デジタル大辞泉 「突然変異」の意味・読み・例文・類語
とつぜん‐へんい【突然変異】
Sponserd by ![]()
翻訳|mutation
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
生物にみられる種々の変異のうち、遺伝子の量的または質的な変化によって生じた変異をいう。したがって突然変異は、それが致死作用をもっていない限りは子孫の細胞や個体に伝えられる。ただし、すでに存在する遺伝子の分離や遺伝的組換えによって新しい形質が現れる場合は含まれない。突然変異のことばを生物学上で初めて使用したのはオランダのド・フリースで、オオマツヨイグサOenotheraにみいだされた遺伝的な変わりものに対して名づけた(1901)。その後、アメリカのT・H・モーガンが、ショウジョウバエ属Drosophilaで白眼の突然変異をみいだし(1910)、さらにH・J・マラーが、ショウジョウバエにX線を照射して人為的に突然変異を作成することに成功して(1928)、その後の遺伝学の発展に大きく貢献した。
突然変異は、遺伝子の本体であるDNA(デオキシリボ核酸)の塩基の一つが他の塩基に置き換わった分子レベルの変化から、遺伝子の存在する染色体の構造や数の変化したものなど、種々の程度の変化が含まれる。
[黒田行昭]
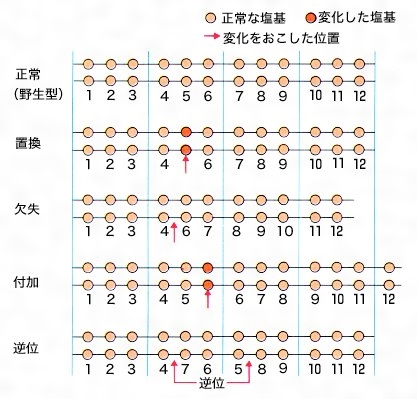
まず、DNAの一つまたは多数の塩基の置換base change、欠失deletion(またはdeficiency)、付加addition、逆位inversion、重複duplicationなどや塩基の枠組み移動flame shiftのような分子レベルでの変化がある。この場合は、そのDNAのもつ遺伝情報によってつくられる酵素やタンパク質がつくられなくなったり、活性が消失したりするような変化が現れる。このような一つの遺伝子の作用に影響を与えるような突然変異を遺伝子突然変異gene mutationまたは点突然変異point mutationという。
遺伝子の存在する染色体の構造や数の変化によって生ずる突然変異を染色体突然変異chromosome mutationまたは染色体異常chromosome aberrationという。染色体の数の変化としては、染色体の全体のセット(ゲノム)が整数倍で増加する倍数性polyploidyや、ある特定の染色体のみが1本増加したり減少したりするトリソミーtrisomyやモノソミーmonosomy、数本の染色体が増減する異数性heteroploidyなどがある。また、染色体の構造の変化としては、1本の染色体のなかでの欠失や重複、逆位などのほか、ほかの染色体との間でのつなぎ換えとして転座translocationがある。
これらの突然変異によって生じた変化が、形態的な形質として認められる場合は形態的突然変異morphological mutationまたは可視突然変異visible mutationという。また、生じた突然変異によって細胞や個体が生存できなくなるものを致死突然変異lethal mutationという。正常または野生型の細胞や個体が突然変異をおこす場合を前進突然変異forward mutationといい、突然変異をおこした細胞や個体が、同じ場所に突然変異をおこして、ふたたびもとの正常または野生型に戻る場合を復帰突然変異reverse mutation(またはback mutation)という。
[黒田行昭]
突然変異は、外から人為的に手を加えなくとも、自然の状態においてもある一定の低い頻度でおこる。これを自然突然変異spontaneous mutationというが、その頻度は生物の種類や遺伝子の種類によって異なり、1遺伝子座当り世代当りの頻度は、微生物では1億から10億回に1回の割合で非常に低いが、高等動植物では10万から100万回に1回の割合と比較的高い。ヒトでは四肢短縮症が10万回に5回くらい、血友病が10万回に3回くらい、無虹彩(むこうさい)症や小眼球症がそれぞれ100万回に1回くらいの割合でおこる。
X線やγ(ガンマ)線、紫外線などの放射線や種々の変異原性化学物質、発癌(はつがん)物質などを作用させると、突然変異の頻度が人為的に上昇する。このような人為的に誘発された突然変異を誘発突然変異induced mutation(人為突然変異)という。そのときの突然変異の頻度は、使用した放射線の線量や照射時間、化学物質の濃度や処理時間などに比例して増大する。
[黒田行昭]
突然変異が人為的につくられるようになってから、これらの突然変異を利用して、多くの生物の遺伝子の連関関係の研究や、種々の生体構成成分の合成や分解の生化学的代謝過程の解析が行われ、遺伝学、発生学、生物学の各学問分野における貢献は計り知れないものがある。また、突然変異の実際的な利用としては、植物の品種改良など育種学、農学、畜産学などの各分野における利用のほか、ペニシリン、ストレプトマイシンなど多くの抗生物質を多量に生産する自然および人為突然変異の菌株の開発や改良、特定のアミノ酸やタンパク質その他の有用物質を多量に生産する微生物の株の開発などにも利用されている。また1970年ごろから、食品や医薬品、農薬など環境中に存在する多数の変異原物質や発癌物質の人間に対する危険度を予測し、これらを環境中からできるだけ排除するために、微生物や哺乳(ほにゅう)類の培養細胞、昆虫、植物、実験動物など種々の生物の検出系を使って、これらの物質の突然変異性の調査も行われている。
[黒田行昭]
『芦田譲治他編『遺伝と変異』(1958・共立出版)』▽『山口彦之編『植物遺伝学4 形態形成と突然変異』(1977・裳華房)』▽『山口彦之他著『突然変異育種』(1983・養賢堂)』▽『R・H・タマリン著、木村資生監訳、福田一郎他訳『遺伝学』上下(1988・培風館)』▽『森脇大五郎著『遺伝学ノート――ショウジョウバエと私』(1988・学会出版センター)』▽『大島康行他著『新版 図説生物学』(1988・朝倉書店)』▽『R・Y・スタニエ他著、高橋甫他訳『微生物学』上(1989・培風館)』▽『黒田行昭編『遺伝学実験法講座5 動物遺伝学実験法』(1989・共立出版)』▽『D・M・フライフェルダー著、志村令郎他訳『分子生物学』上(1989・化学同人)』▽『岡田吉美著『プロテインエンジニアリング』(1989・東京化学同人)』▽『江上信雄他編『メダカの生物学』(1990・東京大学出版会)』▽『蓬原雄三著『イネの育種学』(1990・東京大学出版会)』▽『近藤喜代太郎他著『人類遺伝学の基礎』(1990・南江堂)』▽『柴忠義著『バイオテクノロジー実験操作入門』(1990・講談社)』▽『名取俊二編『薬科分子生物学』(1990・講談社)』▽『P・J・ラッセル著、太田次郎監訳、今泉佐枝子・清水久美子訳『現代遺伝学』(1990・オーム社)』▽『根井正利著『分子進化遺伝学』(1990・培風館)』▽『J・F・クロー著、木村資生・北川修・太田朋子訳『遺伝学概説』(1991・培風館)』▽『G・エドリン著、清水信義監訳、伊藤文昭他訳『ヒトの遺伝学』(1992・東京化学同人)』▽『藤巻宏他著『植物育種学 基礎編』『植物育種学 応用編』(1992・培風館)』▽『蓬原雄三編著『育種とバイオサイエンス――育種学の新しい流れ』(1993・養賢堂)』▽『D・M・フライフェルダー他著、川喜田正夫訳『分子生物学の基礎』第2版(1994・東京化学同人)』▽『野沢謙著『動物集団の遺伝学』(1994・名古屋大学出版会)』▽『太田次郎他編『遺伝のしくみ』(1995・朝倉書店)』▽『黒田行昭編著『21世紀への遺伝学1 基礎遺伝学』(1995・裳華房)』▽『山口彦之著『放射線生物学』(1995・裳華房)』▽『大嶋泰治編著、駒野徹他編『酵母分子遺伝学実験法』(1996・学会出版センター)』▽『山本隆一他編『イネ育種マニュアル』(1996・養賢堂)』▽『三浦謹一郎編『分子遺伝学』(1997・裳華房)』▽『百瀬春生編『分子遺伝学』(1997・丸善)』▽『メイワン・ホー著、小沢元彦訳『遺伝子を操作する――ばら色の約束が悪夢に変わるとき』(2000・三交社)』▽『コリン・パターソン著、馬渡峻輔他訳『現代進化学入門』(2001・岩波書店)』▽『日本放射線技術学会監修、江島洋介・木村博編『放射線生物学』(2002・オーム社)』▽『E・A・バージ著、高橋秀夫他訳『バクテリアとファージの遺伝学――分子生物学・バイオテクノロジーの基盤』(2002・シュプリンガー・フェアラーク東京)』▽『スティーヴ・ジョーンズ著、ボリン・ヴァン・ルーン画、山元大輔訳『超図説 目からウロコの遺伝・DNA学入門――ダーウィンから遺伝子治療まで』(2003・講談社)』▽『鵜飼保雄著『植物育種学――交雑から遺伝子組換えまで』(2003・東京大学出版会)』▽『木村資生著『生物進化を考える』(岩波新書)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
生物のもつ遺伝物質が量的・質的に変化すること,およびその変化によって生じた状態を指す。細胞核内の遺伝物質の突然変異(核性突然変異)は,変化が起こる遺伝物質の単位に応じて,ゲノム突然変異,染色体突然変異,遺伝子突然変異に分けられる。優性突然変異,劣性突然変異という区別は,突然変異がヘテロ二倍体で表現型の変化をもたらすか否かの違いをいうのであり,また常染色体突然変異,性染色体突然変異という区別は,突然変異が起こった染色体の違いをいうものであるが,いずれも突然変異の分類としては基本的ではない。なお核性突然変異に対して,葉緑体やミトコンドリアなどの細胞小器官の遺伝物質の突然変異を細胞質性突然変異というが,細胞質性突然変異は伝達の様式(これについては〈細胞質遺伝〉の項目を参照)が特異的である点を除いて,突然変異の起こる機構としては核性突然変異の場合と基本的な違いはない。
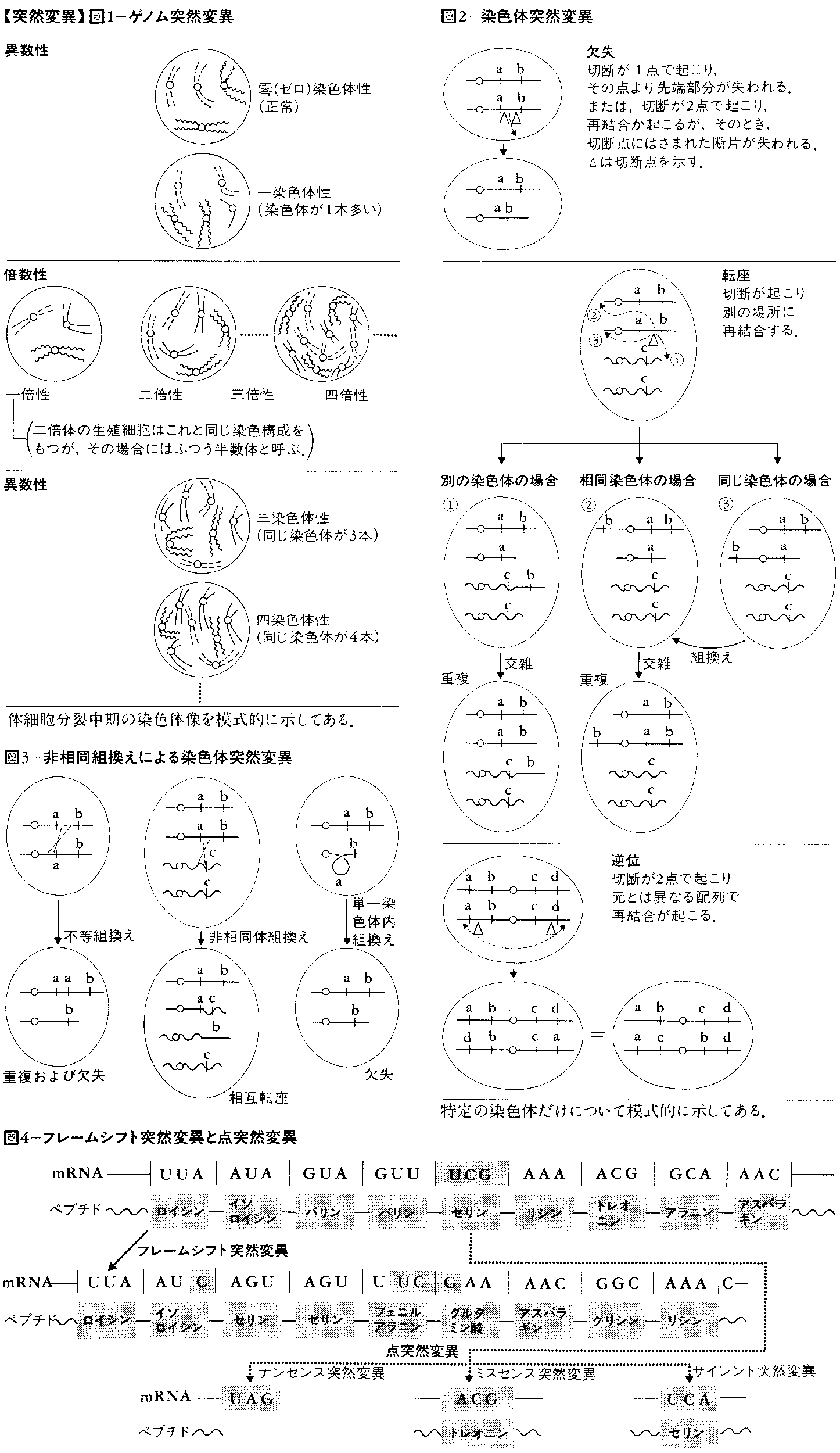
生物は種に固有の染色体構成(ゲノム)をもっているが,それが組として増減するのが倍数性であり,一部の染色体だけが増減するのが異数性である(図1)。これらをゲノム突然変異と呼ぶ。つまり,ゲノム突然変異は遺伝物質の量的な変化であり,遺伝物質である染色体の分配の異常によって生じる。これに対して,染色体突然変異と遺伝子突然変異は遺伝物質(染色体および遺伝子の本体であるDNA)の構造的変化であり,狭義にはこれらだけを突然変異という。
これは元来,光学顕微鏡で観察できる染色体の構造変化,たとえば体細胞分裂中期の染色体の形態,減数分裂中期の染色体対合の状態,ショウジョウバエの唾腺(だせん)染色体のような巨大染色体の縞模様などによって解析されるものを指したが,現在では遺伝解析,電子顕微鏡観察,化学分析などによって検出できる微小なものも含めるようになっている。その中には,染色体の一部(末端部分の場合も中間部分の場合もある)が消失した欠失deletion,染色体の一部が同一染色体の別の場所または別の染色体に移った転座translocation,染色体の一部が本来の場所に隣接または別の場所に付け加わった重複duplication,染色体の一部の方向性が逆になった逆位inversionなどがある(図2)。いずれも染色体の切断-再結合および不等組換え,非相同染色体組換えなどの異常組換えによって生じると考えられている(図3)。もっとも,遺伝的組換えが基本的にDNAの切断-再結合として理解されている現在では,染色体突然変異の第1原因はDNAの切断であると考えてよい。事実,DNAの切断を誘発する電離放射線によって染色体突然変異の出現頻度が増加することはよく知られている。
遺伝物質つまりDNAの構造変化が単一遺伝子内に限られている場合を遺伝子突然変異という。その中には,規模の小さい染色体突然変異といえるもの(塩基の付加additionまたは欠失によってDNAから転写されたメッセンジャーRNAがペプチド鎖に翻訳されるときに読む枠がずれるフレームシフトframe shift突然変異はこの中に含まれる)と一塩基だけが置換する点突然変異とがある。後者はさらに,遺伝子の途中に停止信号が生じたためにペプチド鎖の伸長が途中で止まる停止(ナンセンスnonsenseともいう)突然変異,ペプチド鎖のアミノ酸が一つだけ変化する(それによってそのペプチド鎖の機能が失われる場合と失われない場合とがある)ミスセンスmissense突然変異,ペプチド鎖の変化がみられないサイレントsilent突然変異とに分けられる(図4)。生体の機能的な変化をもたらす突然変異は比較的容易に識別できる。しかし,機能的に識別できないからといって突然変異をもっていないということではない。ミスセンス突然変異の一部やサイレント突然変異は機能的変化をまったくもたらさない。このような突然変異は,ペプチド鎖のアミノ酸配列やDNAの塩基配列の解析によって初めて検出されるもので,偶然に見いだされるのを待たなければならない。また,ある種の突然変異は,それ自体は表現型によって識別はできないが,他の突然変異との相互作用によって検出できる。抑制(抑圧)突然変異はその一例で,転移RNAやリボソームタンパク質の遺伝子の突然変異によって,翻訳に変化が生じたものであることが明らかになっている。なお,DNAのすべての部分が現時点で発現している遺伝情報をもっているわけではなく,そのような部分に起こった突然変異についての理解は今後の研究を待たなければならない。
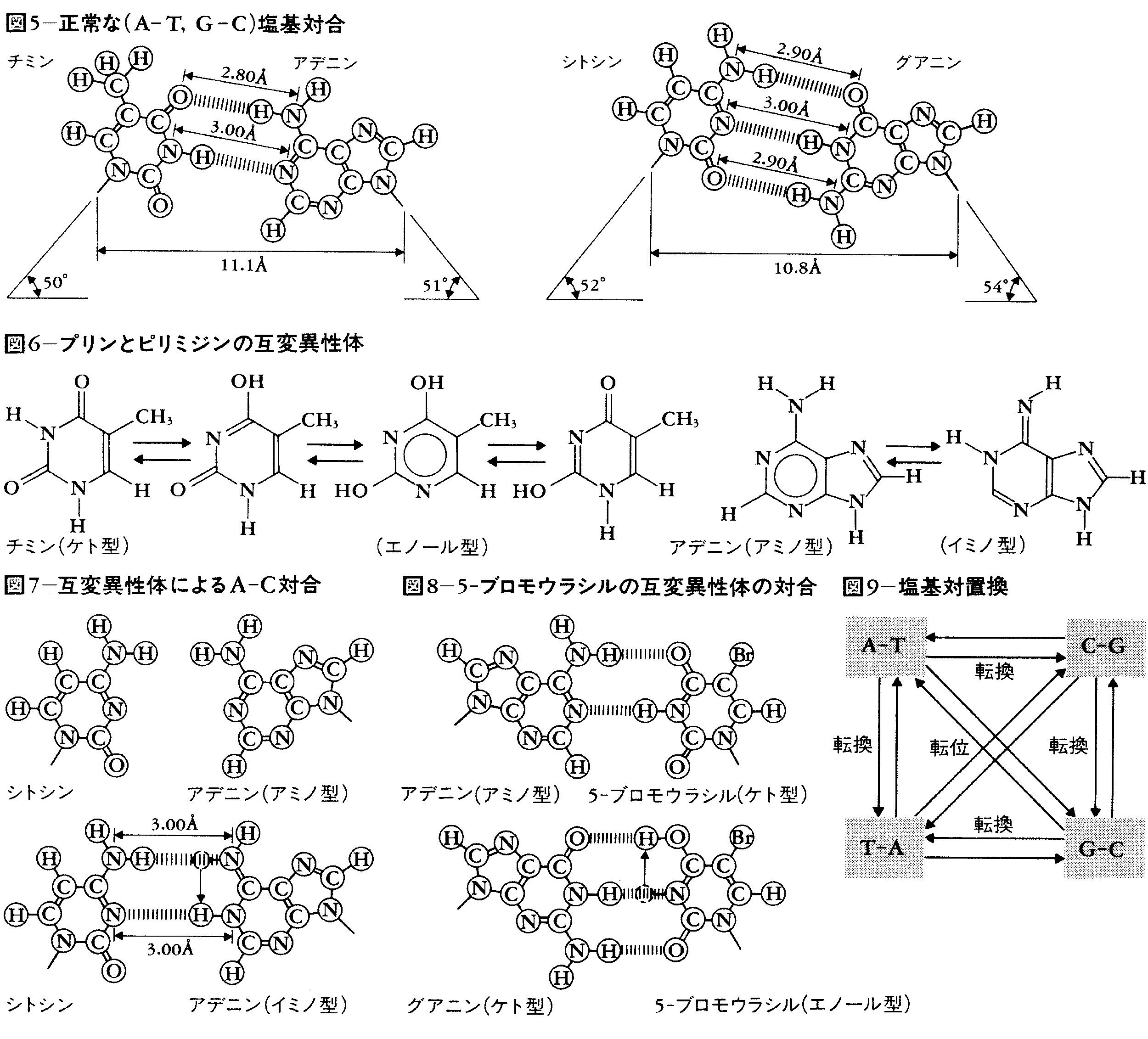
DNAはアデニン(A),チミン(T),グアニン(G),シトシン(C)という4種の塩基の配列によって遺伝情報を構成しているのであるが,その構造の安定性は,A-T,G-Cの特異的塩基対合によっている(図5)。ところが,これらの塩基は互変異性体転換をする(図6)。互変異性体転換の平衡はアミノ型とケト型にかたよってはいるが,きわめて低い確率(10⁻6~10⁻9といわれている)でイミノ型とエノール型に転換する。DNA合成の時点でこの転換が起こると,A-C,G-Tという対合(図7)が可能になる。つまり,DNAはその物質としての物理化学的特性によって,遺伝物質として必須である安定性と自己複製能を担っているが,同時にきわめて低い頻度での突然変異の出現の運命も担っているといえる。
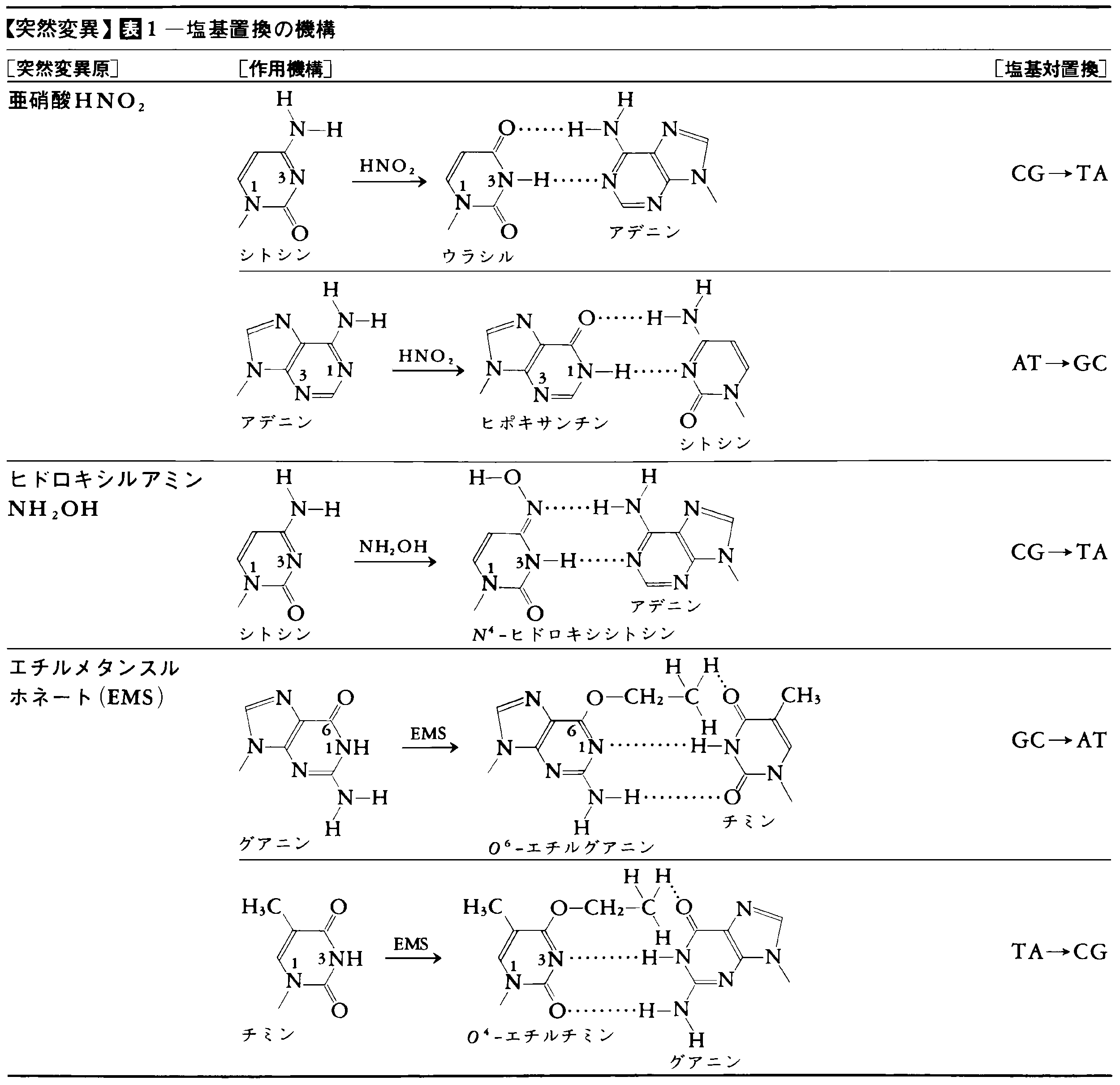
5-ブロモウラシルが突然変異を誘起するのは,これがチミンの構造類似体としてDNAに取り込まれるが互変異性体変換(図8)によって,A-T⇄G-Cの塩基対転位を起こすからである。これとは別に,化学物質の中には,塩基と化学的に反応してその構造を変えるものがある。亜硝酸やヒドロキシルアミン,そしてアルキル化剤であるエチルメタンスルホネートなどはその例である(表1)。これらの化合物はいずれもA-T⇄G-Cという塩基対転位型突然変異を誘発する。塩基対置換の種類としてはピリミジン塩基がピリミジン塩基に,プリン塩基がプリン塩基に代わる塩基対転位と,ピリミジン塩基がプリン塩基に,プリン塩基がピリミジン塩基に置換する塩基対転換がある(図9)が,後者の出現機構についてはよくわかっていない。また,フレームシフト突然変異を誘発する物質としてはアクリジン系色素が知られているが,それはこの色素がDNAの塩基対と塩基対との間に入り込むのに適した構造をもっているためと考えられる。
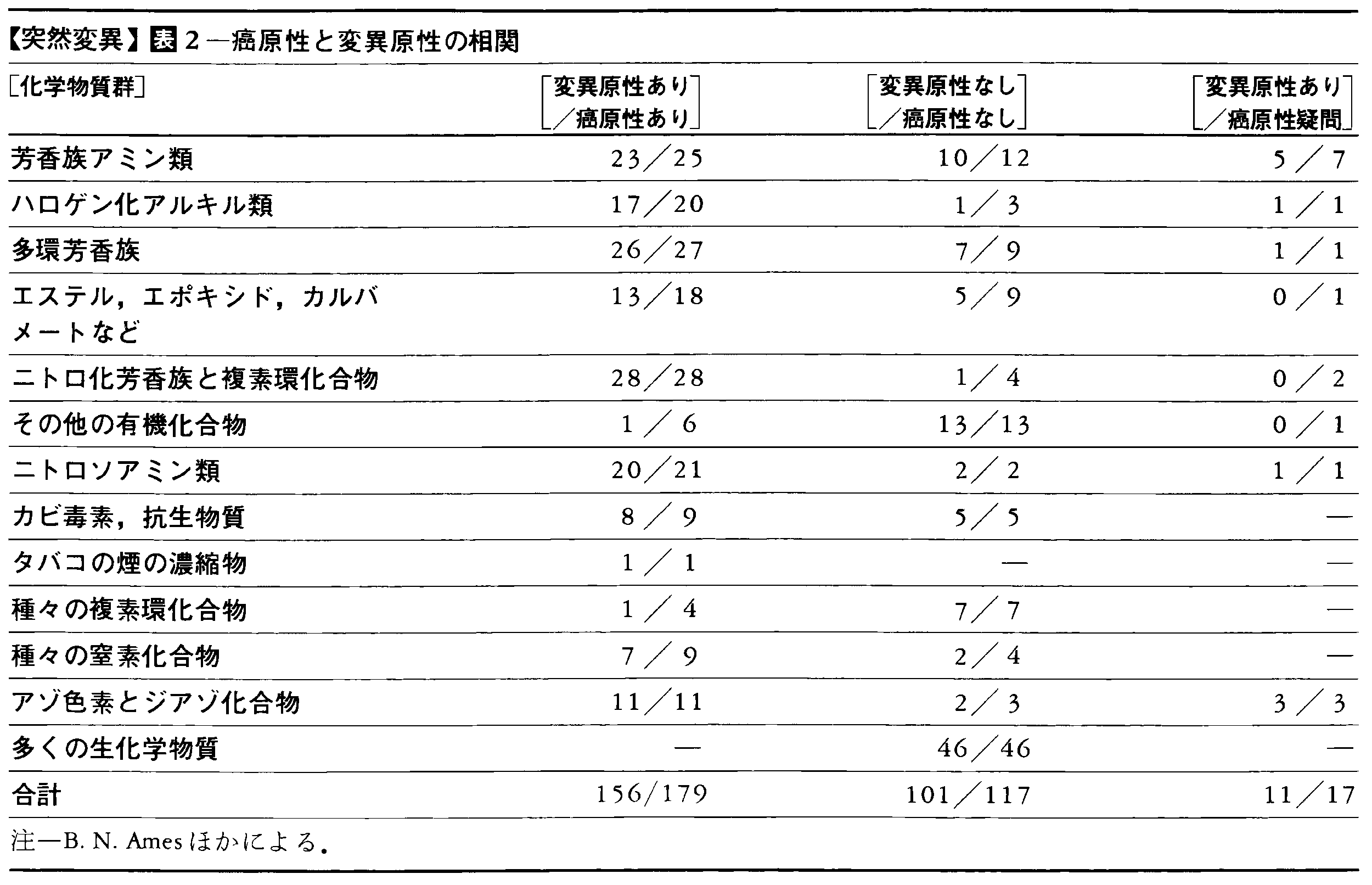
突然変異が生殖細胞に生じた場合には,次代に伝えられ,さらにその生殖細胞を通して後代に伝えられる。しかし,DNAの変化は体細胞にも起こりうる。これを体細胞突然変異somatic mutationと呼ぶが,この場合,突然変異は次代には伝わらず,その影響はその世代限りであり,しかも,その突然変異をもつ細胞で形成されている身体の部分に限られる。体細胞突然変異の生物学的意義は完全には理解されていないが,生物の老化が体細胞突然変異の蓄積に起因するという説が有力である。また,多細胞生物の細胞分化に体細胞突然変異が関与している可能性も考えられており,これらは今後の研究課題となるであろう。さらに,天然と人工とを問わず,種々の化学物質が突然変異を誘発する性質(突然変異原性)があることがネズミチフス菌Salmonella typhimuriumを使ったエームス法によって明らかにされている。その結果,癌を誘発する性質(癌原性)の物質の多くが突然変異原性をもつことが示された(表2)。変異原性物質の多くが癌原性であることと合わせて,このことは癌の少なくとも一部が体細胞突然変異によるものであると思わせる。
電離放射線や化学物質がDNAに作用して突然変異を誘発することを述べたが,DNAに生じた変化がすべて突然変異になるのではない。その大部分は,生命が進化の過程で獲得した機構によって修復される。修復機構の中には,本来の塩基配列を忠実に復元するものだけでなく,DNAの複製の障害の除去を第1の目的として必ずしも本来の塩基配列を復元しないものもある。この場合,細胞は当面の死をまぬがれるが,多くの場合,突然変異をもつことになる。
C.ダーウィンは《種の起原》において,現存する生物種の多様性は進化の産物であるという考えを提唱した(1859)。彼は自然観察を通して,種内にも個体差があり,種の違いにも連続性があることに気づいたのである。そして,個体差が自然淘汰の原動力となり,適者生存の原理に基づいて種が形成されると考えたのである。ところで,ダーウィンは種内の変異が親から子に伝わることをある程度は認識していたが,その伝達の様式を明確にできなかったし,変異の由来を明らかにもしなかった。一方,G.メンデルはエンドウの交配実験によって,生物の表現型が形質という単位に分けられること,そしてある形質については親から子に規則的に伝わることを示した(1865)。そのような形質について,彼は形質を決定する因子(後に遺伝子と呼ばれるようになった)を想定し,それが変化することなく親から子に伝えられると考えたのである。遺伝子は,ダーウィンが考えた個体変異の原因の一つであることが明らかになった。しかし,遺伝子が不変のものであるとすれば,生物の進化を説明できなくなった。一方,H.ド・フリースは,オオマツヨイグサの遺伝の研究から,遺伝形質は必ずしもすべてが連続的な変化をするのではなく,非連続的な変化をする場合があることを見いだした(1901)。そして,彼は遺伝子は不変ではなく,まれに非連続的な変化(突然変異)をするという仮説を出した。今では,ド・フリースが観察したのは染色体内組換えおよびゲノム突然変異であって,真の突然変異ではなかったと考えられている。厳密な意味での遺伝子突然変異を最初に観察したのは1929年のことで,トウモロコシを使ったスタッドラーL.J.Stadlerとショウジョウバエを使ったH.J.マラーである。遺伝子突然変異はきわめて低い頻度(10⁻6~10⁻9)で起こるのが普通であるが,電離放射線や種々の化学物質によって高頻度に誘発されることがわかるとともに,これらの突然変異原は突然変異の出現機構の解明の研究に活用されるようになっただけでなく,得られた突然変異株を利用することによって,遺伝学が急速に発展することになった。突然変異株は基礎研究だけでなく育種にも利用され,突然変異育種というべき分野が確立された。さらにつけ加えていうならば,人為選抜によってはじめて,無方向に生じた突然変異から特定方向の突然変異を得ることができるのであるが,今や遺伝子工学の応用によって,特定方向に突然変異を起こさせることが可能になったという見方もできるであろう。
突然変異は生物の遺伝的変異を維持・増加する機構の一つである。ところで,生物の進化の原動力が自然淘汰であるとするダーウィン以来の考えに対して,木村資生は,生物集団内に蓄積されている遺伝的変異のほとんどには,淘汰に対する有利不利という差がほとんどないという知見に基づいて,進化の原動力は遺伝子の機会的浮動であるという中立説を唱えた(1968)。分子レベルでの研究成果も中立説と矛盾しないものが多いことから,近年,この説は大きく注目されている。ただし,生物の種の違いが分子レベルで解明されているといえない現在では,突然変異と進化の関係を十分に理解するのは困難である。これは生物学におけるこれからの大きな課題といえるだろう。なお,この問題を考える場合には,環境が進化におよぼす効果,進化の異なる段階での突然変異がもつ意義などについても留意すべきであろう。そして,生物進化に先立つ化学進化の段階も含めて,生命進化としての視点をもって理解にあたるべきことはいうまでもない。生命が不完全である時点では,突然変異は生命に有利に働くが,完成に近づくにつれてむしろ逆に働くと考えることもあながち根拠のない推定ではないだろう。現存する生物では,突然変異はおおむね不利に働いているように思われるからである。
執筆者:小野 文一郎
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
生物の形質の遺伝的変異.また,単に変異ということもある.遺伝学的,細胞学的見地から核遺伝と核外(細胞質)遺伝における突然変異があるが,核に起因する変異は,さらに染色体突然変異と遺伝子突然変異とに分けられる.染色体に起因する変異はいくつかの機能に同時に影響を与えるもので,染色体構造の変化(欠損,転座,逆位,重複など)によるものであるが,その実体はまだ明らかにされていないことが多い.遺伝子突然変異は遺伝子DNAの構造変化により起こる.DNAの変化は塩基または塩基対の変化であり,
(1)複製の際にプリン塩基がほかのプリン塩基に,ピリミジン塩基がほかのピリミジン塩基に置き換えられる変異(トランジション)と,プリン塩基とピリミジン塩基が置き換えられるような変異(トランスバージョン),および,
(2)塩基の付加や欠損による遺伝暗号のトリプレット(コドン)の乱れが生じる変異,
に分けて考えられる.これらは突然変異誘起因子によって誘発される.核外遺伝はオルガネラ(ミトコンドリアや葉緑体)DNAの構造変化によるものと説明される.
出典 森北出版「化学辞典(第2版)」化学辞典 第2版について 情報
Sponserd by ![]()
mutation
生物の遺伝的変異を意味し,形質の変化が遺伝子または染色体の変化によって引き起こされたことを指す。突然変異はそのレベルによって次の種類に分けられる。DNAの一つの塩基の変化,塩基の置換によるコドンのコードの変化,欠失(ある長さのDNAの喪失),重複(ある長さのDNAの重複),逆位(ある長さのDNAが180°回転),挿入(ある長さのDNAの挿入),転座(染色体の位置移動)。突然変異の用語は,もとは遺伝的な非連続的変異(いわゆるメンデル形質)に対して用いられていたが,遺伝的な連続変異についてもポリジーンによって説明されるようになり,単なる遺伝子の変化(分子レベルの化学変化)との区別は曖昧になった。突然変異はランダムだとされるが,いつどこでも生じるわけでなく,限定された範囲でしか生じない。例えばショウジョウバエの目の色の突然変異は白・桃・赤・褐色しかないなど。
執筆者:小寺 春人
出典 平凡社「最新 地学事典」最新 地学事典について 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
Sponserd by ![]()
…ある生物のもつ独特の構造や機能を作るために必要な情報は莫大な量に違いないが,この情報も結局はある程度の独立性をもった素情報ともいうべき〈遺伝子〉の集合として理解できることを示唆したのもG.J.メンデルの功績である。20世紀に入ってアメリカのT.H.モーガンらは,遺伝子のもつ情報の変化(突然変異)がおこること,およびその“異常な”情報がメンデルの法則に従って伝達することを示した。さらに彼らは,ある遺伝子の間ではかならずしも〈独立の法則〉があてはまらず,連鎖や交叉(こうさ)という現象がおこることを示し,その結果,一群の遺伝子は規則正しく直線状に配列していることが明らかにされた。…
…遺伝的変異は,例えばいろいろの血液型にみられるような変異で,多くの形態的,生理的形質がこのような遺伝的な変異をしめす。また,雌雄の違いやいわゆる突然変異も遺伝的変異にかぞえられる。非遺伝的変異には,ある1個体において形質が変化する場合と,個体間で形質に違いの現れる場合とがある。…
…遺伝学における突然変異体のことで,オランダの植物学者ド・フリースの《突然変異説Die Mutantionstheorie》(1901‐03)に初めて使われた用語。とくにSFでは根本的な変異をもった生物の意味に使われ,人間のミュータントがしばしば題材となり,この新人類に対して旧人類が不安や恐怖を抱くといった設定が多い。…
※「突然変異」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
夏の暑さに体が慣れること。数日から数十日間で起こる短期暑熱順化と、数年または数世代にかけて起こる長期暑熱順化とがある。→寒冷順化[補説]近年では、冷房設備の普及にともない短期暑熱順化が起こりにくくなっ...