デジタル大辞泉 「維管束」の意味・読み・例文・類語
いかん‐そく〔ヰクワン‐〕【維管束】
Sponserd by ![]()
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
植物の体内で物質の輸送の主要な通路となり、そのための特殊な組織を含む維管束組織を単にさすことが多いが、木部(もくぶ)と篩部(しぶ)とが組み合わさり1本の束となった状態のものをさし、管束ともいう。一般に第一次維管束組織において維管束の構造を示す。維管束は植物体内での場所によって特別の名称でよばれ、葉身(ようしん)の中にあるものは葉脈、茎の中にあるが葉や枝のほうへ分離していく途中のものをそれぞれ葉跡(ようせき)、枝跡(しせき)という。維管束は数種類の組織からなる複合組織で、厚くて硬い細胞壁をもつ細胞を含むことから、植物体を機械的に支持する役割もあわせもつ。植物解剖学では維管束に関する構造はとくに重要な形質として扱われる。植物の器官の中での維管束の配列や、1本の維管束の中での木部と篩部との配列は多様であるが、いくつかの型に分けられる。器官の中を貫いている維管束の太さや数はさまざまで、たとえば1本の茎の中に1本しかないものから、2万本もの維管束をもつものまである。
ドイツの植物学者ザックスは高等植物の組織を表皮系、基本系、維管束系の三つの組織系に大別した。維管束組織は基本組織に囲まれているが、器官の中に多数の維管束をもつ植物体全体では、維管束の互いの離合により、複雑な維管束系を形成する。維管束の生理的、系統的な重要性から、これをもつ植物群をまとめて維管束植物とよび、このなかにはシダ植物、裸子植物、被子植物が含まれる。コケ植物、藻類などの植物群は維管束をもたず、非維管束植物または無維管束植物とよばれる。しかし、コケ植物のなかには道束(どうそく)とよばれる水液の輸送に関与する構造をもつものがあり、褐藻類のコンブのように、被子植物の篩管と同様の組織を分化させているものもある。
[西野栄正]
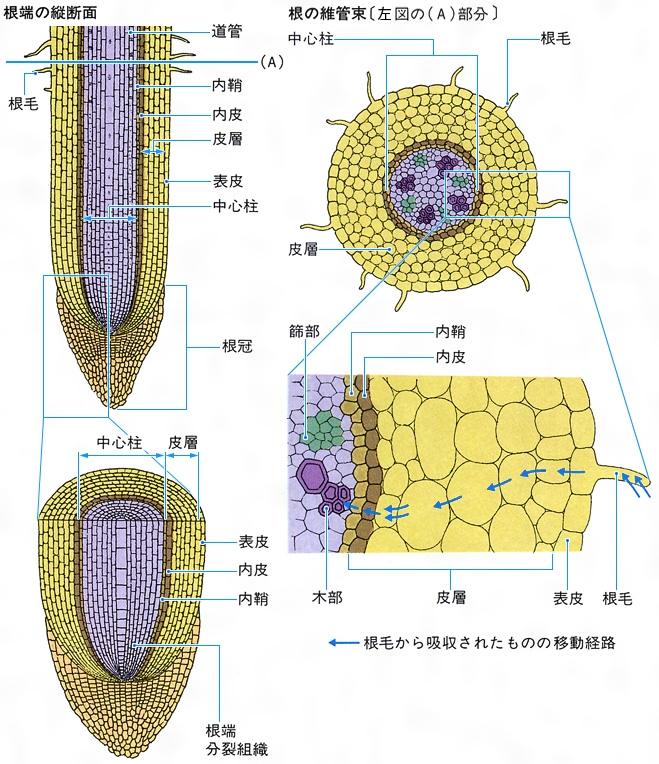
維管束は根から吸収された水液を輸送する木部と、葉などでつくられた養分や代謝産物の輸送および転送を行う篩部とからなる。木部は道管細胞、仮道管、繊維、柔細胞によって構成され、水液の通路は道管細胞や仮道管である。被子植物では道管細胞が連なって中空の管となった道管をもち、裸子植物やシダ植物は仮道管をもつ。しかし、被子植物でも葉脈の末端など細い維管束では仮道管となることが多く、裸子植物やシダ植物でも道管をもつものがある。篩部は篩管細胞、篩細胞、伴(ばん)細胞、繊維、柔細胞によって構成され、養分の通路は篩管細胞や篩細胞である。被子植物は篩管細胞が連なった篩管と、篩管細胞に隣接した伴細胞をもつ。裸子植物とシダ植物は篩細胞をもつ。
[西野栄正]
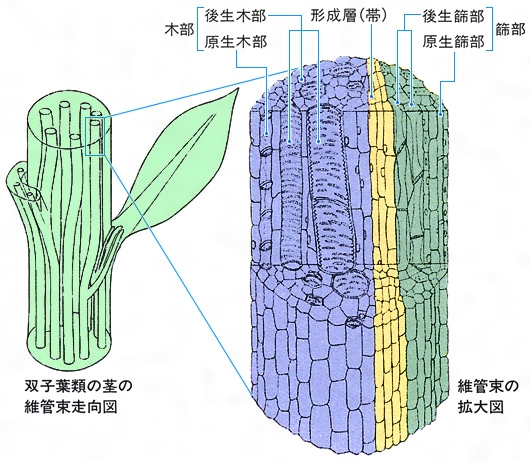
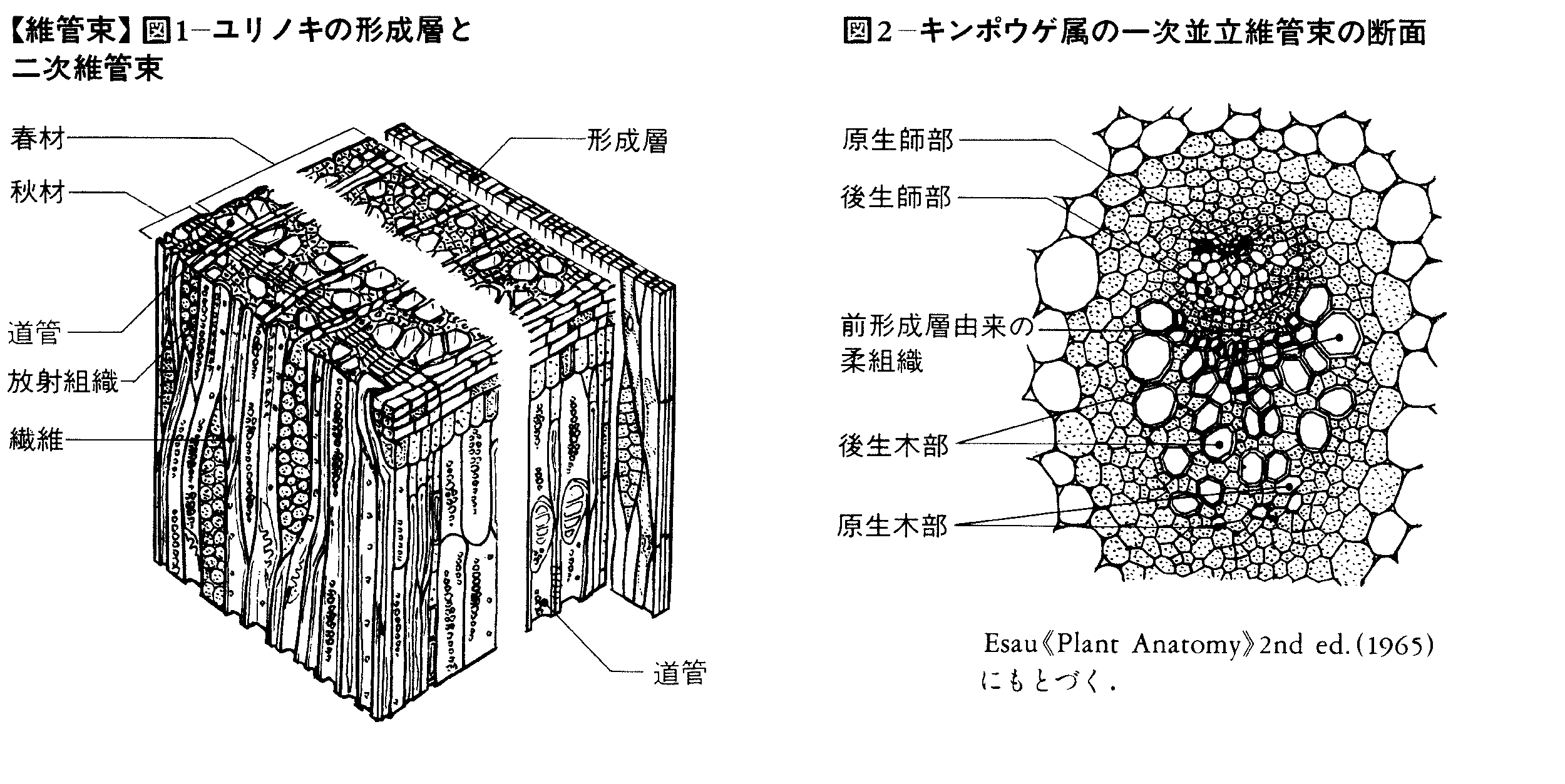
維管束植物の茎や根が軸方向へ成長していくのは、それらの先端にある頂端分裂組織で増殖した細胞が、茎や根の組織に加わることによっておこる。未分化な細胞はしだいに原表皮、基本分裂組織、前形成層に分化、さらにそれぞれ表皮組織、基本組織、維管束組織へと成熟する。葉でも葉原基(ようげんき)の成長の間に同様に各組織が形成される。このように発生したのが第一次維管束組織で、第一次木部と第一次篩部とからなり、器官の中の1本の維管束となる場合や、基本組織によって分断された多数の維管束となる場合がある。前形成層からの木部と篩部の分化はそれぞれ一定の方向におこり、木部の中ではまず原生木部が、ついで後生木部が分化し、仮道管や道管細胞の構造が異なる。篩部も原生篩部と後生篩部とに分けられる。前形成層の細胞が維管束組織に分化したあと、木部と篩部の境界部で維管束内形成層が分化する開放維管束と、それが分化しない閉鎖維管束とがあり、前者はおもに裸子植物と双子葉類の茎や根に、後者はおもに双子葉類の葉とシダ植物や単子葉類の茎や根にみられる。形成層が維管束間の部分にも分化すると環状につながり、その細胞が接線分裂を繰り返して増殖し、内側には第二次木部、外側には第二次篩部という第二次維管束組織を形成する。維管束形成層が第一次維管束組織とは離れた場所で分化する場合もある。
[西野栄正]
維管束は木部と篩部の配置によって、並立(へいりつ)、複並立、包囲の各維管束に分けられる。並立維管束は裸子植物と被子植物の茎や葉にもっとも普通で、木部と篩部とが一面で接し、茎では内側に木部、外側に篩部が、葉では上側に木部、下側に篩部が位置する。ウリ科やナス科植物などの茎には木部の内外に篩部のある複並立維管束があるが、内側の篩部が木部とかなり離れている場合もある。包囲維管束は木部と篩部のうち、一方が他方に囲まれた構造をもつ。木部が篩部に囲まれた外篩(がいし)包囲維管束はシダ植物の茎や葉に多くみられ、被子植物にもまれにみられる。篩部が木部に囲まれる外木(がいもく)包囲維管束は単子葉類の地下茎でよくみられ、第二次維管束組織をつくった単子葉類の地上茎でもみられる。すべての根では木部と篩部とが交互に、それぞれ独立した維管束として環状に配列しているが、この配列から、それぞれを放射維管束とよぶことがある。
[西野栄正]
茎の中の維管束は分類群によって異なる配列を示す。双子葉類や裸子植物では、普通、茎の周辺部に環状に並び、単子葉類では不規則な配列を示す。葉跡が髄や皮層の中を長く迂回(うかい)する場合には、とくにこれらをそれぞれ髄走条(ずいそうじょう)、皮層条とよぶ。シダ植物では1本の維管束をもつものから多数の維管束をもつものまであって多様で、またすべての根のように木部と篩部とが交互に並ぶものもある。茎と根とで維管束の配列が異なる場合は、茎と根の移行部で配列が変化し、芽生えの胚軸(はいじく)を中心とした部分がそこにあたる。維管束のとくに軸方向の推移を維管束走向という。茎と根の維管束を含む領域を中心柱として区分し、維管束の種類や配列と相互の関係を論じるのが中心柱説である。葉身は普通扁平(へんぺい)で、維管束も平面的に配列するが、主要な維管束の配列から二又(ふたまた)脈、網状脈、平行脈に大別される。葉柄(ようへい)内でも平面的な配列を示すが、茎の場合のように立体的で複雑な配列を示す場合も少なくない。
[西野栄正]
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
シダ植物および種子植物の茎・根・葉などの器官の内部を貫く条束状の組織系。これらの植物はそのため維管束植物と呼ばれ,維管束をもたないコケ植物や藻類などと区別される。陸上植物は水中生活をしていた緑藻類のあるものが地上に侵出して進化したものであると一般に考えられ,最も古い維管束植物は4億年前の地層から出土している。維管束の構造と機能はその生活様式や環境と密接な関係がある。つまり植物体の一部が地中に,他の部分が空中にあるという体制を維持するためには,機械的な支持構造と,水分や体内物質が移動して乾燥から身を守り必要な養分を供給する機構が必要であって,維管束はこれらの機能を同時に果たす組織であるといえる。維管束は一般に配偶体には分化しないが,マツバランの配偶体に不連続な維管束が観察されることがあり,配偶体世代と胞子体世代の世代交代の関係を考える上で興味深い現象である。維管束に似た構造が褐藻類やコケ植物の一部にあることが知られているが,前者は維管束植物とは別系統であり,後者は配偶体に維管束が分化するものである。ザクスJ.Sachsは維管束を維管束系として表皮系,基本組織系から区別した。この分け方は植物群や器官の違いをこえて一般的に利用できる。一方ファン・ティガンP.E.L.van Tieghemは維管束を表皮・皮層の内側にある中心柱として定義した。この分け方は系統を考えるときには都合がよい。生理的性質を重視して維管束を通道組織とする分け方もあって,研究目的によって違った区分法が用いられている。
維管束は木部と師部からなり,内皮によって囲まれる場合もあればそうでない場合もある。木部は道管・仮道管・木部柔組織・木部繊維から,師部は師管・師部柔組織・師部繊維などからできている。木部は水分が,師部は養分が移動する通路であるが,その運搬機構については詳しくはわかっていない。木部と師部の配列は植物群や器官によってさまざまであり,1本の維管束で木部が内側に師部が外側にある並立維管束(被子植物と裸子植物の茎と葉),木部の内側にも師部がある複並立(両立)維管束(ウリ科など),一方が他方に囲まれる包囲維管束(外師包囲維管束はシダ植物の茎と葉にふつう),木部と師部が交互にならび環状をなす放射維管束(維管束植物の根)などがある。一次維管束は茎頂・根端および葉縁の各分裂組織の近くで分化した前形成層から分化する。分化は一斉にすすむのではなく維管束の部位によって異なり,伸長生長の盛んな頂端部でまず分化するものを原生木部,原生師部,あとから分化するものを後生木部,後生師部という。一次維管束の大部分は後生木部・師部が占める。1本の維管束における原生木部と後生木部の位置関係はさまざまであり,典型的には原生木部が木部の外端にある外原生(維管束植物の根,小葉植物の茎など),中央にある中原生(シダ類の茎など),内側にある内原生(被子植物,裸子植物の茎など)があって,原生中心柱において原生木部が中心にある場合をとくに心原生(化石のライニアなど)という。この位置関係は植物群や器官によってだいたいきまっていて,系統を考える上で重視されてきている。とくに維管束植物が起源したと推定される下部デボン紀(約千億年前)の植物を,木部の発生様式と胞子囊の特徴によって2系統群に大別する古植物学的知見(たとえばバンクスJ.Banks)は注目に値する。
茎の一次維管束の前段階である前形成層の分化の要因については,茎頂がその分化を誘導することがいろいろな実験によって確かめられている。また前形成層分化と葉原基発生の因果関係は,茎の維管束が葉の維管束の集りであるかどうかという古くから議論されてきた問題と関連し,実験形態学的研究がすすめられているが,両者は相補的な関係にあるといわれている。一次維管束のほかに形成層から由来する二次維管束がある。形成層から内側へ二次木部(材),外側へ二次師部が分化する。それらをつくる細胞,組織は放射組織を除いて一次維管束の場合と同じで,形成層の紡錘体形始原細胞からできる。細胞配列は一次維管束の場合と比べて規則的である。二次維管束にはこれらの縦方向の組織のほかに,放射組織始原細胞に由来する水平放射方向に配列する放射組織がある。二次維管束は系統的には3.7億年前の中部デボン紀には起源しており,裸子植物の祖先と考えられる原裸子植物が裸子植物の材によく似た二次木部をもっていたことが知られている。現生植物ではふつう裸子植物と被子植物,とくに双子葉植物にみられる。現生シダ植物ではハナヤスリ科とミズニラ科に例外的にみられるが,化石植物ではヒカゲノカズラ植物やトクサ植物などいろいろな植物群にあったことがわかっている。
執筆者:加藤 雅啓
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
…篩管とも書き,〈ふるいかん〉とも読む。被子植物の維管束の中にあって,師部を構成する細胞の一つで,体内でつくられた養分の通道の機能を果たす。縦に細長い形をし,両端面は穴のあいた板状の師板sieve plateとなり,互いに連結する。…
※「維管束」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
福岡県福岡市博多区の櫛田神社の夏祭り。壮麗な山笠で知られる。今日,山笠には飾り山笠と舁き山笠(かきやまがさ)の 2種類がある。明治時代に電線が架設されて以降,物語場面の人形などを飾りつけた高さ 15m...