相補的な鎖が合成される。合成は,DNAポリメラーゼという酵素により,4種のデオキシヌクレオシド三リン酸(デオキシアデノシン三リン酸dATP,デオキシグアノシン三リン酸dGTP,デオキシシチジン三リン酸dCTP,デオキシチミジン三リン酸dTTP)を原料とし,DNA鎖とピロリン酸(P-P)を生ずる。この反応は次のように書ける。
n1・dATP+n2・dGTP+n2・dCTP+n1・dTTP─→DNA+(2n1+2n2)P-P
なお,複製の詳しい機構については〈DNA複製〉の項目を参照されたい。
前述の転写と複製のほかに,DNAの関与する機能として,突然変異,組換え,修復,逆転写,制限と修飾がある。突然変異の多くは,一つの塩基が他の塩基に置き変わった点突然変異だが,ほかに欠失・付加・置換・重複・逆位などの変異もある。大多数の変異は生物にとって有害か中立的でしかないが,突然変異により生物は多様性を増し,進化が可能になったことも忘れてはならない。組換えは,いろいろな変異をもったDNAをつなぎ換えて遺伝的に新しい組合せを作る。通常の普遍的組換えは,2本のDNA鎖が部分的に同じ塩基配列をもっていれば,どんな塩基配列でも起こる。このほかに,特別の塩基配列の所でのみ起こる特定部位組換えや,同じ塩基配列でない部分の間にひじょうに低い頻度で起こる非規則的組換えがある。修復は,紫外線,放射線,薬品などでDNAの塩基や鎖が傷ついたときに,これを元に戻す機構である(〈DNA修復〉の項目を参照)。
DNAの代りに例外的にRNAを遺伝子としてもつ動物ウイルスの一部のものは,宿主に感染後,遺伝子RNAに相補的なDNAを合成する。これを逆転写という(〈RNA依存性DNAポリメラーゼ〉の項目を参照)。また多くの生物は,DNAの特定の塩基配列(4または6塩基対の回文構造が多い)を認識して切断する制限酵素をもち,外から侵入したDNAを切断する。これらの生物は,同時に同じ塩基配列を認識して塩基をメチル化する修飾酵素をもつ。メチル化された塩基配列は制限酵素が切断しないので,自殺を免れるようにできている。
DNAを人為的に操作する技術は,最近急速に発達してきた。任意の塩基配列をもつDNAの人工合成は,ヌクレオチドやそれが少数つながったオリゴヌクレオチドを縮合剤を用いて結合させることにより行われる。このときに,リン酸基をトリエステルにしておくと,縮合の効率がよい(リン酸トリエステル法)。塩基上の反応しやすい基は,あらかじめ修飾して保護しておき,DNA鎖が完成してから保護基を外してもとに戻す。現在,20塩基程度のポリヌクレオチドは,比較的容易に合成できるようになった。
遺伝子工学では,試験管内DNA組換えという方法が用いられる。これは,プラスミドやバクテリオファージなどの自律的に増殖できるDNA(ベクターvectorという)に,例えば高等生物からとった増やしたいDNA断片を試験管内で挿入することである。この場合,DNAを切るには種々の制限酵素,つなぐにはDNAリガーゼという酵素を用いる。作成した人工組換え体をベクター本来の宿主細胞に入れるとベクターは増殖し,挿入したDNAを大量に増やすことができる。また,挿入したDNAの前に宿主に適合したプロモーターを置くなどの配慮をすると,挿入した遺伝子の産物を生合成させることも可能である。人工組換えDNAを宿主細胞に入れるには,宿主細胞を適当に処理して裸のDNAを取り込ませる形質転換法と,DNAを試験管内でバクテリオファージ粒子に取り込ませ,そのバクテリオファージを宿主細胞に感染させる方法とがある。
DNAを操作することは,生命の設計図の中枢部をいじることになる。人間に応用したとき,それは遺伝病の治療など大きな益を生み出す可能性をもつが,また,十分に注意深く適用すべき技術であることはいうまでもない。
執筆者:桂 勲
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
deoxyribonucleic acidの略。デオキシリボース,リン酸,塩基からなる核酸。デオキシリボ核酸とも。生物の遺伝情報の物質的な本体といえる。塩基成分はアデニン(A),グアニン(G),シトシン(C),チミン(T)の4種類があり,これらの並び順(塩基配列)が遺伝情報となる。塩基配列は種や個体によって異なるため,分子系統学やDNA鑑定に利用される。古代の生物標本や,土壌や水などの環境中に存在するごく微量なDNAの塩基配列を決定することも可能になっている。
執筆者:二階堂 雅人
出典 平凡社「最新 地学事典」最新 地学事典について 情報
Sponserd by ![]()
deoxyribonucleic acidの略称.[同義異語]デオキシリボ核酸
出典 森北出版「化学辞典(第2版)」化学辞典 第2版について 情報
Sponserd by ![]()
Sponserd by ![]()
出典 内科学 第10版内科学 第10版について 情報
Sponserd by ![]()
「デオキシリボ核酸」のページをご覧ください。
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
…1910年代に入って,T.H.モーガンらはキイロショウジョウバエを用いて,遺伝子と染色体部分の直接的な結びつきを証明し,遺伝に染色体という物質的な基礎を与えた。現在,この分野では種々の遺伝学的現象を細胞学的現象に関連・対応づけるため,核や細胞小器官(オルガネラ)自体およびそれらに含まれる染色体やDNA分子の構造・複製・伝達・化学的組成などを光学・紫外線・蛍光あるいは電子顕微鏡を用いて研究している。
[突然変異遺伝学mutation genetics]
1920年代の半ばに入り,H.J.マラー(1927)はショウジョウバエ,スタッドラーL.J.Stadler(1928)はトウモロコシにX線照射を行い,遺伝子の突然変異が人為的に誘発できることを証明した。…
…遺伝情報と遺伝的形質との間の関係は,ビードルG.W.Beadleらのアカパンカビを用いた遺伝生化学的研究により,〈一つの遺伝子は一つの酵素を作る〉という概念が誕生したことで,さらに具体的に理解できるようになった。一方,遺伝情報を実際に蓄えている物質については,20世紀中ごろにおこなわれた一連の実験により,DNA(デオキシリボ核酸)がその本体であることが確定した。たとえば,肺炎双球菌の無毒株が有毒株から精製したDNAを取り込むことで有毒株になる(形質転換)というO.T.エーブリーらの実験,そして,大腸菌のウイルスが増殖するためには,タンパク質ではなくDNAが必要であることを証明したハーシーA.D.HersheyとチェースM.Chaseの実験などである。…
…1947年生物学に転じ,生命の本質を生気論的な瞑想を排して,構成分子の構造からとらえようと考え,キャベンディシュ研究所でタンパク質構造の研究を始めた。そこでアメリカのファージグループ出身のワトソンと出会い,DNA(デオキシリボ核酸)の構造解明の重要性を確信し,X線結晶学者M.H.F.ウィルキンズやR.E.フランクリンらの得たDNAのX線回折像およびE.シャルガフらの化学分析値などから,53年J.D.ワトソンとともにDNAの二重らせん分子構造モデルを提出する。このモデルは遺伝子がDNAであることを多くの研究者に確信させ,その後の分子生物学の方向を定め,急激な発展をもたらすことになった。…
…1910年代に入って,T.H.モーガンらはキイロショウジョウバエを用いて,遺伝子と染色体部分の直接的な結びつきを証明し,遺伝に染色体という物質的な基礎を与えた。現在,この分野では種々の遺伝学的現象を細胞学的現象に関連・対応づけるため,核や細胞小器官(オルガネラ)自体およびそれらに含まれる染色体やDNA分子の構造・複製・伝達・化学的組成などを光学・紫外線・蛍光あるいは電子顕微鏡を用いて研究している。
[突然変異遺伝学mutation genetics]
1920年代の半ばに入り,H.J.マラー(1927)はショウジョウバエ,スタッドラーL.J.Stadler(1928)はトウモロコシにX線照射を行い,遺伝子の突然変異が人為的に誘発できることを証明した。…
…同じことは高等動植物の若干の遺伝子についても証明されてきた。一方,遺伝子の本体がDNAであることが確実になり,バクテリアやウイルスでは一つのゲノムの全遺伝子が単一のDNA分子に組み込まれていること,また,高等動植物においても電子顕微鏡で確認できるかぎりでは,1本の染色体に含まれるDNAは連続した1本の糸であることもわかり,遺伝子の統一概念は自然に消滅した。
[遺伝子の作用――転写と翻訳]
1950年代以降の分子遺伝学の発達により,まず,遺伝子の本体とポリペプチドの生産を支配する機構の概要が明らかとなった。…
…遺伝情報と遺伝的形質との間の関係は,ビードルG.W.Beadleらのアカパンカビを用いた遺伝生化学的研究により,〈一つの遺伝子は一つの酵素を作る〉という概念が誕生したことで,さらに具体的に理解できるようになった。一方,遺伝情報を実際に蓄えている物質については,20世紀中ごろにおこなわれた一連の実験により,DNA(デオキシリボ核酸)がその本体であることが確定した。たとえば,肺炎双球菌の無毒株が有毒株から精製したDNAを取り込むことで有毒株になる(形質転換)というO.T.エーブリーらの実験,そして,大腸菌のウイルスが増殖するためには,タンパク質ではなくDNAが必要であることを証明したハーシーA.D.HersheyとチェースM.Chaseの実験などである。…
…嫡出否認(夫婦間に出生した子すなわち嫡出子を父親が否認する),認知,親子関係不存在確認(もらい子などが偽りの届出により嫡出子になっているが,ほんとうは親子でないことを確認する)などの場合や,まれではあるが病(産)院で子を取り違えた疑いが生じた際などに,当事者間に生物学的な親子関係が存在するかどうかを自然科学的に検査して決めること。これには古くから,規則正しい単純な遺伝を示す血液の多型形質が利用されてきたが,最近ではDNA多型(DNA型。ゲノムDNAやミトコンドリアDNA上の塩基配列の差によって生じる遺伝的個体差)が検査の主役を担うようになっている。…
…最初ミーシャーF.Miescherにより1869年ころに細胞核(核)の成分として発見されたが,細胞核だけの成分ではなく細胞質にも含まれる。核酸は化学構造上DNA(deoxyribonucleic acid,デオキシリボ核酸)とRNA(ribonucleic acid,リボ核酸)の2種に大別できるが,この2種には細胞内における分布や生物学的役割に大きな差がある。DNAは真核生物の核,葉緑体,ミトコンドリア,原核生物の細胞ならびに多くのウイルスの粒子内に含まれる。…
…1947年生物学に転じ,生命の本質を生気論的な瞑想を排して,構成分子の構造からとらえようと考え,キャベンディシュ研究所でタンパク質構造の研究を始めた。そこでアメリカのファージグループ出身のワトソンと出会い,DNA(デオキシリボ核酸)の構造解明の重要性を確信し,X線結晶学者M.H.F.ウィルキンズやR.E.フランクリンらの得たDNAのX線回折像およびE.シャルガフらの化学分析値などから,53年J.D.ワトソンとともにDNAの二重らせん分子構造モデルを提出する。このモデルは遺伝子がDNAであることを多くの研究者に確信させ,その後の分子生物学の方向を定め,急激な発展をもたらすことになった。…
…この現象は病原性形質が死菌から生菌に移行することによる生菌の遺伝形質の転換を意味するところから,形質転換と呼ばれるようになった。ついで44年O.T.エーブリーらは形質転換がDNAによって引き起こされることを明らかにした(図2)。これは,遺伝物質がDNAであることを直接的に示した最初の研究である。…
…生体,死体,あるいは死体の一部がだれであるかを決定すること。ときには人体に由来するもの,たとえば血液・唾液・精液などの体液・分泌物(およびそれらに含まれるDNA),あるいは指紋・足跡その他の痕跡なども個人識別の対象になる。生体では,氏名を偽ったり黙秘する事件の容疑者,あるいはまれには精神異常者,乳幼児などが対象となる。…
… 遺伝現象を支配している法則は,基本的にはヒトでも他の生物でも同じである。ヒトの配偶子には,22本の常染色体と1本の性染色体(精子ではXまたはY染色体,卵子ではX染色体)が存在し,その中に全部で約28億塩基対のDNAが含まれ,DNAの特定の領域が遺伝子として働いている。ヒトの体細胞には,22対の常染色体と1組の性染色体(男ではXY,女ではXX)が存在し,その中に約56億塩基対のDNAが含まれていることになる。…
…もともとは生物であったのかもしれないが,他の生物の細胞に〈寄生〉して生きるようになったため,情報のシステムだけが残ってしまったものと考えてもよいだろう。 すべての生物がもつ基本的な情報システムは遺伝子DNA(一部のウイルスはRNAを遺伝物質としている)であるが,細菌,ラン藻のように初期の生物の姿をとどめていると考えられる生物では,遺伝子DNAが裸のまま細胞の中に収まっている。しかし,すべての動物,植物では,遺伝子DNAは染色体の中に収められ,この染色体の集団が細胞分裂の間期には核膜に包まれて,いわゆる核を形成している。…
… 分子生物学は,生物現象を要素的,還元的にとらえる。要素は生体分子であり,DNA,RNA,タンパク質分子が生物を構成する基本単位となる。中心概念は,生物を分子的な機械とみなすところにある。…
※「DNA」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
夏の暑さに体が慣れること。数日から数十日間で起こる短期暑熱順化と、数年または数世代にかけて起こる長期暑熱順化とがある。→寒冷順化[補説]近年では、冷房設備の普及にともない短期暑熱順化が起こりにくくなっ...