デジタル大辞泉 「神経系」の意味・読み・例文・類語
しんけい‐けい【神経系】
Sponsored by ![]()
Sponsored by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponsored by ![]()
神経組織によって構成される器官系をいう。生体の特徴の一つは,刺激に対して反応することである。これは1個の細胞についてもいえることで,例えばアメーバのような単細胞生物を針で突いて刺激すると,刺激された部分とは反対の方向へ偽足を伸ばして刺激を避けようとする。一方,多細胞生物になるとしだいに細胞の役割分担がはっきりしてきて,幾種類かの細胞群の分業的かつ協調的な働きによって生体の機能がまっとうされるようになる。例えば,もっぱら情報の受容にかかわる細胞(感覚細胞)や,その情報に応じての生体の反応の発現に直接かかわる細胞(筋細胞や腺細胞)のほか,この両者の間にあって情報の伝達,処理,そしておそらくは貯蔵(記憶)にもかかわる細胞,すなわちニューロンneuron(神経細胞ともいう)が分化してくる。このようなニューロンの集合が神経系である。一般によく使われる神経nerveという日本語は,神気の〈神〉と中国医学でいう経脈の〈経〉とを組み合わせたものであり,《解体新書》(1774)ではじめて用いられた。
一般に生体の器官には実質と支質が区別される。それぞれの器官の特徴的な機能に直接かかわる細胞が実質(実質細胞)であり,実質細胞を一定の形にまとめあげ,これを維持する細胞,およびこのような細胞がつくり出す物質が支質である。例えば,炭水化物の貯蔵とその代謝の調節や,胆汁の生成などは肝臓の主要な機能であるが,これらの機能は肝臓の実質細胞である肝細胞の働きの表れである。一方,これらの肝細胞を肝臓という一つの形にまとめあげ,その栄養,維持にかかわっている結合組織も存在する。これらが肝臓の支質である。神経系ではニューロンがその実質細胞であるが,神経系にはこのほか支質にあたる細胞として神経膠(しんけいこう)(神経膠細胞,グリアgliaともいう)がある。
ニューロンは一般に神経膠よりも大きく,リボソームRNAを多量に含んでおり,したがってタンパク質合成能が大きい。これに対して神経膠は嫌気的解糖能が大きい。このほか,ニューロンと神経膠は脂質組成,細胞膜のガングリオシド組成などの点でも差がある。一方,ニューロンと神経膠は一定の条件下では代謝面で相関的変化を示すことも知られており,ニューロンは神経膠と結びついてはじめてその正常な代謝機能が発揮できる。
神経系の実質細胞,すなわちニューロンの機能は興奮を伝導することである。ニューロンの興奮伝導の能力はその細胞膜の性質(半透膜)と結びついている。
一般に細胞においては,それぞれの細胞の細胞膜の性質によって,その内部環境と外液との間にそれぞれの細胞に特有な大きさの電位差(静止電位resting potential)がある。興奮していないニューロンでは,細胞内液のカリウムイオンK⁺濃度は細胞外液のそれよりも高く,ナトリウムイオンNa⁺濃度は細胞外液のほうが高い。このようなイオン濃度のこう配を保つためには,ニューロンは代謝エネルギーを消費して多くのイオンを能動的に運搬しなければならない。このような静止電位は,いろいろなイオンに対する細胞膜の透過性を変化させると変化する。静止電位の変化を起こさせるような要因は,その細胞に対する刺激として働く。ニューロンに刺激が働いて,ニューロンのある小部分の静止電位(局所電位local potential)が変化し,その大きさが一定値よりも小さくなると,活動電位action potentialないしスパイクspikeが発生する。このスパイク電位は全か無の法則(悉無律(しつむりつ)all or none law)にしたがう。すなわち,刺激の強さがある一定の強さ(閾値(いきち))以上であれば,スパイクは刺激の強度の大小とは無関係に一定の大きさと時間経過をもって発生する。ニューロンはその細胞膜の性質によってこのようなスパイク電位を伝導し,さらに別のニューロンに伝達することができる。伝導されていくスパイクのことを神経インパルスという。このような興奮の伝導と伝達の能力がニューロンの情報伝達能の基本となっており,神経系はこのような実質細胞,すなわちニューロンの集合であるから,ニューロンをもって神経系の機能単位とみることができる。
ニューロンと呼ばれる細胞には三つの部分が区別される。すなわち,(1)核を取り囲む細胞体soma,(2)興奮を細胞体のほうへ伝える樹状突起dendrites,(3)興奮を細胞体から送り出す1本の軸索axon(ただし,分枝は多い)である。生体内でニューロンがスパイクを発生するときには,まず軸索の起始部(軸索小丘axon-hillock)のところからインパルスが形成される。また,軸索の末端(軸索終末axon terminal)では,次のニューロンに興奮の伝達が行われる。軸索は一般に長い突起であるから(脊髄から四肢の末端部にまで達するもの,大脳皮質から脊髄の末端部にまで達するものなど),神経繊維nerve fiberと呼ばれることが多い。また,軸索はしばしばリン脂質を主成分とする鞘(しよう)(髄鞘またはミエリン鞘 myelin sheath)をかぶっている。髄鞘をかぶっている軸索を有髄繊維(有髄神経繊維),髄鞘をもたない軸索を無髄繊維(無髄神経繊維)と呼ぶ。髄鞘の実体は神経膠の細胞膜であって,軸索のまわりをなん層にもなって取り巻いていることが多い。この髄鞘を形成する神経膠は,中枢神経系ではオリゴデンドログリアoligodendrogliaと呼ばれる神経膠の一種であり,末梢神経系ではシュワン細胞Schwann's cell(末梢神経系の神経膠)である。
ニューロンの樹状突起や軸索を細胞体から切り離すと,切り離された部分は必ず変性して消失してしまう。つまり,樹状突起や軸索はその栄養を細胞体に依存している。この意味でニューロンは神経系の機能単位であるとともに,栄養単位でもあるわけである。
軸索は長い突起であるにもかかわらず,その内部にタンパク質合成が行われていることを示す構造(粗面小胞体やリボソーム)をもっていない。したがって,軸索の栄養や機能に必要な物質の大部分は細胞体で合成され,はるばる軸索の末端まで輸送されなければならない。このような軸索内の物質の輸送(軸索輸送axoplasmic transport)は,軸索の内部に証明される〈流れ〉,すなわち軸索流axoplasmic flowによって行われている。軸索流による物質の輸送はエネルギーを必要とする過程であって,酸素とATPを必要とし,温度に敏感である。また,シアン化合物やジニトロフェノールなどの代謝阻害剤によっても停止する。軸索流にはいくつかのチャンネルがあるらしく,輸送される物質によってその速度が違う。おそいものでは1日1~2mm,速いものでは1日1000mm以上の速度で移動する。さらに,軸索流には細胞体から軸索の末端のほうへ向かうもの(順行性)ばかりでなく,軸索の末端から細胞体のほうへ向かうもの(逆行性)もあることが知られている。破傷風毒素やある種の神経親和性ウイルス(狂犬病ウイルス,ポリオウイルス,単純ヘルペスウイルスなど)などは軸索の末端からニューロン内に取り込まれた後,逆行性軸索流に乗ってニューロンの細胞体に達する。軸索流を担う構造として,とくに速い軸索流にとっては神経微小管が重要であることが知られているが,軸索流のメカニズムの詳細はまだよくわかっていない。
神経系は複数のニューロンから成る。したがって,ニューロンとニューロンの間で興奮の伝達が行われなければならない。ニューロン間の興奮の伝達は,通常,あるニューロンの軸索終末から次のニューロンの樹状突起ないし細胞体へと行われる。ニューロン間の興奮伝達が行われる部位をシナプスsynapseという。シナプスには興奮伝達の様式の違いによって電気シナプスと化学シナプスが区別されているが,電気シナプスの例は少ない。哺乳類のシナプスは通常,化学シナプスであり,興奮伝達は化学物質(化学伝達物質chemical transmitter)を介して行われる。化学シナプスのシナプス前要素,すなわち軸索終末には化学伝達物質を含むシナプス小胞synaptic vesicle(直径30~60nm)が集合しており,軸索終末の細胞膜は次のニューロンの樹状突起ないし細胞体の細胞膜と幅20~30nmの間隙(かんげき)(シナプス間隙synaptic cleft/ synaptic gap)をもって相対している。ニューロンの興奮にともなって軸索終末にインパルスが到達すると,シナプス小胞に含まれているノルアドレナリンやアセチルコリン,γ-アミノ酪酸gamma aminobutyric acid(略してGABA)などの伝達物質が分泌される。伝達物質はシナプス間隙を拡散して,次のニューロンの細胞膜の特殊な部位(伝達物質を認識する特殊な膜構造,すなわち受容体のある部位)に働いて,細胞膜のイオン透過性を変化させる。これにより,次のニューロンのシナプス部位の膜,すなわちシナプス後膜の脱分極(細胞膜の静止電位が小さくなる)が起こると次のニューロンの興奮性が高められ,過分極(細胞膜の静止電位がさらに高くなる)が起こると興奮性が抑制される。シナプスにおけるこのような興奮伝達の際にはカルシウムイオンCa2⁺が重要な役割を果たしていることが知られている。また,シナプス後膜には化学伝達物質に対する分解酵素が存在する。なお,感覚細胞とニューロンとのあいだの機能的連絡や,ニューロンと筋肉細胞ないし腺細胞とのあいだの機能的連絡も,一応,特別な形のシナプス連絡と考えてよかろう。
一般に個々のニューロンが合成する伝達物質は,1種類だけであるとされている。これをデールの原理Dale's principleというが,近年,多数の伝達物質ないしその候補が報告されており,この法則の修正を求める説も現れはじめている。
執筆者:水野 昇
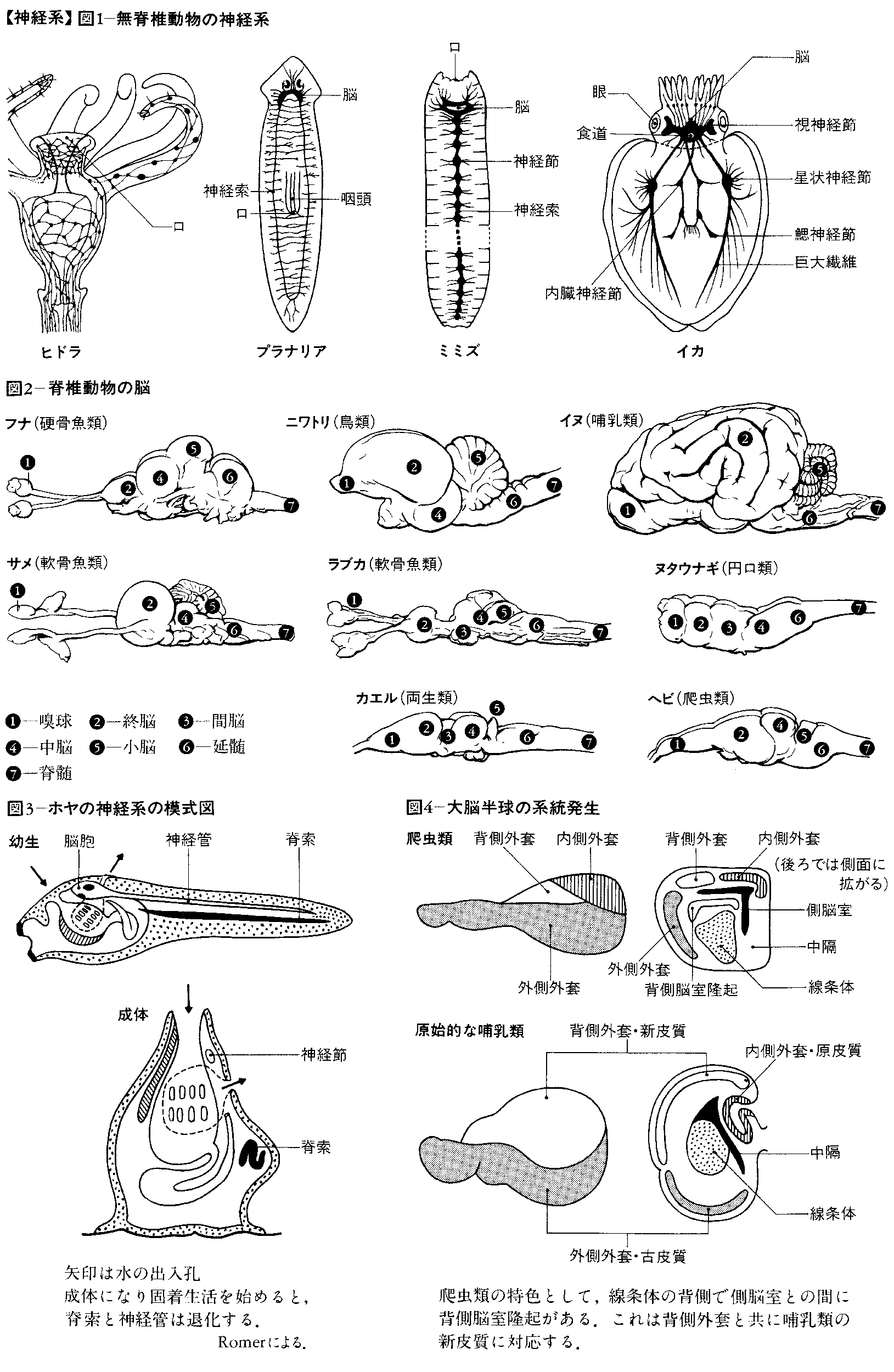
進化の初期の段階で出そろった無脊椎動物の神経細胞には,突起の少ない細胞と分泌性細胞が多い。無脊椎動物の神経系はいずれも外胚葉に由来するが,散在神経系と集中神経系の二つの様式がある。ヒドラのような腔腸動物では散在神経系で,神経細胞がその突起で互いに連結し,口を中心に体の全域に網状に分布する。興奮は一定の方向に進むのではなく,すべての経路を通って伝わる。クラゲになると神経細胞が所々に小集団をつくり,神経は放射状に走るようになる。腔腸動物より進化した無脊椎動物では,原則として左右相称の集中神経系が出現し,神経細胞体が比較的大きな集団を形成し,突起は索状に束になって体の腹側を縦に走る。また左右の神経系が融合する場合がある。頭端にある大きな細胞集団は脳,突起の束は神経と名付けられる。しかし棘皮(きよくひ)動物のヒトデやウニでは,体は幼生のとき左右相称で,成体になると放射相称に変わり,神経系は腔腸動物のように放射状,輪状になる。
プラナリアのような扁形動物では,はしご型で左右の索は互いに節状に横に走る繊維で連絡され,また索から末梢へ細い繊維が出て体の表面下に,あるいは消化管の壁に神経叢をつくる。環形動物たとえばミミズでは,腹側の神経索に沿って,各節ごとに細胞の集団つまり神経節が形成される。体の前方で咽頭の上にある神経節すなわち脳はよく発達し,神経系の集中化が進む。脳から出る神経は口の周辺,また体の後方に向かって走る。種によっては特定の部位の神経節が相対的に大きくなる。巨大繊維は伝導速度が速く,速やかに体を収縮させ,敵から身を守るのに役だつ。また各節にある輪状に走る筋と縦走する筋が,交互に収縮する一連の反射で,緩慢なはう運動が起こる。ゴカイでは光,重力,物質の性質に対する受容器が比較的複雑になり,積極的に自由な索餌行動ができる。カタツムリ,タコやイカのような軟体動物では,さらに体表面に触角,足などが分化し,神経系はいっそう集中的になるとともに,種によっては体とともにねじれる。
軟体動物さらにクモ,昆虫のような節足動物では,無脊椎動物の中でもっとも複雑な神経系になり,学習行動ができるようになる。基本設計はミミズと同じであるが,神経機能が頭部にいっそう集中する傾向がある。節状の神経節は融合し,消化管壁,外套膜,足の基部などに集中する。また脳(神経節)がよく発達し,食道を取り巻くようになる。したがって位置移動,摂食行動のような特殊化した運動は,神経系の特定部分からの制御を受ける。これらの動物は脊椎動物に似た目をもち,その神経系は対象物の識別,学習および記憶のような連合機能を果たす。軟体動物の外套に分布する巨大繊維の興奮はきわめて速い運動を起こす。節足動物では移動に役だつ脚と羽が発達し,脳がさらに分化する。脳はその前端部で目(複眼),触角などの感覚装置から光(紫外線,偏光も含む),機械的刺激による興奮を受け,中心部で情報の相関を行い,運動を制御し,また口や消化管の前部に向かう神経を経て,摂食行動を支配する。このような神経系は外観上脊椎動物の自律神経系に似る。エビでは巨大神経は尾の運動をつかさどり,迅速な逃避運動に関係する。
セミ,コオロギのような鳴く昆虫では,脚の関節に音の受容器があり,知覚神経が分布する。節足動物では脳の神経分泌物は側心体に蓄えられ,血液中に出て,他の内分泌細胞を刺激して,そのホルモンで変態を起こさせる。社会性昆虫では,その分業に応じて神経系の様相が異なる。たとえばアリでは,働きアリは他のアリより運動をつかさどると考えられる有柄体(キノコ体)が大きいが,目と光の興奮を受ける視葉は退化する。女王アリと雄アリは視葉や目が相対的に大きい。これは性行動に関係すると思われる。
全般に無脊椎動物の神経系は,神経細胞の数が少なく,しかも細胞体が大きく,巨大繊維が含まれているので,行動の有効な研究モデルになる。
終生脊索をもち,脊椎動物の祖先と考えられるホヤ,ナメクジウオの類でも,体の背側正中線上に,ヒトと同様に内腔をもった管状の神経系が発生する。脊椎動物の神経系も,無脊椎動物と同様に,神経細胞の集積によって形成される。神経管の前端部は脳,他の部分は脊髄となる。脳と脊髄つまり中枢神経系と末梢の受容器および効果器とを連絡するのが末梢神経系である。
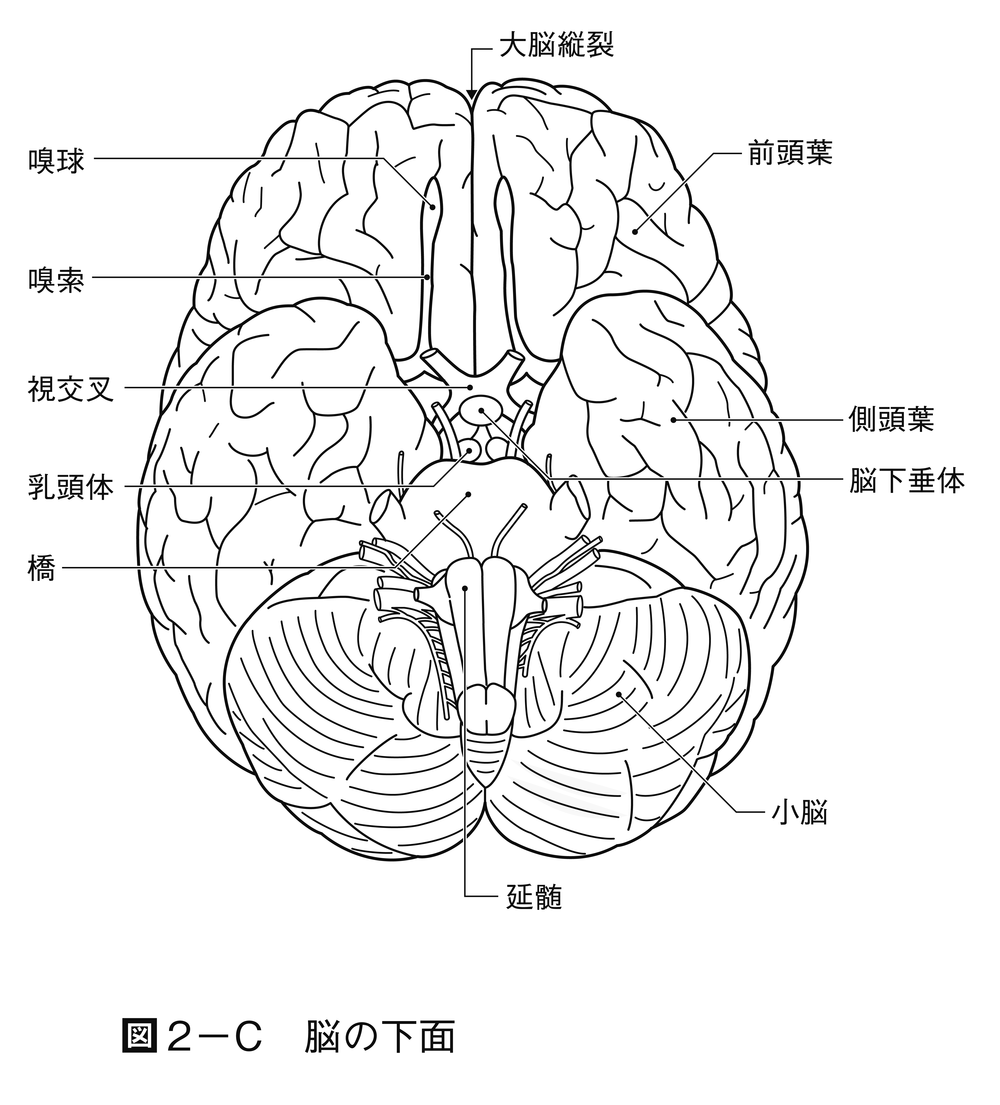
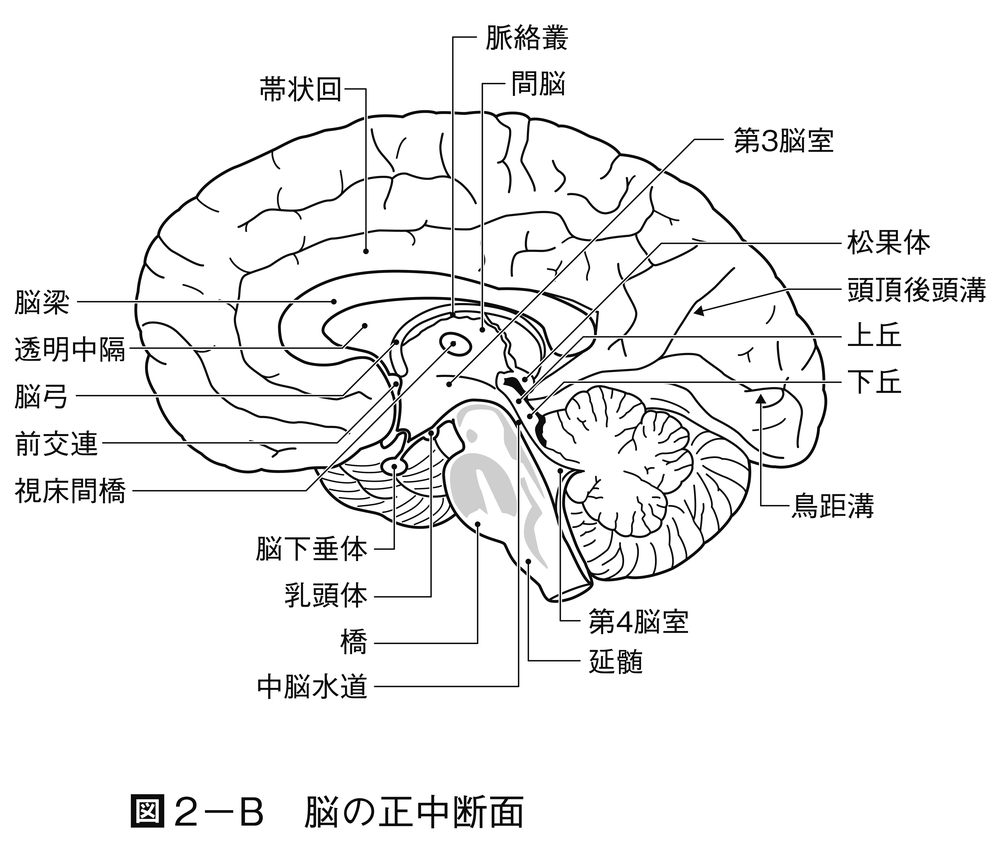
脊椎動物の進化の初期において,特殊化した感覚系が脳の各主要部分と連絡する。すなわち嗅覚系は前脳(前脳胞)と,視覚系は中脳(中脳胞)と,波動のような機械的感覚系は後脳(後脳胞)と関連する。さらにこれらの3部分つまり脳胞brain vesicleでは,細胞集団が背外側に発達し,大脳,視蓋および小脳が形成される。この発達の結果,3部分は5部分に変形されて脳が完成する。すなわち終脳,間脳,中脳,後脳および髄脳が区分される。小脳を除いた後脳の腹側部分と髄脳とを合わせた部分が菱脳(りようのう)と名付けられる。哺乳類以外ではこれを延髄というが,哺乳類では髄脳の部分だけが延髄と名づけられる。髄脳の背側部は外側に展開したため,天井は膜状になる。このような形成過程はヒトにおいても同じである。これらの神経中枢は進化に並行して,さらに神経要素が増加し,知覚神経と運動神経との間にいっそう広範囲の協調coordinationや相関correlationが可能になる。脊椎動物の各綱の脳における,以上述べた主要部分の形成過程は図2に示してある。脳の主要部分の大きさと生活上でのその部分の働きの優位性との間には関係がある。
中脳の天井つまり視蓋は,元来第一次の視覚中枢で,左右交叉した視神経が入る。視蓋は硬骨魚類,両生類では神経系の中で優位の相関中枢で,生体の活動を支配し,大きな影響を与える。爬虫類と鳥類になると,視蓋の支配権の上に,大脳半球の影響が加わりはじめる。哺乳類では,視蓋の統合integrationの能力は相対的に減退して,代りに大脳皮質が発達し,間脳は各種の感覚伝達の中継地となる。一般に前進的な進化に並行して,神経機能の中心は下位脳幹部から高位の中枢へとしだいに脳の前端部に移動する(中枢の頭方移動)。また哺乳類では機能が優位になった大脳が小脳,延髄,脊髄などの下位の部分に高度の指示を与えるために神経路を出す。その神経繊維が通る部分つまり橋,錐体などが後脳の腹側部に付加される。
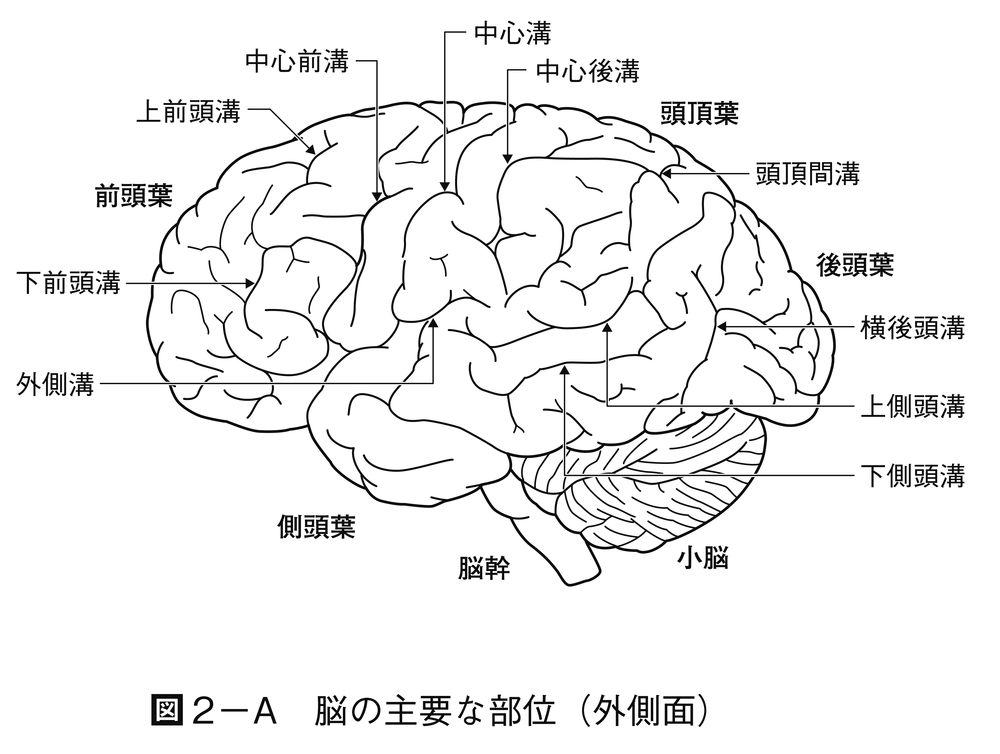
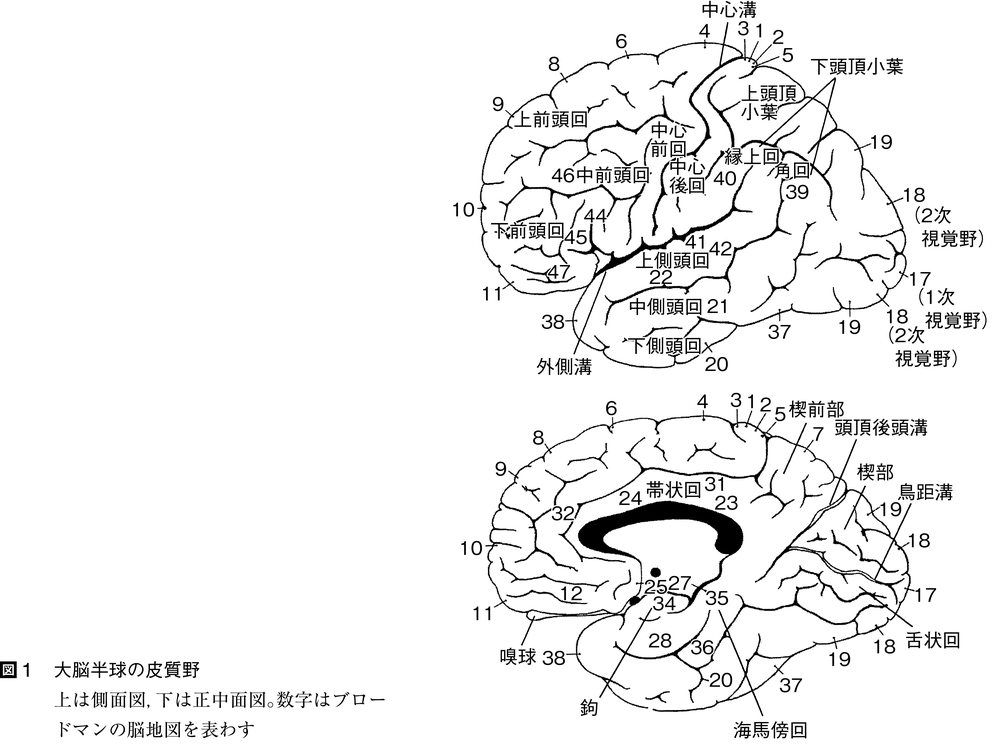
脊椎動物の前進的進化の特色は,大脳半球の表面つまり大脳皮質が連合associationの領域として重要性を増すことである。大脳半球は知覚域に属し,はじめは嗅覚入力を受けて,興奮を脳幹部に出す働きをする部分であった。半球の壁は背側の外套(がいとう)部palliumと腹側の外套下部subpalliumに分かれ,さらに外套部は内側外套,背側外套および外側外套に分かれる。外套下部は内側の中隔と外側の線条体から構成される。外套下部は相対的に脊椎動物の各綱を通して変わらないが,外套部は哺乳類になると表面の細胞シートつまり皮質がひじょうに発達し,内側の原皮質,背側の新皮質および外側の古皮質が明りょうに区別される。種によっては新皮質はしわを生じて表面積を広げ,脳の他の部分を覆う。新皮質は連合,学習のような高度の神経活動を受けもつことになる。ヒトのような高次の神経活動が要求される種では大脳はひじょうに大きくなる。
執筆者:正井 秀夫
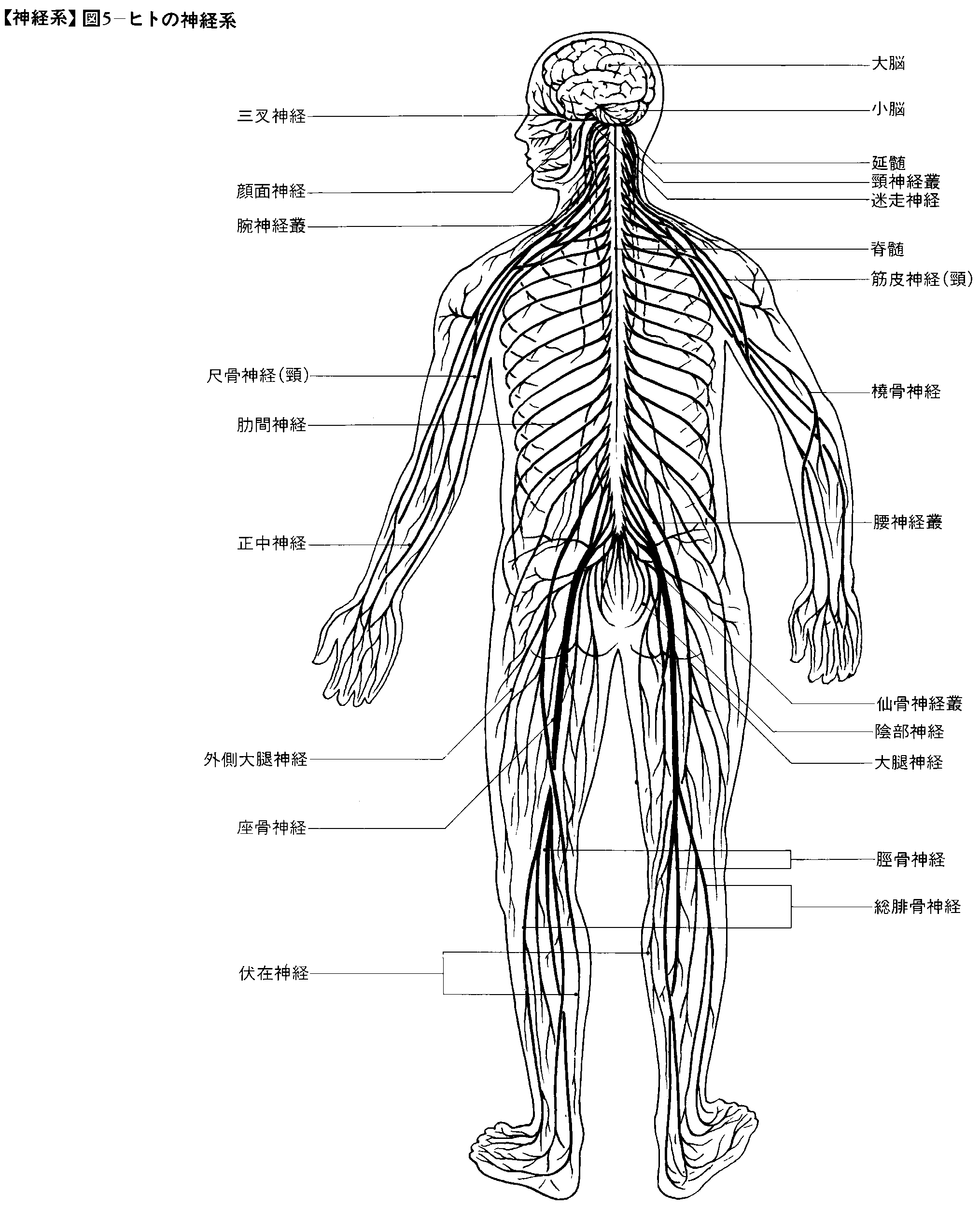
脊椎動物の神経系には中枢神経系central nervous systemと末梢神経系peripheral nervous systemが区別される。中枢神経系とは脳brainと脊髄spinal cordのことである。末梢神経系は生体の各部と中枢神経系との間を連絡する神経系であって,脳に出入する脳神経cranial nervesと,脊髄に出入する脊髄神経spinal nervesに分類される。末梢神経系のことを脳脊髄神経系craniospinal nervous systemと呼ぶこともある。
脳に出入する末梢神経系,すなわち脳神経は爬虫類以上の脊椎動物では一般に12対ある。嗅神経,視神経,内耳神経(平衡聴覚神経または前庭蝸牛(かぎゆう)神経)のほか,眼球を動かす動眼神経,滑車神経,外転神経,表情筋を動かす顔面神経,頭顔部の感覚を伝達する神経繊維のほかに,咀嚼(そしやく)筋を動かす神経繊維を含む三叉(さんさ)神経,そのほか舌咽神経,迷走神経,副神経,舌下神経である。以上12対の脳神経のうち,副神経は本来は迷走神経の副枝である。また,舌下神経は舌を欠く動物には存在しない。副神経や舌下神経が,はっきり独立した脳神経としてみとめられるのは,爬虫類以上の動物だけである。両生類以下の脊椎動物の脳神経は一般に10対である。一方,両生類以下の水生動物には側線神経があり,三叉神経,顔面神経,迷走神経と連なっているが,この側線神経は陸生の動物では退化している。
脳に出入する脳神経が,頭顔部の機能にかかわる末梢神経であるのに対し,脊髄に出入する脊髄神経は,主として頸部以下の身体部位の機能にかかわる神経である。脊髄はその対応する身体部位にしたがって,頸髄,胸髄,腰髄,仙髄,尾髄に区分され,一方,それらの部位に出入する脊髄神経は頸神経cervical nerves,胸神経thoracic nerves,腰神経lumbar nerves,仙骨神経sacral nerves,尾骨神経coccygeal nervesと呼ばれる。ヒトでは,頸神経は8対,胸神経は12対,腰神経は5対,仙骨神経は5対,尾骨神経は1対で,全部で31対の脊髄神経がある。頸神経の数は,ナマケモノなどのごく一部の例外を除いて,哺乳類では一般に8対である。しかし,脊髄神経の数は椎骨の数の変化にしたがって変わるから,脊椎動物全体としては一般に変化に富む。例えばネコの胸神経は13対,腰神経は7対,仙骨神経は3対であり,カエルでは脊髄神経は全部で10対ほどである。
末梢神経を構成する神経繊維のなかには興奮を身体の各部から中枢神経系のほうへ伝導する求心性繊維(求心性神経繊維)centripetal fiberと,興奮を中枢神経系から身体の各部へと伝導する遠心性繊維(遠心性神経繊維)centrifugal fiberとがある。脊髄神経の領域では,求心性繊維は脊髄の背側(後方)から脊髄に入るが,この部分を後根という。一方,遠心性繊維は脊髄の腹側(前方)から脊髄を出ていくが,この部分を前根という。後根と前根は脊髄のすぐ近くで合一しており,この合一によって31対の脊髄神経が形成されるのである。したがって,それぞれの脊髄神経には求心性繊維と遠心性繊維が含まれており,前者は後根を通って脊髄に入り,後者は前根を通って脊髄から出てくる。
要するに求心性神経繊維は,皮膚,筋,筋膜,関節,骨膜,内臓,血管などからの情報を,中枢神経系に伝達する感覚神経繊維である。これらの情報には,意識にのぼるものもあればのぼらないものもある。一方,遠心性神経繊維には骨格筋の運動をつかさどる運動神経繊維と,平滑筋・心筋・腺を支配する自律神経繊維がある。身体の各部にみられるいわゆる神経(繊維束)には,ほとんどの場合,以上の感覚神経繊維,運動神経繊維,自律神経繊維がいろいろの割合で混在しているのである。
身体の各部からの情報を中枢神経系に伝達するニューロン,すなわち求心性神経繊維を出すニューロンの細胞体は中枢神経系の外に存在し,集合して感覚神経節sensory ganglionをつくっている。感覚神経節のニューロンは,1本の突起を身体の末梢のほうに伸ばしており,他の1本を中枢神経系に向かって伸ばしている。感覚神経節は,脊髄神経の領域では,前根と後根が合一して脊髄神経が形成されるところよりも,少々中枢側で後根の途中に存在する。したがって,それぞれの脊髄神経に対応して1個の感覚神経節があり,これらを脊髄神経節spinal ganglionまたは後根神経節dorsal root ganglionという。感覚神経節は求心性神経繊維を中枢神経系に送るニューロンの細胞体の集合であるから,脳神経の領域でも求心性神経繊維を含む脳神経にも感覚神経節がある。すなわち,三叉神経の三叉神経節(半月神経節,ガッセルの神経節),顔面神経(厳密には中間神経)の膝(しつ)神経節,内耳神経のらせん神経節と前庭神経節,舌咽神経の上神経節(内神経節)と下神経節(外神経節,岩様部神経節),迷走神経の上神経節(頸静脈神経節)と下神経節(節状神経節)である。
感覚神経節のニューロンは,皮膚,筋肉,筋膜,関節,靱帯,骨膜などからの情報のほか,内臓や血管などからの情報をも中枢神経系へ伝達する。これらのほか,脳神経の領域(頭顔部)には特殊に発達した感覚器からの情報(嗅覚,視覚,平衡覚,聴覚,味覚)を伝達する求心性繊維がある。これらの特殊感覚神経繊維は,嗅神経,視神経,内耳神経(平衡聴覚神経)のほか,味覚繊維は顔面神経(厳密には中間神経)と舌咽神経(一部はさらに迷走神経)のなかを走って脳に入る。
身体の各部からの情報を中枢神経系に伝達するニューロンの細胞体が中枢神経系の外部,すなわち感覚神経節に集合しているのに対し,中枢神経系からの信号を身体各部の効果器(筋肉と腺)に伝達するニューロンの細胞体は中枢神経系内にある。したがって,このような中枢神経系からの出力ニューロンでは,その樹状突起,細胞体と軸索のはじめの部分は中枢神経系内にあり,その軸索の大部分が末梢神経系内を走ることになる。ところで,生体の効果器には,骨格筋のように動物性の機能(運動)にかかわるものと,平滑筋,心筋,腺のように植物性の機能(消化,吸収,呼吸,循環,分泌,栄養,生殖など)にかかわるものとがある。これらの効果器のうち,骨格筋には中枢神経系の出力ニューロンの一つである運動ニューロンmotoneuronの軸索が直接連絡している。ポリオのウイルスは,この運動ニューロンを侵す神経親和性のウイルスである。脊髄神経の領域では,運動ニューロンは脊髄の前角に集合しており,脊髄前角細胞anterior horn cell,ventral horn cellと呼ばれる。脳神経の領域においても,動眼神経,滑車神経,三叉神経,顔面神経,舌咽神経,迷走神経,副神経,舌下神経には,運動ニューロンの軸索が含まれており,これらのニューロンの細胞体は動眼神経核(中脳),滑車神経核(中脳と橋の境界レベル),三叉神経運動核(橋),顔面神経核(延髄),疑核(延髄にあり,舌咽神経と迷走神経に遠心性繊維を送る),副神経核(頸髄),舌下神経核(延髄)に連合している。一般にこれら運動ニューロンの化学伝達物質はアセチルコリンである。
一方,平滑筋の収縮,緊張や腺の分泌をつかさどる神経系は自律神経系autonomic nervous systemである。この系では中枢神経系からの出力ニューロン(自律神経起始ニューロン)の軸索が,平滑筋,心筋,腺などの効果器に直接に連絡することはない。すなわち,中枢神経系と効果器の間には少なくとももう一つ別の自律神経ニューロンが介在し,このような自律神経ニューロンの集合を自律神経節autonomic ganglionという。中枢神経系内に細胞体をもつ自律神経起始ニューロンのことを節前ニューロンpreganglionic neuron,その軸索を節前繊維preganglionic fiberと呼び,自律神経節に細胞体をもつニューロンのことを節後ニューロンpostganglionic neuron,その軸索を節後繊維postganglionic fiberと呼ぶ。自律神経節で節後ニューロンとシナプス結合する節前ニューロンが,アセチルコリンを化学伝達物質とするアセチルコリン作動性ニューロンであるのに対して,効果器に直接に連絡する節後ニューロンには,アセチルコリン作動性のものと,ノルアドレナリン作動性のものとがある。ノルアドレナリン作動性の節後ニューロンに連絡する系を交感神経系sympathetic nervous systemと呼び,アセチルコリン作動性の節後ニューロンに連絡する系を副交感神経系parasympathetic nervous systemという。ヒトの交感神経系の出力ニューロン(節前ニューロン)の細胞体は,胸髄と腰髄上部の主として側角(中間外側核)に集合している。これらのニューロンの軸索は前根を通って脊髄から出ており,末梢神経系内に存在する種々の自律神経節において,節後ニューロンとシナプス結合する。次いで節後ニューロンの軸索は動脈壁に沿って走ったり,脊髄神経といっしょに走ったりして全身に分布する。一方,ヒトの副交感神経系の出力ニューロン(節前ニューロン)の細胞体は仙髄と脳にある。このうち仙髄から起こる副交感神経系は骨盤内臓を支配するから,この系が損傷されると膀胱直腸障害(排尿障害や排便障害)が起こったり,インポテンツが起こる。脳にある副交感神経系の節前ニューロンの軸索は,動眼神経,顔面神経(厳密には中間神経),舌咽神経,迷走神経のなかを走る。これらは瞳孔収縮筋,毛様筋を支配するほか,涙腺,鼻腺,口腔腺(唾液腺など)などの分泌をつかさどるが,とくに迷走神経のなかを走る副交感神経繊維は胸腔や腹腔にまで達し(〈迷走〉),胸部内臓(心臓や肺など)や腹腔内臓(胃,小腸,肝臓など)の運動,分泌をつかさどる。
それぞれの脊髄神経は前根と後根が合一して形成されるが,これらの脊髄神経は脊椎管を出ると直ちに大きく前枝と後枝に分枝する。前根が遠心性繊維(運動ニューロンの軸索と自律神経節前ニューロンの軸索),後根が求心性繊維(脊髄神経節ニューロンの中枢神経系へ向かう突起)で形成されているのに対し,前枝と後枝には遠心性繊維と求心性繊維の両方が含まれる。すなわち,それぞれの脊髄神経の前枝と後枝には運動ニューロンの軸索,自律神経ニューロンの軸索,および感覚神経節細胞の突起すなわち求心性繊維がさまざまの割合で含まれているのである。前枝と後枝のうち,後枝が体幹の背側部に分布するのに対し,前枝は体幹の腹側部と四肢に分布する。この場合,それぞれの脊髄神経の後枝と胸神経の前枝とがそのまま走って体幹に分布するのに対して,頸神経,腰神経,仙骨神経,尾骨神経の前枝は頸部や四肢に向かって走るのに先立って,それぞれ隣り合う前枝の間で神経繊維の交換を行う。このため,脊髄神経前枝の間に神経繊維の交錯を生じ,一種のネットワークができる。これを神経叢(または神経集網)plexusという。このような神経叢として,ヒトでは頸神経叢(第1頸神経から第4頸神経までの前枝によって形成される),腕神経叢(主として第5頸神経から第1胸神経までの前枝によって形成),腰神経叢(主として第12胸神経から第4腰神経までの前枝によって形成),仙骨神経叢(主として第4腰神経から第3仙骨神経までの前枝によって形成),陰部神経叢(主として第2から第4までの仙骨神経の前枝によって形成),尾骨神経叢(主として第4および第5仙骨神経と尾骨神経の前枝によって形成)がある。
以上の神経叢のうち腕神経叢brachial plexusには上肢を支配する腋窩(えきか)神経,筋皮神経,正中神経,尺骨神経,橈骨(とうこつ)神経などがつづいている。腕神経叢は頸の付け根の付近にあるから,交通事故などの際に頸が一方に強い力で引っ張られると,腕神経叢を形成している神経繊維がその根のところで損傷されやすい。腋窩神経が損傷されると,上腕の外転(上腕を外側に向かって水平に上げること)ができなくなる。また,ひじを打ったとき小指のあたりに〈しびれ〉を覚えることがあるのは,ひじの後内側部を走る尺骨神経が打撃を受けるためである。橈骨神経が損傷されると,手が手首のところで垂れ下がったままの状態になり,指を伸ばすこともできなくなる。正中神経が損傷されると,親指に近い手のひらの領域の感覚麻痺が起こるとともに,親指の屈曲ができなくなる。
下肢を支配する末梢神経は腰神経叢lumbar plexusや仙骨神経叢sacral plexusにつづく。上腿の腹側(前面)の筋や皮膚を支配する大腿神経は腰神経叢につづく末梢神経である。上腿の背側(後面)および下腿,足の筋や皮膚を支配する座骨神経は仙骨神経叢につづく末梢神経である。以上のように脊髄神経の前枝は,支配部位に分布するに先立って神経叢を形成するのであるが,前枝のうちで胸神経の前枝だけは,一部が腕神経叢や腰神経叢に参加するほかは,神経叢を形成することなく,脊髄神経の後枝と同様,そのまま走って体幹の腹側部(前面)と側面に分布する。このような胸神経の前枝を肋間神経と呼ぶ。肋間神経はヒトでは12対ある。
→自律神経系 →ニューロン
執筆者:水野 昇
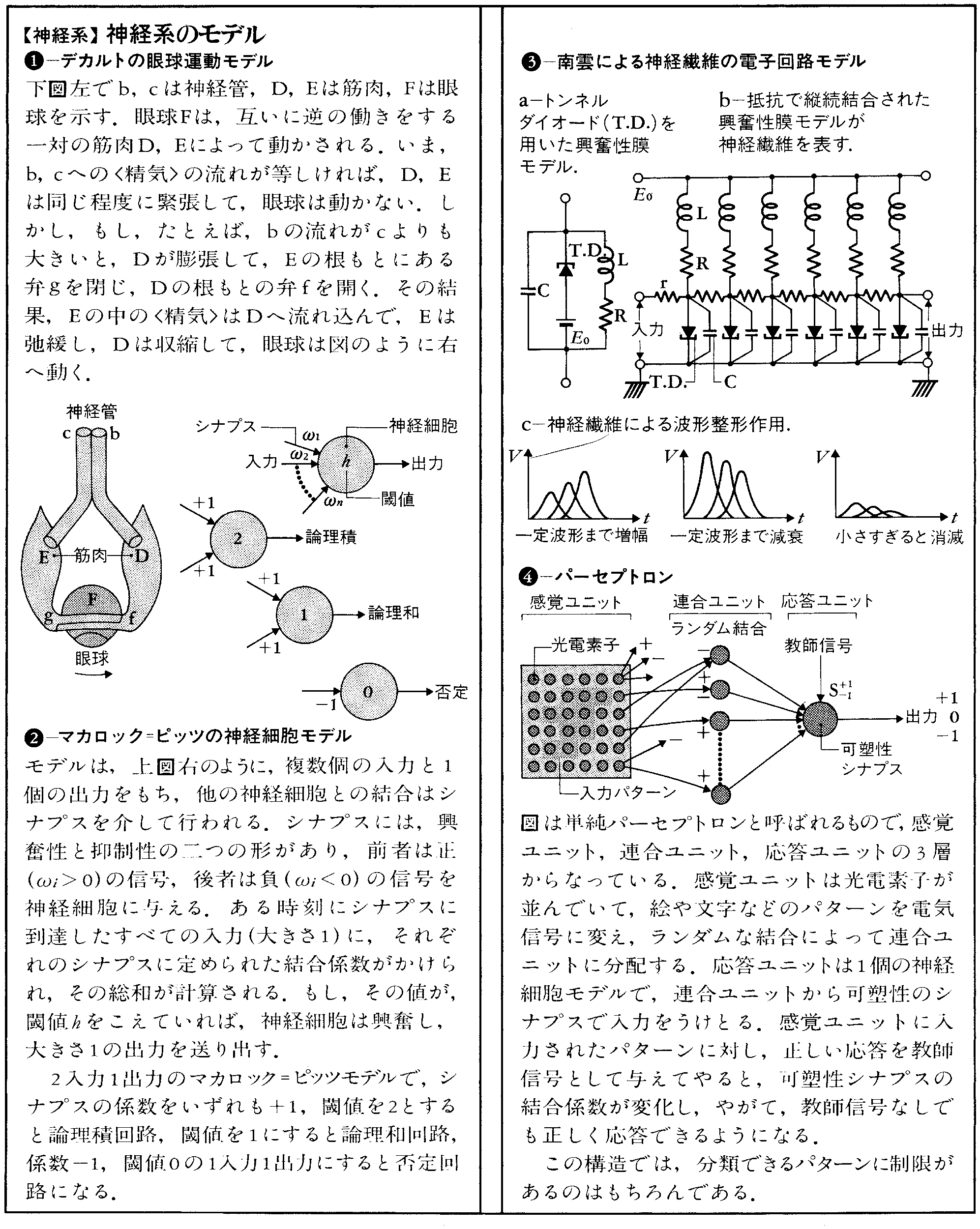
神経系のモデルとしては,古くはデカルトの眼球運動のモデル(1637)がある。デカルトは精神と肉体を分離し,肉体は機械的に理解できるという二元論の立場をとり,両者を結ぶものとして〈精気〉を考えた。デカルトによると,脳室に蓄えられている〈精気〉が神経管を通って筋肉に送り込まれ,眼球運動が生ずるという。これは当時流行していた水力オートマトン(からくり人形)のしくみとの類推で,眼球運動を理解しようとしたものと思われる(コラム(1)参照)。
このようにモデルとは,一般に,現象の背後にある未知のしくみを,既知の世界の言葉を使って表現したものということができる。さて,現在の神経系のモデルは,実体よりも神経系の機能を現すことに重点がおかれている。すなわち,神経生理学の進歩によって,脳の構成要素である神経細胞の性質と結合様式がかなり明らかにされてきた。そこで,神経細胞の基本的性質を,電子回路あるいは数式によって表現し,それを組み合わせてできる神経回路の動作を解析する,あるいは,ある機能をもたせるには,どのような構成にしたらよいかを検討する。これによって,神経系を理解しようというものである。
まず,神経細胞の閾作用,すなわち入力の和がある値をこえたときのみ,一定の大きさの出力がでるという性質を定式化したものとして,マカロックW.S.Mc-CullochとピッツW.H.Pittsの数学モデル(1943)が知られている。神経細胞の複数の入力をxi(i=1,2,……,n),それぞれの入力に対する結合係数をωi,閾値をhとしたとき,出力xは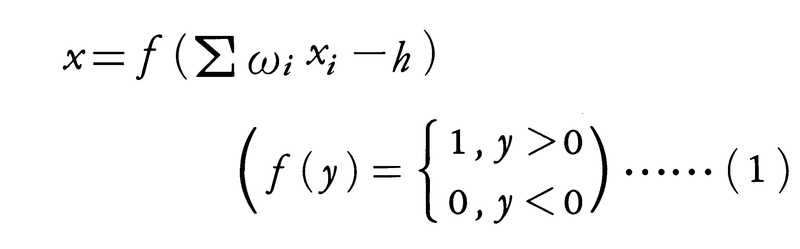
で与えられるとする。最も単純な形式をとり,神経回路網の解析の基本モデルとなっている(コラム(2)参照)。
神経細胞のもう一つの特性として,神経繊維では,信号として一定波形のインパルスが伝播(でんぱ)することが知られている。このインパルスの発生を説明するモデルとしては,ホジキンA.L.HodgkinとハクスリーA.F.Huxleyが,ヤリイカの巨大神経繊維の実験データに基づいてたてたH-H方程式(1952)がある。これは,神経繊維の膜を横切って流れるイオン電流についてのひと組みの微分方程式であるが,この性質が,フィツヒューR.Fitzhugh,南雲仁一らによって詳しく調べられ,より扱いやすい数学モデルおよび電子回路モデル(1962)が提案された。南雲らは,これを用い,神経インパルスが減衰したり,ひずんだりすることなしに繊維を伝播することを示した。これを波形整形作用,このような性質をもつ線路を能動線路という(コラム(3)参照)。
次に,神経細胞で構成される神経回路の機能の解析,あるいは,ある機能をもつ神経回路の合成が問題となる。この研究は,最初,二つの方向に分かれた。一つは,神経細胞のもつ不応期や順応などの性質をとり入れたより複雑なモデルを用い,小規模な神経回路を対象としたものである。これによって,感覚神経系における情報の符号化や運動系におけるリズム発生の機構などが解析されている。他の一つは,できるだけ単純な神経細胞モデルを用い,それらが多数ランダムに結合した神経回路を対象とするものである。そこでは,個々の細胞の動作は問題とせず,神経回路全体の動作を表す量(これを巨視的状態量と呼ぶ。たとえば平均活動度)をとりあげる。甘利俊一はこれを統計神経力学と呼び,結合係数ωiや閾値hなどのパラメーターが巨視的状態に対してもつ意味などを明らかにした。
ランダム神経回路は,理論的に取り扱いやすく,脳の機能を解析する出発点となった。次は,神経回路にパターン認識など高次の機能をもたせることである。それには,一様ランダムでなく,適当な構造をもたせないといけない。どのような機能にはどのような構造が必要か。それを知る方法として,脳の可塑性に基づく自己組織の原理がとられた。つまり,神経回路にある目標をあたえ,学習によってその目標を達成するように自分の構造をきめさせるというものである。
脳の可塑性が,シナプスの結合係数の変化によるという考えは,現在,最も一般的にとられている考えである。ヘッブD.O.Hebbは,シナプスによって結合した二つの神経細胞がともに興奮したとき,その結合係数が増すという考えを出していた(1949)。これを数式で表すと次のようになる。すなわち,細胞iから細胞jへのシナプスの結合係数をωij,各細胞の状態をxi,xj(興奮していれば1,していなければ0)とすると,変化量⊿ωijは,
⊿ωij=αxixj-βωij ……(2)
で表される。第1項は,二つの細胞が同時に興奮したときの結合の強化を,第2項は自然の減衰を示している。
このような可塑性のある神経細胞を組み合わせた自己組織回路のモデルとして,ローゼンブラットF.RosenblattのパーセプトロンPerceptron(1962)がある。分類すべきパターンを与えたとき,出力が正しい応答を示すように,シナプスの結合係数を変化させる。小脳は随意運動に関し重要な機能をもつといわれているが,マーD.Marrは,小脳の苔状繊維-顆粒(かりゆう)細胞-プルキニエ細胞の系にパーセプトロンの考えを適用した(コラム(4)参照)。その後,伊藤正男らによって,その生理学的根拠が与えられ始めており,モデルの有効性を示す例として注目される。
視覚神経系に関し,ヒューベルD.H.HubelとウィーゼルT.N.Wieselによる特徴抽出の階層性の神経生理学的研究がある。これによると,階層構造の初段の細胞は,それぞれ,特定の方向の線分の検出を分担することになる。この方向検出の機能が,生後の特定の時期の経験に強く依存することも知られた。このことから,この階層構造の自己組織の過程が問題となる。これについては,マースバーグC.von der Malsburgはじめ,いくつかのモデルが報告されているが,基本的な考えは,一つの細胞についているシナプスの結合係数の総和を一定に制限し,一つの方向の線分に敏感になった神経細胞は,他の方向の線分には反応しにくくしようというものである。また,福島邦彦は,多層の神経回路を用意し,ある領域ごとに最も強く活動している細胞への結合だけが強化されるという条件で,各層間の結合を自己組織化させた。その結果,このモデルに視覚神経系のパターン認識の機能をもたせることに成功している。彼はこれをコグニトロンCognitronと名づけた。
人間の記憶に特徴的な連想機能をもった神経回路も,自己組織によって構成することができる。中野馨は,神経回路に二つの入力パターンA,Bを同時に,繰り返し与え,そのつど,(2)に従ってシナプス結合を強化すると,神経回路の自己組織化が進み,Aなる入力からBなる出力の得られることを示した。この回路はアソシアトロンAssociatronと呼ばれている。
このように,神経系の高次の機能は,学習による自己組織によって発達してきたことが理解できる。
執筆者:鈴木 良次
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponsored by ![]()
出典 最新 心理学事典最新 心理学事典について 情報
Sponsored by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponsored by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponsored by ![]()
Sponsored by ![]()
夏の暑さに体が慣れること。数日から数十日間で起こる短期暑熱順化と、数年または数世代にかけて起こる長期暑熱順化とがある。→寒冷順化[補説]近年では、冷房設備の普及にともない短期暑熱順化が起こりにくくなっ...