デジタル大辞泉 「聴覚」の意味・読み・例文・類語
ちょう‐かく〔チヤウ‐〕【聴覚】
[類語]視覚・嗅覚・味覚・触覚
Sponsored by ![]()
Sponsored by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponsored by ![]()
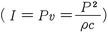 。密度と音速とをほぼ一定とみなせば,強度は音圧の2乗に比例する。聴覚刺激として適正な強度のダイナミック・レンジは非常に広く,エネルギー(単位W/m2:ワット毎平方メートル)で表わすとおおよそ10-12W/m2から103W/m2となる。そのため,聴覚刺激の呈示レベルを記述する際には強度レベルないし音圧レベルを使用する慣例がある。強度レベルは
。密度と音速とをほぼ一定とみなせば,強度は音圧の2乗に比例する。聴覚刺激として適正な強度のダイナミック・レンジは非常に広く,エネルギー(単位W/m2:ワット毎平方メートル)で表わすとおおよそ10-12W/m2から103W/m2となる。そのため,聴覚刺激の呈示レベルを記述する際には強度レベルないし音圧レベルを使用する慣例がある。強度レベルは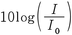 ,音圧レベルは
,音圧レベルは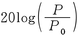 で算出し,単位は㏈を使用する。ただし,ここでI0,P0はそれぞれ基準強度,基準音圧であり,10-12W/m2,20μPaである(Pa:圧力の単位パスカル)。強度レベルと音圧レベルは厳密には異なるものであるが,実際上は両者の値に大きな差が生じることはなく,聴覚刺激の呈示レベルを記述するにあたって強度レベル,音圧レベルのいずれを用いてもかまわない。実際の物理的な測定では,大半のトランスデューサ(マイクロフォン)が音圧を測定するものであるため音圧レベルが使用される場合が圧倒的に多い。
で算出し,単位は㏈を使用する。ただし,ここでI0,P0はそれぞれ基準強度,基準音圧であり,10-12W/m2,20μPaである(Pa:圧力の単位パスカル)。強度レベルと音圧レベルは厳密には異なるものであるが,実際上は両者の値に大きな差が生じることはなく,聴覚刺激の呈示レベルを記述するにあたって強度レベル,音圧レベルのいずれを用いてもかまわない。実際の物理的な測定では,大半のトランスデューサ(マイクロフォン)が音圧を測定するものであるため音圧レベルが使用される場合が圧倒的に多い。
図6 ノッチ・ノイズ法によって推定され…

図5 ノッチ・ノイズ・マスカーと聴覚フ…
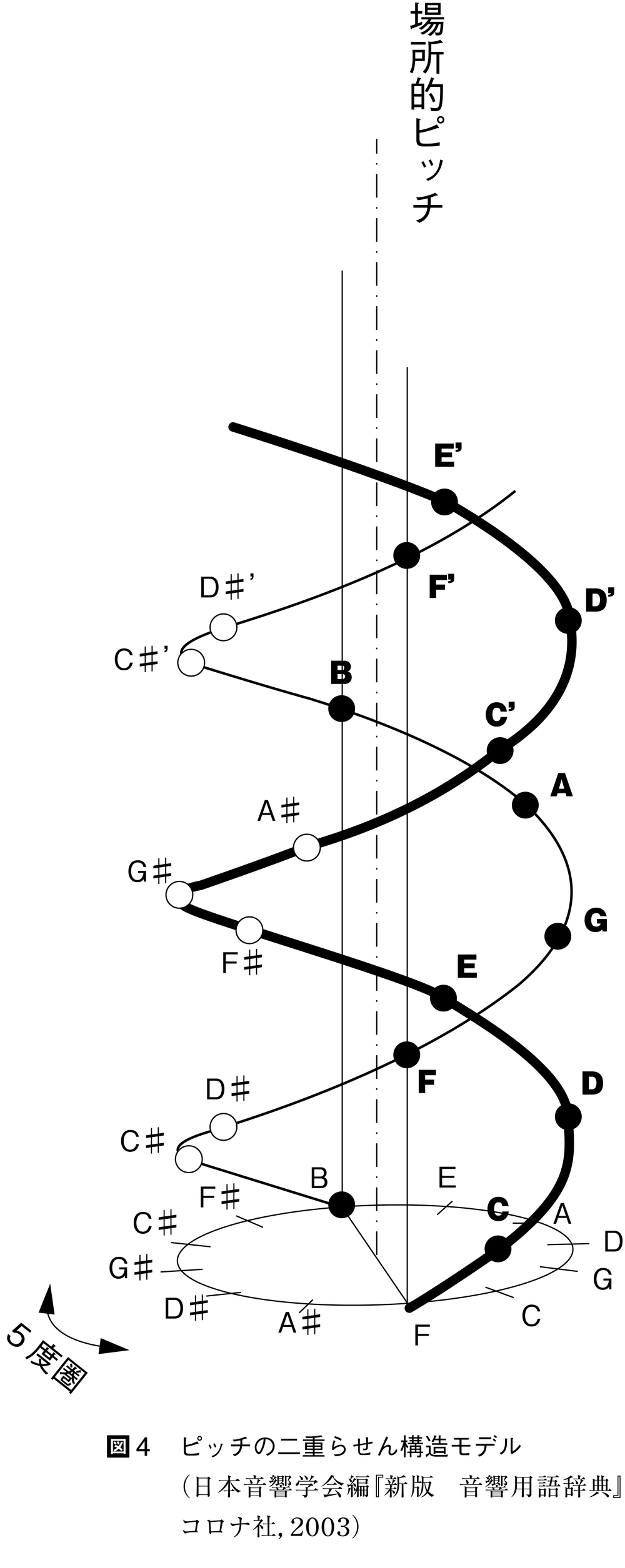
図4 ピッチの二重らせん構造モデル
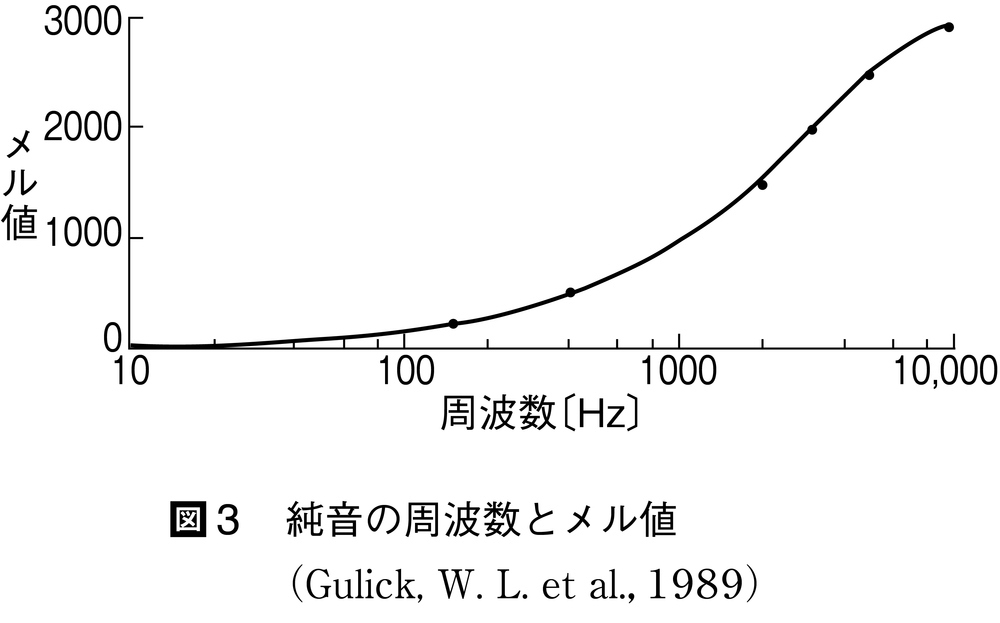
図3 純音の周波数とメル値
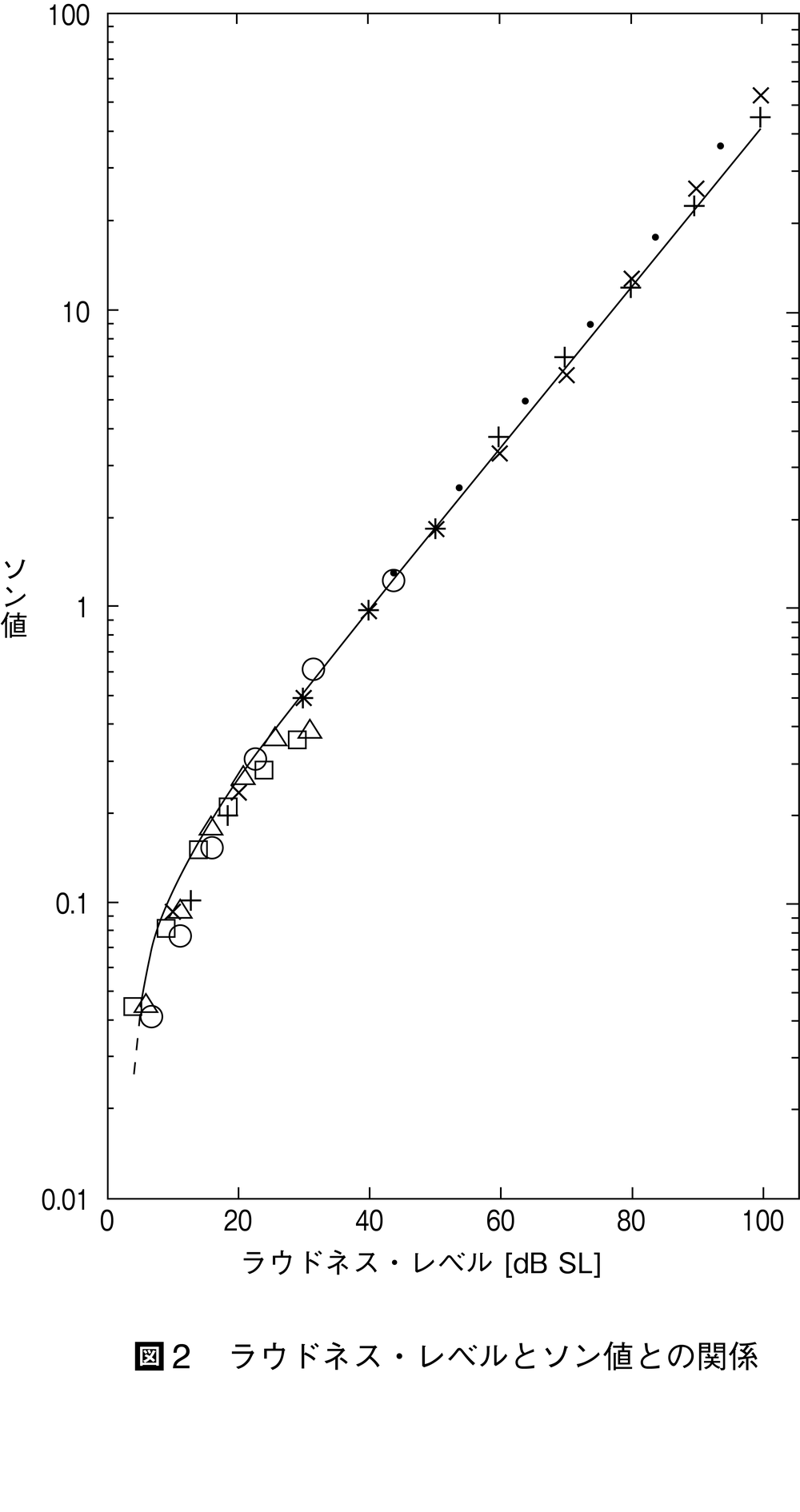
図2 ラウドネス・レベルとソン値との関…

図1 等ラウドネス曲線
出典 最新 心理学事典最新 心理学事典について 情報
Sponsored by ![]()
ある範囲の周波数の音の刺激によって生じる感覚で,視覚が利用できない暗やみや森林,草むらの中で生活する動物にとっては,遠く離れた周囲の状況を知るのに重要な感覚である。例えば動物は,迫ってくる危険を避けたり,種内の交信,雌雄の求愛,交尾などに聴覚を利用する。
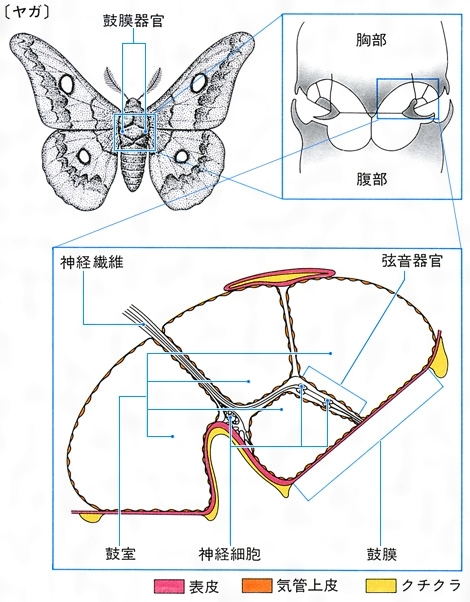
聴覚が発達した動物の一つに昆虫が挙げられる。昆虫のうち,直翅(ちよくし)類のキリギリス,コオロギ,バッタやセミ類のように発音する昆虫では,同種個体間の交信に聴覚が用いられるが,ヤガやカゲロウなどでは,コウモリから逃れるためにコウモリの出す音を聞き分けている。これらの昆虫にはよく発達した聴覚器官である鼓膜器官tympanic organがある。鼓膜器官はキリギリスやコオロギでは前肢脛節(けいせつ)に,バッタやセミでは腹部に,ヤガでは胸部に,そしてカゲロウでは羽にそれぞれ存在する。鼓膜器官の生理的応答から,スズムシやコオロギ,バッタなどが聞くことができる音の周波数(可聴周波数)はヒトの可聴音に近いが,ヤガやヤブキリ,カゲロウなどはヒトが聞き分けられない高周波音(超音波)を可聴音として聞いている。このほかにハエやカは羽音を聞いて応答することが知られている。聴覚器官は触角にあるジョンストン器官Johnston's organである。羽毛状の触角が羽音のような低周波音に共鳴し,これが触角基部のジョンストン器官を刺激する。また,直翅類などの腹部末端にある尾葉の細長い毛すなわち尾毛は低周波音に応答する。例えばスズムシの尾毛は0.3~3kHzの音に応答する。
脊椎動物は例外なく聴覚器官をもっている。魚類の聴覚器官は内耳だけからなり,いわゆる中耳や外耳を欠く。コイやナマズのようなコイ類(骨鰾(こつひよう)類)では中耳の代りにウェーバー器官があり,音はうきぶくろからウェーバー小骨連鎖を経て内耳に伝わる。両生類や爬虫類には中耳がみられるが,外耳はなく,鼓膜が露出している。外耳は哺乳類で発達するが,鳥類にも一部みられる。内耳のうちで聴覚に関係するのは球形囊で,鳥類では球形囊が長くのび,哺乳類ではさらに蝸牛(かぎゆう)管に発達する。これに伴い有毛細胞の数も多くなっている。例えばガマでは有毛細胞の数は数百であるが,哺乳類では数千,とくに聴覚の発達したイルカでは約1万7000もある。
魚類が聞くことができる音の周波数範囲(可聴範囲audible range)は狭い。わりあい広いとされている真骨魚類でも約1kHzまでしか応答できない。可聴範囲内で最も感度よく応答できる周波数を特徴周波数と呼ぶが,例えばキンギョでは特徴周波数は約300Hzである。両生類の可聴範囲は約100Hzから数千Hzであるが,特徴周波数は低周波部分と高周波部分に二つある。その理由は,両生類の有毛細胞が2群に分かれているからである。1群は基底乳頭と呼ばれる所に集まり,1群は両生類乳頭に集まる。例えばガマでは基底乳頭の特徴周波数は約1500Hz,両生類乳頭の特徴周波数は約400Hzである。ガマでは高音も低音も配偶行動に関係するが,小型のアマガエルでは高音は配偶行動,低音は天敵である大型のカエルからの逃避行動に関係するらしい。爬虫類の可聴範囲は約100Hzから数千Hzで,トカゲ類では広いが,ヘビやカメ類は200~700Hzの低周波音によく応答するが,高周波音に対する感度は急に悪くなる。鳥類の可聴範囲はさらに高周波側に広がり,1万2000Hz~1万5000Hzの音までが可聴範囲になるが,哺乳類に比べると狭い。哺乳類の聴覚の可聴範囲は動物のうちで最も広い。一般には可聴範囲の上限は数千Hzで,コウモリでは12万Hz,イルカでは15万Hzまで可聴範囲になる。この点ではヒトは哺乳類のうちでは狭い可聴範囲をもつといえる。
コウモリやイルカなどは,自分が発した音の反響(こだま)を聞いて餌や障害物の定位をしている(反響定位)。小さなものに対して反響定位をするのには,波長の短い高周波音を使わなければ,こだまは返ってこない。コウモリが超音波を発し,これを聞くことができるのには,このような利点がある。コウモリは暗黒中で径0.2mmの金属線を避けて飛ぶことができる。また,接近してくる餌と遠ざかる餌とをこだまの違いで区別できる。
執筆者:立田 栄光
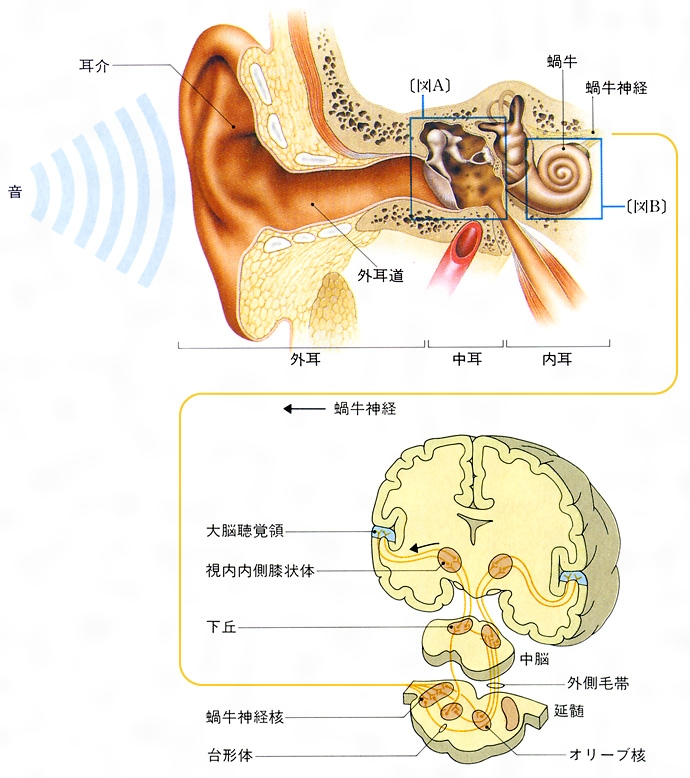
音の感覚は,空気の振動に基づくもので,ほとんどすべての陸生動物と一部の水生動物にみられ,それぞれさまざまな聴覚器(聴覚器官)をもっているが,ヒトを含めて哺乳類は聴覚器(音受容器官)として蝸牛cochleaをもつ。蝸牛は内耳の一部をなすが,そのほかに聴覚に関係するのは,動物が水中から陸上に上がった際に獲得した中耳と,集音のための外耳である。空気中を伝わってきた音波は,外耳で集められ,中耳の鼓膜とこれに続く耳小骨連鎖(哺乳類では,つち骨,きぬた骨,あぶみ骨という3個の耳小骨が関節をつくり,音波の伝播(でんぱ)の能率を高めている)により液体(外リンパ)で満たされた蝸牛へ送り込まれるのである。この機構はH.L.F.vonヘルムホルツにより1863年に明らかにされた。
内耳は蝸牛と平衡器で,ともに骨中に埋没している。ヒトの蝸牛はカタツムリの殻に似た形で,2.7回転ほどらせん形に巻いている。この回転数は動物の種によりいろいろであるが,これは動物により聞く音の範囲が異なることを示している。蝸牛の内部はその中央を基底膜により上下の2階に分けられ,上階は前庭階,下階は鼓室階と呼ばれる。中耳から伝達された音波は,あぶみ骨のはまり込む卵円窓を経て前庭階に伝わり,液体中を通って蝸牛先端に達し,その部にある基底膜の小孔を経て鼓室階を通って蝸牛基底部に戻り,そこの正円窓を通って中耳腔に帰るのである。この間に基底膜に新しい上下に揺れる進行波が生ずる。
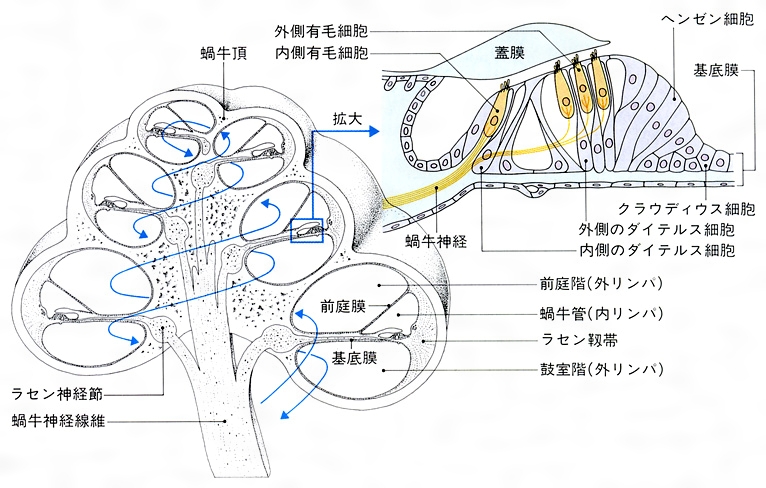
前庭階にはまた,薄いライスネル膜により,断面が三角形の蝸牛管cochlear duct(cochlear canal)がつくられ,これもらせん状に巻いている。中央階とも呼ばれるこの部分は,膜迷路の一部で,内部は内リンパで満たされ,底面をなす基底膜の上には,聴覚器であるコルチ器Corti's organがのっている。コルチ器には感覚細胞である内・外有毛細胞があり,前者は1列,後者は3列に細胞が並び,蝸牛管の全長にわたる。これらの感覚細胞列を支持細胞が囲み,さらに有毛細胞上面は網様膜で囲まれて蝸牛管の壁の一部をつくり,内・外リンパの境界をなしている。小孔で通じる前庭,鼓室の両階を満たすのが外リンパであり,蝸牛管を満たすのが内リンパであるが,この内リンパに浸り,有毛細胞をおおうように蓋膜が張られている。外有毛細胞の毛の一部は蓋膜の中に入り込み,膜運動はそのまま毛の運動として伝達される。内有毛細胞の毛は蓋膜から離れているといわれる。
内耳における音の分析については,古くはヘルムホルツの共鳴説とラザフォードRutherfordの電話説があった。前者は有毛細胞をそれぞれ共鳴器と考えたのに対し,後者は蝸牛は電話器のように働き,音の分析は脳内で行われるというのであったが,ともに実験結果ではなく単なる仮説にすぎなかった。1941年にベケシーGeorg von Békésy(1899-1972)はヒトの死体から内耳を含む頭蓋骨を切り取り,刺激音とわずかに異なる回数で明滅する強い光を用いるストロボ方式によって,音刺激の際の基底膜の運動を緩やかにして観測することに成功した。基底膜の幅は蝸牛先端に向かうにしたがって広くなり,膜の各部の弾性が異なっており,刺激音の周波数が低いほど,膜の上で進行波は先端に到達し,高いときは基底部にとどまることを見た。これがベケシーの進行波説である。その後,今日までに種々のより精巧な方法が考案され,分子運動に近い精度(nm)で測定が生きた運動の蝸牛について行われたが,原則的にはベケシーの観測が誤りでなかったことが認められている。複雑な音では,多くの最高値を示す弾性波が基底膜上に生じ,音波形のフーリエ解析が基底膜上で行われるのである。
一方,ウェーバーE.G.WeverとブレーC.W.Brayは,ネコの中耳の正円窓付近の骨の上から刺激音と同じ形をした電位波形を記録し,これを蝸牛のマイクロホン電位と呼んだ(1930)。その後の研究から,この電位変動は内耳で起こる電気現象であって,有毛細胞群から起こる電位変化と,有毛細胞に接続する蝸牛神経束の活動電位の総和であること(エードリアンE.D.AdrianとデービスH.Davis,1931)が,各種の電極がくふうされて明らかとなり,とくに田崎一二による田崎法(1952)はこの点で功績が大きかった。こうして死体を用いた実験結果は,生体の電気現象と本質的に差のないことが証明できたのであった。一方では内・外リンパの化学的成分でナトリウムイオンNa⁺とカリウムイオンK⁺の比が著しく異なることが明らかとなり(スミスC.Smith,1952),さらにベケシーにより,内・外リンパの間に+100mVに近い電位差があることが見いだされた(1952)。なお,この電位差の源は蝸牛管側壁にある血管条におけるK⁺の分泌電圧であるとされた(田崎ら,1959)。
現在,有毛細胞上面の毛の動きから,上記の電位差による電気抵抗の変化がマイクロホン電位の起源だというデービスの考え(1957,可変抵抗説)が一般に通用している。この問題に関連して内・外有毛細胞の細胞内電位の測定が行われ(ラッセルJ.J.RusselとセリックP.M.Sellick,1978。田中康夫ら,1980。ダロスP.Dallossら,1982),音刺激により直流的脱分極と音波形と同じ交流成分が低周波の際には現れ,1kHzを超えると交流成分がみられなくなることがだいたい容認された(モルモットで)。一方,ウミガメを用いたクローフォードA.C.CrawfordとフェティプレースR.Fettiplaceの実験(1981)では,各有毛細胞は固有の周波数の応答を示すというから,この結果からはヘルムホルツの共鳴説に近い考え方が導きだされる。このような細胞内電位の変化により,毛の運動に一致して細胞底部のシナプスから伝達物質の放出が起こり,蝸牛神経繊維に放電が開始される。しかし,この部位には上位脳から下行する抑制性繊維がきており,両者の総和の影響で神経放電が上位脳に向けて,神経情報として送り出されるのである。
極微小電極の発達から単一有毛細胞のみでなく,単一の聴神経繊維や神経細胞の電気的応答(放電)が記録できるようになり,延髄,中脳,間脳,はては大脳皮質の単一ニューロン(神経繊維と細胞の両者)の応答の記録が可能となり,最近はHRP法(ホースラディッシュ過酸化酵素法)により各繊維の全貌が示されるようになった。蝸牛神経の1側には約3万本の繊維が含まれ,高音から低音へと順序よく並んで上行する。延髄に入ると中継核があり,大部分は反対側に交叉(こうさ)して上行し,残りの繊維も上行中反対側に移り,間脳における中継核に達するまでに全繊維が交叉して,反対側の大脳皮質聴覚野に至る。聴覚野ではとくに言語の理解が行われるが,動物実験では不可能なので,とくに放射性物質を血管中に送り,CT検査法やNMR法が導入されるなど,最近は新しい方法により目覚ましい発展がみられる。
動物の脳でとくに方向感覚の発達したものにコウモリやイルカの例がある。蝸牛にみられた音周波数による配列は,各中継核にもみられ,隣接するニューロン間の干渉の存在もみられるが,ヒトを含めた霊長類の脳における方向感覚はあまり優れたものではないと思われる。
ヒトの聴覚の最重要な問題は,すでに述べた言語の理解である。動物でも音による交信はあるが,ヒトに比べればはるかに簡単である。ヒトの言語音を含む全音域は16~2万Hzで,2000Hz付近が最も感度がよい。3万本の蝸牛神経繊維のなかで1000~2000Hzを担当する繊維の数が最も多いが,母音,子音の理解のためにこの付近の繊維がいちばん多くあることはいうまでもないであろうし,この範囲の繊維の共鳴度が最も鋭いこともいうまでもなかろう。音の強さに対する放電数の変化もいちばん多いと思われる。
他方,聴覚障害を生ずる著明なものは結核の特効薬ストレプトマイシン類である。これらの薬物は,有毛細胞の毛の運動に必要な微量のカルシウムイオンCa2⁺や毛に含まれているリン脂質とよく結合してその作用を妨げることが,日本における研究によって明らかとなった。
→耳
執筆者:勝木 保次
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponsored by ![]()
聴覚は音響刺激によっておこる感覚であって、音響を媒介として、身体より隔たったところのできごとを認知することができる。音響は空気の疎密波であり、外耳、中耳はこの音波を内耳に伝導するための装置である。普通、音波は外耳、中耳を経て内耳に達する(これを空気伝導という)。これに対し、空気の振動が頭蓋骨(とうがいこつ)を経て直接内耳に伝えられるのを骨伝導(こつでんどう)という。聴覚の受容器は、内耳の蝸牛(かぎゅう)にある聴細胞である。
[市岡正道]
(1)耳介・外耳道 俗に耳ともよばれる耳介は、音波を集める働きをしている。また、外耳道は耳介と鼓膜との間にある空間で、音波の強さを減ずることなく、それを鼓膜に伝える。ウマ、イヌ、ネコなどでは、音波刺激によって反射的に耳介を音源の方向に向ける。これをプライヤーの反射〔ドイツの生理学者・心理学者W. T. Preyer(1841―1897)にちなむ〕という。
(2)鼓膜 鼓膜は中耳側にくぼんで張られた漏斗(ろうと)状の膜(厚さ約0.1ミリメートル)である。形状は不整な円形をなし、その緊張も不均一である。さらに中耳側にはツチ骨柄が付着している。こうした形状のため、鼓膜は固有振動数をもたない膜として、広い範囲の振動数の外来音波に対して均等に共振できることとなる。
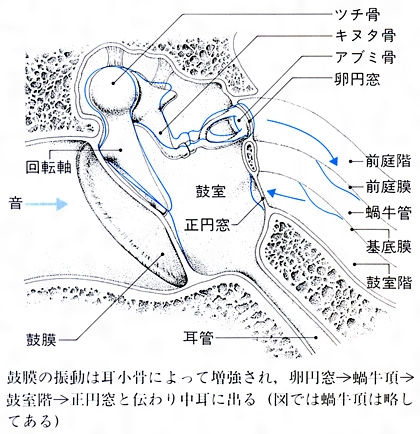
(3)耳小骨 鼓膜の内方には3個の耳小骨(ツチ骨、キヌタ骨、アブミ骨)が関節で連結し、全体として一つの角槓杆(かくこうかん)をなしている。鼓膜の振動は、耳小骨によって、音圧が約20倍に増強されて卵円窓に伝えられる()。
(4)耳管 耳管は中耳腔(くう)と鼻咽頭(びいんとう)を連結しているが、通常は閉じている。しかし、嚥下(えんげ)、そしゃく、あくびなどのときには開き、鼓膜内外の気圧を等しくし、鼓膜の振動を容易にする。
(5)蝸牛 ヒトの蝸牛は螺旋(らせん)状に2回と4分の3回転した管で、基底膜と前庭膜とによって、蝸牛の全長にわたって3階〔前庭階、蝸牛管(中央階)、鼓室階〕にくぎられている。前庭階と鼓室階は外リンパに満たされ、蝸牛管は内リンパに満たされている。基底膜の上には、有毛細胞(聴受容器)を主としたコルチ器(ラセン器)がある。卵円窓に伝えられた振動は、蝸牛頂→鼓室階→正円窓と伝わり、最後には中耳に出るが、この間に基底膜を振動させる。このため、コルチ器の有毛細胞が興奮し、その興奮は蝸牛神経によって中枢に伝えられる()。二次ニューロンは中脳の蝸牛神経核よりおこり、下丘→視床内側膝状体(しつじょうたい)→大脳聴覚領に達している。卵円窓の振動は前庭階の外リンパを振動させるが、これは基底膜に沿って縦方向に進行する外リンパの波を発生させる。この進行波は、低音ほど蝸牛頂に近いところに、また、高音ほど蝸牛頂に遠いところに振幅の最大がみられる。このようにみると、音の高低は蝸牛で解析されていることがわかる。
[市岡正道]
感覚としてわれわれが感受する音は、純音、楽音、騒音に分けられる。純音とは倍音を含まない音(たとえば音叉(おんさ)の音)であり、楽音とは基音と倍音との合成音であり、しかも、音波が反復するというパターンを示す。これに対し、騒音では音波が非周期的であり、反復しない形となる。騒音の強さをデシベル(dB、後述)で表すと、ささやき・20dB、会話・60dB、交通量の激しい所・80dB、走行中の地下鉄・100dB、耳に痛みを感じる音・140dB、飛行中のジェット機・160dBといわれている。
音には大きさ(強さ)、高さ(調子)、音色という三つの性質がある。大きさは音波の振幅と、高さは音波の振動数とそれぞれ関係をもっている。すなわち、だいたいにおいて振幅が大きいほど音は大きく、振動数が多いほど音は高くなる。これらに対し、音色は、その音に含まれている上音の含まれ方によって決められるものである。
音の強さと振動数とを変数としたとき、音の聞こえる範囲を「聴野」という。ヒトは約20ヘルツから約2万ヘルツまでの音を聞くことができ、最低の聴覚閾値(いきち)は、主言語の振動数帯である約200ヘルツから約3000ヘルツの間にあることがわかる。また、音波があまり強いときには、痛さを感じる。さまざまな振動数をもつ純音をイヤホンを通じて聞かせ、その閾値を正常者の閾値のパーセントとして描かれたグラフを聴力図(オーディオグラム)といい、このための装置を聴力計(オーディオメーター)という。
音波の圧力で示された音の強さ(S)は、普通、デシベル(dB)単位で表される。すなわち、S0を閾値の強さとすると、Sは20logSデシベルとなる。S=S0のときは0dBであり、したがって閾値の強さも0dBということになる。なお、閾値の強さS0は、
S0=0dB=0.000204dyne/cm2
と決められている。
高低2音が耳に入るとき、低音によって高音は聞こえにくくなる。これを「音の隠蔽(いんぺい)(マスキング)」という。大脳皮質の聴覚領では、音の調子を感受する部位が一定の配列をなしている。それは、ちょうど蝸牛を聴覚領の上に広げたような配列となっており、低音は前外方で、高音は後内方で感受されるようになっている。また、音源の位置や方向の認知は、音波が両耳に到達する時間差、その結果としての両耳における音波の位相差、および、音源に近いほうの耳では音が大きく聞こえることなどが総合されて初めて可能となる。
[市岡正道]
聴覚は振動覚の一種である。聴覚以外の振動覚や平衡覚など、他の機械的感覚に関係する受容器は、動物界に広く分布する。これと対照的に、聴覚器は脊椎(せきつい)動物と節足動物に知られているにすぎず、比較的遅れて進化した受容器と考えられている。節足動物、とくに昆虫類に発達した聴覚器には、空気や水の動きを直接受容するものと、圧受容器として音圧を受容するものとがある。ゴキブリの尾毛のような触毛(または聴毛)や、ハエやカなど多くの昆虫の触角にあるジョンストン器官は前者に属する。昆虫の鼓膜器官は、表皮と気管壁からできた鼓膜をもった、音圧を受容する真の聴覚器であり、キリギリス、コオロギでは前肢に、バッタ、セミでは腹節に、ドクガ、ヤガでは後胸にある。ヤガの鼓膜器官は、天敵であるコウモリの発する超音波に反応する。脊椎動物の聴覚は、鳥類、哺乳(ほにゅう)類においてよく発達し、同種間の情報伝達のほかに、コウモリやイルカなどの反響定位に活用されている。
[村上 彰]
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponsored by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponsored by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponsored by ![]()
字通「聴」の項目を見る。
出典 平凡社「普及版 字通」普及版 字通について 情報
Sponsored by ![]()
…これらの働きは網膜に入った視覚の刺激が上丘や大脳皮質を経由して起こるものである。(3)視覚や聴覚の刺激による運動反射 これは目の前に飛んできた物体を反射的に避けたり,音の刺激の方向に反射的に頭を向けたりするときに役立っている。いずれも上丘に達した光や音のインパルスが,上丘から出る視蓋脊髄路により頸髄に伝えられることによって起こる。…
…脊椎動物の頭部にある有対の感覚器官で,平衡覚と聴覚をつかさどる。ふつう〈耳の形〉などというときには,哺乳類の頭の両側に突出した耳介を指すが,解剖学的にいえば耳には内耳,中耳,外耳の3部分が含まれる。…
※「聴覚」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponsored by ![]()
夏の暑さに体が慣れること。数日から数十日間で起こる短期暑熱順化と、数年または数世代にかけて起こる長期暑熱順化とがある。→寒冷順化[補説]近年では、冷房設備の普及にともない短期暑熱順化が起こりにくくなっ...