デジタル大辞泉 「自律神経系」の意味・読み・例文・類語
じりつしんけい‐けい【自律神経系】
Sponsored by ![]()
Sponsored by ![]()
心臓と全身の血管,皮膚の立毛筋と汗腺,さらには内臓諸器官に含まれる腺細胞ならびに平滑筋細胞に分布するような神経系を総称していう。かつて,脳の支配から比較的独立して働くと考えられたため,この名がつけられた。ラングリーJ.N.Langleyの命名(1905)による。交感神経系と副交感神経系の2系統が含まれるが,いずれの系統の場合も,脳と脊髄(すなわち中枢神経系)から遠ざかる向きに神経刺激を伝えるような遠心性要素と,その反対の向きに神経刺激を伝えるような求心性(または知覚性)要素とが存在するのであるが,求心性要素を自律神経からまったく除外することもあるので注意を要する。自律神経系は,全身の骨格筋ならびに皮膚に分布する体性神経系としばしば対比され,後者が目に見えるような身体動作もしくは随意運動に強く関与するため動物性神経系とも称されるのに対して,植物性神経系と称されることがある。すなわち,体性神経系が生体の外界への反応を媒介する性格を示す一方で,自律神経系は生体の内部環境を整えるものであるとの見方ができる。自律神経の遠心性要素は,節前ニューロンと節後ニューロンとの2者で構成される。
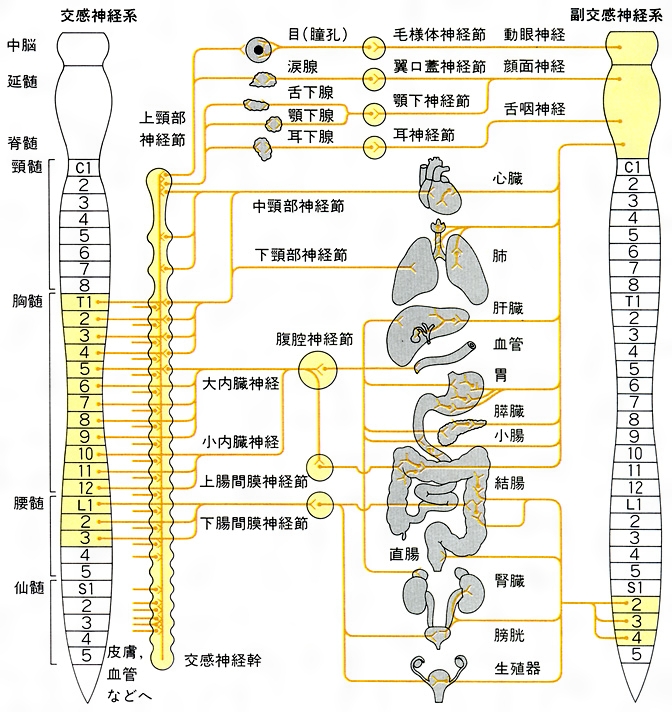
交感神経系の節前ニューロンをなすものは,その細胞体が第1胸髄から第3腰髄までの髄節の高さにおける脊髄内に存在し,神経突起が脊髄神経の前根から白交通枝を経て交感神経幹に入り,幹神経節または椎前神経節内の交感神経系節後ニューロンの樹状突起または細胞体とシナプスを形成するような神経細胞である。幹神経節は,頸部から尾骨部にまで至る範囲の高さで脊髄の両側にとびとびに存在し,1側当り頸部で3個,胸部で10~12個,腰部で4~5個,仙骨部で3~5個を数えるが,これらを縦に1本につなぐものが交感神経幹にほかならない。椎前神経節(脊椎の前面に位置するためにその名がある)と幹神経節を合わせたものが交感神経節である。節前ニューロンの神経突起は髄鞘で覆われ,いわゆる交感神経系節前繊維の軸索をなす。このものの長さは成人では数cmから十数cmであり,脊髄のある高さの髄節部分から出た節前繊維が同じ高さで交感神経幹内にいったん進入したのちに,さまざまな距離を走行することを示している。幹神経節を通り過ぎて椎前神経節に向かう節前繊維は当然のことながら交感神経幹を離れるのであるが,その際に相集まる傾向を示して大内臓神経,小内臓神経,仙骨内臓神経などを形成する。1本の節前繊維は終末部で多数の枝分れを示し,交感神経節内における多数の節後ニューロンに対してシナプス伝達を行う。
交感神経系の節前ニューロンはアセチルコリン作動性であり,節後ニューロンはノルアドレナリン作動性である。したがって交感神経節内でシナプス伝達がなされるとき,アセチルコリンが節前繊維末端から放出されて節後ニューロンの興奮が引き起こされるのであるが,その結果としてのノルアドレナリン放出も少し遅れて起こり,これが節前繊維の活動を抑制させることが知られている。
交感神経節内に位置する交感神経系の節後ニューロン細胞体のおのおのから1本の軸索状神経突起(無髄繊維を形成することが多く,長さ十数cmから1m余にも及ぶ)が出て体壁または内臓に向かう。このうち,体壁に向かうものは幹神経節内に起始細胞体をもつ場合が多く,(1)交感神経幹と脊髄神経との間の灰白交通枝を通って脊髄神経の中に入り,頸部以下の全身皮膚の立毛筋あるいは汗腺に達するもの,(2)交感神経幹を離れるや否や最寄りの動脈壁に達し,動脈の平滑筋に対する支配を行いながら動脈分布上の最終地点としての頭部の皮膚と内臓(眼球など),あるいは頸胸部内臓に達するもの,(3)交感神経幹を離れてから,心臓に向かう上,中,下の心臓神経あるいは胸心臓神経をつくって心臓に直行するもの,のいずれかとなる。
一方,椎前神経節内の起始細胞体から出る節後繊維は腹部内臓に向かうものが多く,その際に内臓に分布する動脈壁を通路として利用する。交感神経系の節後繊維の末端からは神経伝達物質としてのノルアドレナリンが放出されるが,これにより皮膚の立毛筋はいっせいに収縮を起こして,いわゆる総毛立つ状態となり,また少なくとも皮膚の血管は内腔の狭小化を示すために皮膚の蒼白化および血圧上昇を招く。これらの全身的な変化に随伴する現象として,散瞳(瞳孔散大筋の収縮による),眼裂開大(上瞼板筋の収縮による),心悸亢進(歩調取り特殊心筋の活動が高まることによる),細気管支の拡張(壁の平滑筋が弛緩することによる),心筋血流量の増大(冠動脈壁平滑筋の弛緩による),腹部内臓血流量の減少(内臓に分布する動脈壁平滑筋の収縮による),などをあげることができる。交感神経系の遠心性要素の活動は副腎髄質からのアドレナリンの内分泌をも亢進させるが,これは副腎髄質のクロム親和細胞(アドレナリンの生合成,貯蔵,放出を行う)に対して交感神経系節前繊維の興奮がシナプスを介し伝えられるためである。アドレナリンは交感神経系節後ニューロンの伝達物質であるノルアドレナリンに近い化学構造と生体作用を示す物質であり,これが副腎髄質から内分泌されて全身の血液中に運ばれるという現象の意義ははなはだ大であるといえる。
交感神経系の求心性要素となるものは第1胸髄から第3腰髄までの高さの脊髄神経節内に細胞体を有する第1次知覚性ニューロンであり,その細胞体から末梢側に伸びる樹状突起(軸索状の形態を示す)が交感神経系遠心性要素に伴行しながら血管壁や内臓諸器官にまで至り,同じ細胞体から出て中枢側に伸びる神経突起(これも軸索状の形態を示す)が脊髄内の後角に達して,そこで第2次知覚性ニューロンに対するシナプスを形成する。内臓からの痛覚を伝えるものは主としてこのような交感神経系の求心性要素であるとされる。ときには内臓に病変が生じているのにもかかわらず,その内臓の痛みをほとんど感じないことがある。しかし,その場合でも当該内臓に分布する交感神経系求心性要素は脊髄後角に向かう異常神経刺激を伝えているのであり,これが脊髄神経節レベルあるいは脊髄神経後根から脊髄内の後角に至るレベルにおいて隣接する他の第1次知覚性ニューロンへと伝染することがある。このような神経刺激伝染を受けたニューロンが皮膚からの痛覚を伝えるものであれば,そのニューロンが支配する健康な皮膚領域からの痛みが意識にのぼる結果となる。特定の内臓疾患の病変時に特定部位の健康な皮膚に痛みが放散する現象,すなわち関連痛がこのようにして発生する。
副交感神経系の遠心性要素は,交感神経系のそれと同じく,節前ニューロンと節後ニューロンとの2者で構成されるが,節前繊維の長さに比較し節後繊維が極端に短いという点が特異である。すなわち,副交感神経系節前ニューロンの細胞体は,脳幹内における動眼神経副核,上および下唾液核,迷走神経背側核のいずれか,あるいは第2~4仙髄の高さの脊髄の内部に存在し,そこから出る神経突起が成人では約10cmから数十cmに至る長さの節前繊維をなす。一方,副交感神経系節後ニューロンの細胞体は,そのニューロンの支配下にある内臓諸器官の壁内もしくは近接部位に位置を占める副交感神経節の内部などに存在する関係上,節後繊維の長さは数mmからたかだか数cmでしかない。概算すれば,交感神経系における節前繊維と節後繊維の長さの比がおよそ1対10であるのに対して副交感神経系における対応値は10対1となり,きわめて対照的であることが明らかとなろう。1対10という比は末広がり型の,また10対1という比は末つぼみ型の,節前および節後ニューロンの配列様式をそれぞれ意味する。交感神経系の遠心性作用が広範な身体部位を巻き込む,すなわち交感的連鎖反応を引き起こす性質を示すのに対して副交感神経系の遠心性作用の効果が限局的であるといわれるゆえんである。
副交感神経系の節前ニューロンと節後ニューロンは,ともにアセチルコリンを神経伝達物質としている。動眼神経内を走行する副交感神経系節前繊維(動眼神経副核を中心とした脳幹内領域から出る)は毛様体神経節に至り,同神経節内の節後ニューロン細胞体に対してシナプスを形成する。この細胞体から出る節後繊維(長さ約2cm)が眼球内の瞳孔括約筋および毛様体筋に分布する。これらの筋はいずれも平滑筋繊維のみ(ただし鳥類と爬虫類では横紋筋繊維のみ)で構成されるが,副交感神経系の神経刺激により収縮して瞳孔の縮小と水晶体曲率の増大を起こす。
顔面神経内を走行する副交感神経系節前繊維(上唾液核をなす神経細胞体から出る)には,翼口蓋神経節に至るものと顎下神経節に至るものとがあるが,いずれにせよ,これら神経節内で節後ニューロンに対するシナプスを形成する。翼口蓋神経節からの節後繊維(長さ3~5cm)は,眼窩(がんか)に存在する涙腺に進入し涙の分泌支配を行う。また,顎下神経節からの節後繊維(長さ1~3cm)は,顎下腺または舌下腺に進入して唾液分泌を支配する。舌咽神経内を走行する副交感神経系節前繊維(下唾液核をなす神経細胞体から出る)は,耳神経節に至り,同神経節内の節後ニューロンに対するシナプスを形成する。耳神経節からの節後繊維(長さ2~4cm)は,耳下腺に進入し唾液分泌を支配する。副交感神経系の神経活動により涙および唾液の分泌は亢進する。迷走神経内を走行する副交感神経系節前繊維(迷走神経背側核をなす神経細胞体から出る)は,咽頭から横行結腸までの消化管のすべてと,これに付属する肝臓,膵臓および胆囊,咽頭から肺に至る呼吸器のすべて,心臓などにまで達するような,さまざまの長さのものであり,これらの臓器の壁内に散在する節後ニューロンの細胞体あるいは樹状突起に対するシナプスを形成する。臓器壁内の節後ニューロンの神経突起がつくる節後繊維(長さ数mmから数cm程度)は,臓器内における平滑筋または心筋の運動,さらには腺細胞の分泌活動を支配する。副交感神経系の神経興奮により,これらの内臓の平滑筋および腺細胞の活動が促進されるが,心筋の活動のみは抑制される。以上述べた副交感神経系遠心性要素には,節前繊維が脳神経内を走行する(あるいは節前ニューロン細胞体が脳内に存在する)という共通点がみられるため,これらを副交感神経系の脳部と呼ぶことがある。
脊髄の下端に近い,第2~4仙髄内に細胞体があり,同じ高さの脊髄神経前根を経由して骨盤内臓神経に入るような節前繊維をもつような遠心性要素も存在し,これは副交感神経系の仙骨部の中に含まれるものである。骨盤内臓神経は,横行結腸から直腸に至る消化管,膀胱,卵巣,卵管,子宮,精巣,精管,精囊,前立腺などに向かう細枝に分かれるが,これらの細枝は骨盤内で吻合(ふんごう)と枝分れを繰り返して骨盤神経叢と呼ばれる構造をつくる。この神経叢内には交感神経系および副交感神経系の節後ニューロン細胞体も混在している。骨盤内臓神経に含まれてこの神経叢にまで達した副交感神経性節前繊維の一部は,神経叢内で副交感神経性節後ニューロンに対するシナプスを形成するが,残りは上記の内臓壁にまで達して,そこではじめて壁内の副交感神経系節後ニューロンに対するシナプスを形成する。骨盤神経叢内から発する副交感神経系節後繊維には,短い走行ののちに付近の骨盤内臓器に進入するものと,比較的長い(数cm)もので動脈壁にまつわりつきながら会陰部の海綿体組織にまで達するものとがある。後者の活動が陰茎,陰核,前庭球の勃起を生じさせる(海綿体組織に注ぐ動脈の壁の平滑筋を弛緩させて動脈血流量を増大させることによる)ことから,骨盤内臓神経には勃起神経の別名がある。内臓壁内から発する仙骨部副交感神経系の節後繊維はきわめて短いものと考えられている。
副交感神経系の求心性要素をなすものは,少なくとも舌咽神経,迷走神経,および骨盤内臓神経に含まれている。舌咽神経に含まれるものは,舌咽神経の下神経節の内部に細胞体があり,末梢側突起(知覚性軸索)を頸動脈洞と頸動脈小体にまで送り,中枢側突起を延髄内の迷走神経背側核にまで届かせているような知覚性ニューロンである。このニューロンは,頸動脈洞における血管圧変化を感受し,頸動脈小体では血流中の化学的成分(酸素分圧,水素イオン濃度)変化を感受する。迷走神経に含まれる副交感神経系求心性要素をなすものは,迷走神経の下神経節の内部に細胞体をもち,末梢側突起(知覚性軸索)を大動脈弓,胸腹部内臓などに送り,中枢側突起を延髄内の迷走神経背側核にまで届かせているような知覚性ニューロンである。このニューロンは,大動脈弓壁の圧変化受容と大動脈小体での血液化学成分変化の受容とを行うほか,胸腹部(ただし仙骨部副交感神経系の分布範囲を除く)の内臓からの機械的刺激および化学的刺激をも延髄にまで伝える。脳神経のうちの舌咽および迷走神経に含まれるこれらの知覚性ニューロンが伝える神経刺激は,無意識のうちに営まれる内臓・血管反射に役だつものである。骨盤内臓神経に含まれる副交感神経系求心性要素をなすものは,脊髄の第2~4仙髄の高さの脊髄神経節の内部に細胞体を有し,末梢側突起(知覚性軸索)を骨盤内の臓器にまで伸ばし,中枢側突起を仙髄に進入させているような知覚性ニューロンである。この種のニューロンが伝える骨盤内臓器からの刺激には,膀胱壁伸展度を伝える機械刺激(尿意),妊娠子宮壁の収縮度を伝える機械刺激(陣痛)など,意識にのぼるものが少なくない。このような求心性刺激に応じた内臓反射も引き起こされるのであるが,その際に正常状態下では,ある程度までは内臓反射(排尿や分娩)の発現が大脳皮質による意志の統制下に置かれ,場所や状況が適当であるか否かの判断がからむ点が特異である。
一般に,交感神経系の神経活動は生体エネルギーを外界に向け放散させる効果をもたらし,副交感神経系の神経活動は生体内にエネルギーを蓄える効果をもたらすといえる。両系は多くの場合に一つの器官内に共存して互いに拮抗的な作用を示す。たとえば心臓に対して,交感神経系の神経活動は拍動リズムを速め,かつ毎回の拍出量を増大させるが,副交感神経系の神経活動はまったく反対の向きの,マイナス効果をもたらす。また,消化器官に対しては,交感神経系の神経活動が胃腸管運動を抑制し消化酵素や胆汁の分泌を低下させる一方で,副交感神経系の神経活動は正反対のプラス効果をもたらす。これらの例におけるような,いわばプラスの手綱とマイナスの手綱の役割をそれぞれ果たす2系の自律神経系遠心性要素が単一臓器の支配にあずかる現象は,下等動物では比較的まれとなる(魚類では心臓も消化器官も副交感神経系による1本の手綱による活動調節を受ける)。
支配臓器内に進入した自律神経系の遠心性要素は,最終的には無髄性の節後繊維となって臓器内の平滑筋細胞,心筋細胞,腺細胞などの周囲をとりまく網目を形成するが,その場合に交感神経系節後ニューロン(ノルアドレナリン作動性)の軸索と副交感神経系節後ニューロン(アセチルコリン作動性)の軸索が共通のシュワン鞘に覆われて,互いに接しながら走行する。そのために1系の節後ニューロンの活動が他系の節後ニューロンに対しても,その活動を促すような影響を与える。すなわち,終末網目をつくる節後ニューロン軸索は,いずれもその内部に伝達物質を含有するシナプス小胞を多数備えており,伝達物質の軸索からの放出が随所で起こるものと考えられる。
このようにして放出された神経伝達物質は,臓器内の細胞間隙をかなりの距離にわたり拡散し,不特定の数の効果器細胞(平滑筋細胞,心筋細胞,腺細胞などの自律神経支配を受ける細胞をいう)の表面にまで達するのが普通である。しかし,臓器によっては終末網目から節後ニューロン軸索がさらに伸び出して効果器細胞の表面に密着する場合がある。涙腺の腺細胞および筋上皮細胞,瞳孔の散大筋と括約筋をなす平滑筋細胞,精管壁の平滑筋細胞などがそのような密着型の自律神経支配を受け,神経伝達が効果器細胞に対して,より強力に行われるしくみがそこにみられる。事実,前記のいわば遠隔型の神経伝達を受ける平滑筋細胞群(血管,胃腸管,尿管,膀胱,子宮などの壁に含まれているもの)や心筋は,神経に対する依存度が比較的低く,自動能を有しており,神経終末から放出されるノルアドレナリンやアセチルコリンは,そのような細胞の活動レベルを設定するような,一種の組織ホルモンとして作用するという面がみられる。これに対して密着型の神経支配を受ける平滑筋細胞群に共通していえることは,それが自動能を示さず,収縮を行う場合に自律神経系からの神経刺激にまったく依存するということである。
自律神経系の遠心性要素がアセチルコリンを伝達物質とする節前ニューロンとノルアドレナリンを伝達物質とする節後ニューロンによる組合せ(交感神経系),またはいずれもがアセチルコリンを伝達物質とするような節前および節後ニューロンの組合せ(副交感神経系)で構成されるとの考え方は,古典的なものではあるが今日でもなお大筋において正しいものと信じられる。しかし,近年の研究の進歩により,自律神経系の遠心性要素の中にはγ-アミノ酪酸,セトロニン,ATP,あるいはペプチド類を伝達物質とするニューロンが混在する可能性がしだいに明確になりつつあるのが現状である。これらのニューロンの多くは胃腸管の壁内に節後要素として存在するらしい。
また自律神経系の知覚性要素のうちで,少なくとも血管壁あるいは内臓からの痛みを伝えるニューロンは,P物質と呼ばれるペプチドを伝達物質としている可能性が強い。このようなニューロンの知覚性軸索の末梢端は,自律神経系の遠心性要素とともに終末網を形成しながら,生体の内部環境モニターとしての役割を果たしている。
→シナプス →神経系 →ニューロン
執筆者:山内 昭雄
自律神経系の最高中枢は間脳の視床下部にあるが,そこにおける自律性機能の中枢のあり方については諸説がみられた。視床下部の前部が副交感神経系で,後部が交感神経系であるという意見や,視床下部には交感神経系の中枢だけが存在し,副交感神経系は視床下部より前方にあるという説などがあった。しかし近年,黒津敏行が広範囲の実験的,組織学的な研究結果に基づいて,第三脳室から外方へ順次にa,b,cの3細胞層があり,bの部分が交感神経系の中枢,aとcとが副交感神経系の中枢であることを明らかにした。視床下部から下行して,下位の自律神経系の中枢に連絡する繊維の走行については,まだ詳細が不明な点もあるが,現在新しい方法で追究されつつある。
脊椎動物の各綱を通して自律神経系は基本的には同じ様相を示す。系の本幹ははしご状で,一見昆虫などの神経系に似ている。自律神経系は臓器を支配し,神経系のなかでも原始的なものである。自律神経系が機能的に交感神経と副交感神経とに分かれるのは系統発生上新しい。ヤツメウナギでは,消化管はほとんど副交感神経に属する迷走神経で支配されている。脊椎動物の自律神経の遠心性繊維は脊髄の前根を通過する。サメ類では神経節を連絡する索は形成されていないが,硬骨魚類になると交感神経索が出現する。完全に陸上生活に成功した爬虫類以後の動物では,空気中の生活条件に対する広範囲の順応,つまり体温調節などのために汗腺,毛などが生じ,摂餌のため大きな唾液腺が発達し,さらに体内受精,母体内での胚の発生に伴って,内部生殖器が発達する。その結果,頭部と骨盤部に分布する自律神経系が発達する。そのうえ運動性の増大によって,交感神経系が優位になる。すでにスッポンの自律神経系は基本的にはヒトによく似ている。
執筆者:正井 秀夫
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponsored by ![]()
脳や脊髄(せきずい)から発して身体の隅々にまで達する末梢(まっしょう)神経のうち、呼吸、循環、消化のような生命の維持に直接関係する諸臓器の機能、すなわち植物性機能に関係する神経を総称していう。自律神経の活動は意志とはまったく無関係に、もっぱら反射によって調節され、昼夜を分かたず諸臓器の機能を調整している。自律神経系は交感神経系および副交感神経系の2系統からなる。ヒトの場合、交感神経は脊髄の胸および腰の部分(胸髄・腰髄)から発し、副交感神経は中脳・橋(きょう)・延髄の脳神経核から発する迷走神経と、脊髄の最下部、すなわち仙髄から発する骨盤神経からなる。これらの自律神経末梢路に対する上位中枢は延髄および視床下部であり、ここで交感・副交感両神経系の統合が行われている。さらに自律神経系は大脳皮質、とくに辺縁葉とも密接な関係にあり、その影響を強く受けている。
[真島英信]
自律神経の末端は主として心臓、その他の内臓・血管などの平滑筋、各種の外分泌腺(がいぶんぴつせん)・内分泌腺に分布し、それらの活動の程度を調節している。交感神経および副交感神経は、多くの場合、一つの器官にそれぞれが独立して神経線維を送っている(二重支配という)。しかもこの二つの神経系の作用は互いに反対であり、ある器官に対して一方がその活動を促進する場合は、他方は逆に抑制するように働く。つまり交感・副交感両神経系は相互に拮抗(きっこう)的な作用を及ぼしている(拮抗支配という。ただし、血管の場合は例外で、多くの血管には交感神経しかきていない)。さらに、ある器官に対する交感神経の活動が亢進(こうしん)しているときには副交感神経の活動は抑制され、逆に副交感神経が活発に活動しているときには交感神経の活動は抑制される(これを相反支配という)。また、両神経系とくに交感神経は、つねにある程度の興奮状態を持続しており、支配する器官に対して刺激(インパルス)を送り続けている。これを持続神経支配という。したがって両神経系の及ぼす刺激効果がちょうどつり合ったところに、その器官の興奮性が維持されているといえる。
[真島英信]
交感神経の作用は精神的に興奮したときや運動時に亢進する。このことは、われわれが恐ろしい場面から逃げ出そうとするときなどに経験するさまざまな身体的変化を考えてみれば比較的容易に理解することができる。このような場面にあっては、交感神経が興奮することによって、目は見開かれ(瞳孔(どうこう)散大)、顔面は蒼白(そうはく)となり(顔面の血管の収縮)、さらに精神性発汗が盛んになり、いわゆる「手に汗を握る」状態となる。また、気管支平滑筋が弛緩(しかん)するため、気管支は拡張し、呼吸は促進される。さらに心臓の活動(心活動)も促進されて収縮が強くなり、かつ心拍数が増加して胸の鼓動を感じるようになる(いわゆる胸がどきどきする状態)。以上が交感神経の作用が亢進したときの例であるが、このとき、激しい身体の運動には直接関係しない消化管の蠕動(ぜんどう)運動や消化液の分泌、および泌尿生殖器系の活動は逆に抑制される。なお、動物が精神的に興奮したときには立毛筋が収縮して体毛が逆立つ現象がみられる。これは、闘争または逃避のための準備状態が交感神経の活動によってつくられたといえる。また、交感神経は副腎(ふくじん)髄質を刺激してアドレナリンというホルモンの分泌を促進させる。アドレナリンは交感神経の作用とよく似た効果を諸臓器に引き起こし、交感神経興奮の効果を増幅させる働きをもっている。
[真島英信]
副交感神経は、一般にわれわれが安静にし、ゆったりとくつろいでいるとき、すなわち休息中や睡眠時にその活動が亢進する。その結果として瞳孔の縮小、気管支平滑筋の収縮が生じるほか、心臓の拍動が抑制されて1回心拍出量の減少、心拍数の減少、血圧の低下がおこる。また、消化管の蠕動運動や消化液の分泌、泌尿生殖器系の活動は促進される。このようにみると、副交感神経はその活動によって消耗した身体の回復に適した条件を整える働きをもっているといえる。ただし、すでに述べたように、このような状態においても交感神経系は絶えず活動を続けており、諸臓器に刺激を送っている。
[真島英信]
電気的信号である興奮が神経線維を伝わってその末端に到達すると、そこから化学物質が放出される。この物質は化学伝達物質とよばれ、臓器の細胞に作用してその機能を変化させるものである。交感神経と副交感神経の作用が前に述べたようにまったく異なっているのは、この化学伝達物質が両者間で異なっているからにほかならない。交感神経線維末端から放出される化学伝達物質はノルアドレナリンであり、副交感神経線維末端から放出されるのはアセチルコリンである。これらの化学伝達物質は神経線維末端から放出され、各臓器に作用を及ぼしたのち、血液中の酵素の働きによって速やかに分解される。諸臓器に対する交感・副交感神経興奮の影響は、それぞれの末端から放出される化学伝達物質の多寡によって決まってくる。したがって、神経を刺激しなくても、直接、臓器にこれらの化学伝達物質を作用させることによって同様の効果を得ることが可能となる。たとえばノルアドレナリンを静脈内に注射すると、心活動の促進による心拍数増加、動脈収縮による血圧の上昇、瞳孔の散大など、交感神経が興奮したときと同様の変化を生じさせることができる。
[真島英信]
脊椎(せきつい)動物の自律神経系にもヒトと同様に2系統があり、それぞれが拮抗的に作用することは同じであるが、ここでは、神経繊維(医学では線維と表記する)の連絡様式についてさらに詳しく述べる。
自律神経系においては、中枢神経系内の神経細胞から出た有髄の神経繊維(節前繊維)は、その支配する器官に達する前に、神経節または神経集網においてもう一つのニューロン(神経単位)に中継される。このニューロンの繊維は一般に無髄で、節後繊維とよばれる。交感神経の節後繊維は、汗腺を支配するものを除き、一般にアドレナリン作動性である。これに対し、副交感神経系の節後繊維、および交感・副交感神経系の節前繊維はコリン作動性である。以上のような自律神経系の基本的な性質は、ネコやイヌなどヒトを含む哺乳(ほにゅう)類を中心として調べられたものである。
哺乳類の発達した自律神経系は、脊椎動物の祖先に近いと考えられる半索動物や原索動物の内臓、とくに咽頭(いんとう)や腸とその付属器官の働きを調整している神経集網にその原型をみることができる。扁形(へんけい)動物や円形動物などの消化系にも神経集網があり、内臓の機能を制御する神経系は広く三胚葉(はいよう)性動物(扁形動物以上のすべての動物)に存在するものと考えられるが、研究はほとんど行われていない。原索動物のナメクジウオでは、中枢から出た節前繊維に相当する繊維は、直接、末梢の神経集網に達している。交感神経幹はなく、皮膚、血管系ともに中枢からの神経支配を受けていない。円口類では副交感神経は迷走神経に限られている。サメ類の血管は、中枢からの神経支配を受けており、中枢に近い神経節でニューロンの交代が行われる。魚類の自律神経系の発達は、グループによる変異が大きく、自律神経系の基本的な系が確立したのは、両生類以上の脊椎動物であるとされている。
[村上 彰]
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponsored by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponsored by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponsored by ![]()
Sponsored by ![]()
…一般にこれら運動ニューロンの化学伝達物質はアセチルコリンである。 一方,平滑筋の収縮,緊張や腺の分泌をつかさどる神経系は自律神経系autonomic nervous systemである。この系では中枢神経系からの出力ニューロン(自律神経起始ニューロン)の軸索が,平滑筋,心筋,腺などの効果器に直接に連絡することはない。…
…脳はその前端部で目(複眼),触角などの感覚装置から光(紫外線,偏光も含む),機械的刺激による興奮を受け,中心部で情報の相関を行い,運動を制御し,また口や消化管の前部に向かう神経を経て,摂食行動を支配する。このような神経系は外観上脊椎動物の自律神経系に似る。エビでは巨大神経は尾の運動をつかさどり,迅速な逃避運動に関係する。…
※「自律神経系」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponsored by ![]()
夏の暑さに体が慣れること。数日から数十日間で起こる短期暑熱順化と、数年または数世代にかけて起こる長期暑熱順化とがある。→寒冷順化[補説]近年では、冷房設備の普及にともない短期暑熱順化が起こりにくくなっ...