デジタル大辞泉 「葉緑体」の意味・読み・例文・類語
ようりょく‐たい〔エフリヨク‐〕【葉緑体】
Sponsored by ![]()
翻訳|chloroplast
Sponsored by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponsored by ![]()
クロロプラストともいう。藻類と緑色植物に見られる色素体の一つで,光合成の生化学的過程はこの中で進行する。葉緑体は多くの植物では直径5μm前後,厚さ2~3μmの碁石形であるが,藻類には板状,網目状,らせん形,星形,カップ形など特殊な形やひじょうに大きい葉緑体をもつものがある。葉緑体は黄色のカロチノイドのほかに多量の葉緑素(クロロフィル)を含んでいるので緑色に見える。褐藻や紅藻の葉緑体は葉緑素のほかにフィコキサンチンやフィコエリトリンを含んでいるので褐色または紅色に見える。葉肉細胞に含まれている葉緑体の数は通常数十~数百個程度であるが,1個の細胞に数千個以上の葉緑体が含まれる例や,ただ1個だけ含まれている例も知られている。
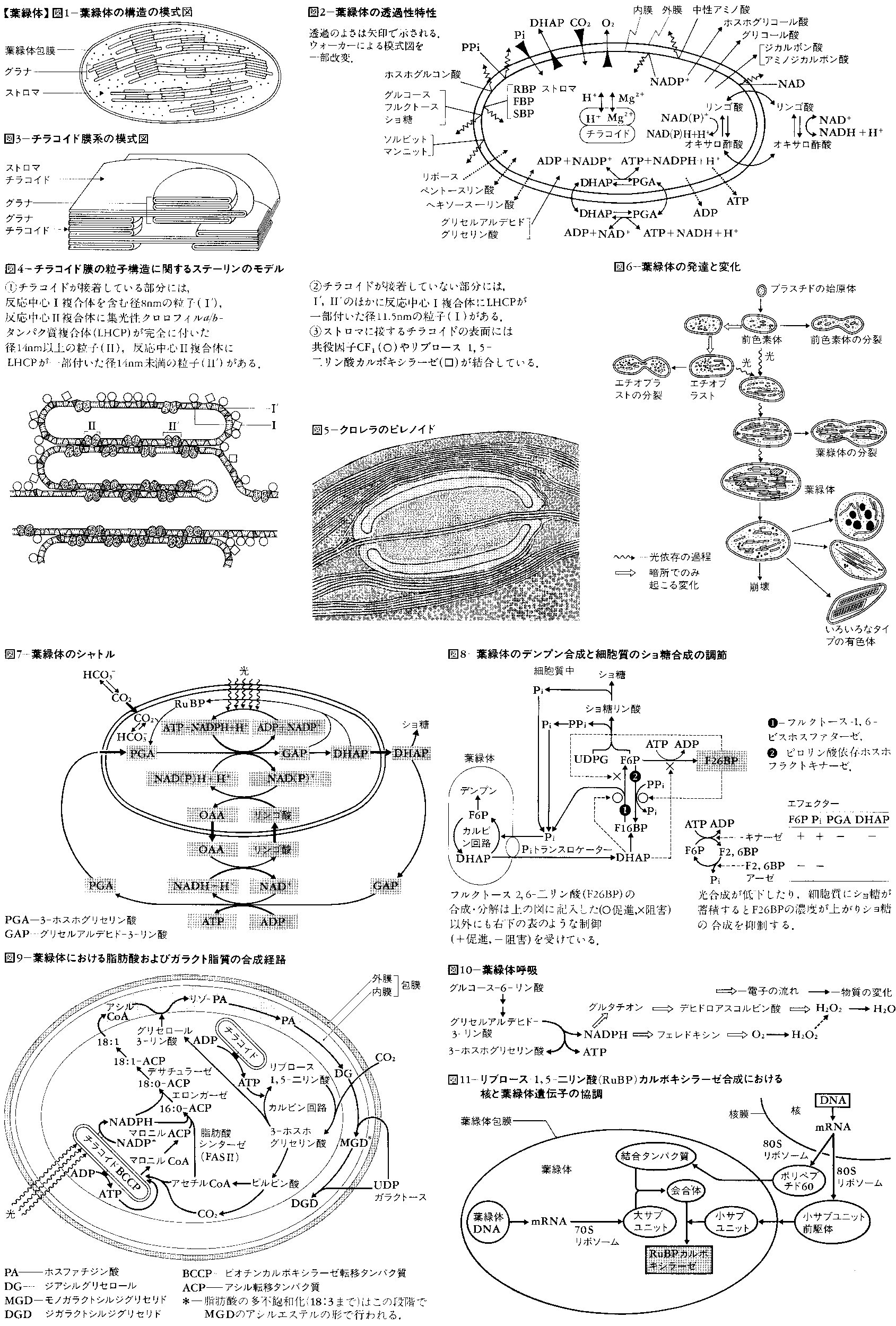
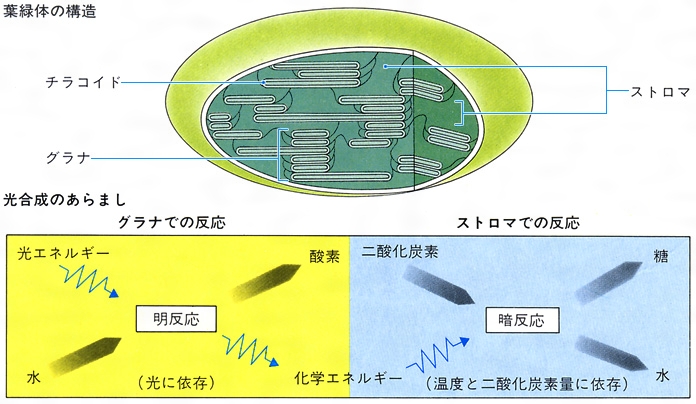
葉緑体の内部構造は,包膜envelopeとそれに包まれた基質(ストロマstroma)およびストロマ中に含まれる膜系(チラコイドthylakoid。かつてはラメラlamellaと呼んだ),その他の構造体に大別される(図1)。
包膜は内膜,外膜の2枚の膜から成り,それぞれイオンや種々の物質に対して選択的透過性をもっている(図2)。
チラコイドは扁平な袋で,成熟した葉緑体では葉緑体の長軸に平行に配向し,チラコイドが折りたたまれ,あるいは小型のチラコイドがコインを積み重ねたような状態になったもの(グラナgranaまたはグラナスタックgrana stack)が,大きなチラコイドの間にはさまっている(図3)。グラナを構成するチラコイドをグラナチラコイド,これをつないでいるチラコイドをストロマチラコイドまたはグラナ間チラコイドという。断面ではチラコイドが積重ねに見える場合でも,チラコイドの内腔は連続しているのではないかと考えられている。近年,チラコイド膜の表面に露出および内部に埋め込まれた粒子の分布がフリーズ・エッチング法(膜の周りの氷を昇華させたとき露出する表面の凹凸を見る)とフリーズ・フラクチャー法(膜内部の疎水性部位に沿って膜が裂け,引きはがされて生じる面の凹凸を見る)により調べられている(図4)。これらと生化学的な実験結果を総合すると,ストロマチラコイドとストロマに接しているグラナの最外層のチラコイドには光化学系Ⅰがあり,その外表面には光リン酸化反応の共役因子CF1,リブロース-1,5-二リン酸カルボキシラーゼやリボソームが結合していると考えられる。グラナチラコイドの積重なりのある部分には光化学系ⅠおよびⅡがある。CF1はチラコイド膜を低イオン強度の液で洗うと膜から離れ,マグネシウムイオンMg2⁺を加えると再び膜に結合し,EDTAまたはシリコタングステン酸も共役因子をチラコイド膜から離すことがわかっている。CF1の配置はADP,無機リン酸がストロマに存在することと符合する。各種の光合成色素や光合成の電子伝達系成分,共役因子などはチラコイドに存在し,光エネルギーの捕捉,電子伝達,ATP生成まではチラコイド上およびその表面近くで進行し,これによって生成したNADPHとATPを用いるCO2固定はストロマ中で行われる。C4植物の葉肉細胞の葉緑体の構造は前述の構造と変わらないが,維管束鞘(いかんそくしよう)細胞の葉緑体は,リンゴ酸(CO2源であると同時に還元力源ともなる)型のものではグラナ構造をもたず,アスパラギン酸(CO2源にしかならない)型ではグラナ構造をもつ。
緑藻,褐藻,紅藻,接合藻,ケイ藻など多くの藻類およびツノゴケのような蘚苔植物の葉緑体にはピレノイドpyrenoidがある(図5)。ピレノイドはストロマ中にあるタンパク質の塊(0.5~1μm)で,5~10nmの粒状物質が密に詰まっている。多くの場合その周りをデンプンなどの同化物質が取り囲み,ピレノイドの中をチラコイドが貫通している。ピレノイドはタンパク質の貯蔵場所と考えられている。
ストロマには葉緑体の重さの60~80%の水があり,ここに種々のイオン,低分子有機化合物,親水性のタンパク質,酵素(カルビン回路のものを含む),リボソーム,RNA,DNAなどが含まれ,またしばしばデンプン粒が見られる。
葉緑体内には直径50~150nmのオスミウムでよく染まる顆粒(かりゆう)(プラスト顆粒plastoglobule)が見られる。この顆粒はプラストキノン,α-トコフェリルキノン,ビタミンKなどのキノン類に富み,少量のカロチノイドその他の脂質を含む。プラスト顆粒は成熟した葉緑体よりも黄化組織のエチオプラストetioplastに多く含まれ,黄化組織が光を受けチラコイドが形成されるときには減少し,緑葉が老化しチラコイド膜系がこわれるとき増大することから,キノン化合物の貯蔵場所になっているものと考えられている。ストロマにはタンパク質やリボソームが少なく電子線をよく透過させる部域があり,そこにDNAと同定される糸状(太さ2.0~2.5nm)または棒状(長さ5~10nm)の構造が見られる。DNAを含むこの部域は核質nucleoplasmまたはゲノフォアgenophoreと呼ばれる。また,ストロマにはフェリチン鉄の塊をタンパク質が取り囲んだ直径約10nmの球状の粒子(フィトフェリチンphytoferritin)が観察されることがある。多くの場合多数集まり大きな塊になっているが,結晶状態になっていることもある。フィトフェリチンはチラコイド膜のチトクロムやフェレドキシンなどの含鉄タンパク質への鉄の供給源または貯蔵場所と考えられている。
葉緑体およびチラコイド膜の物質組成を表1に示す。チラコイド膜はおもに脂質とタンパク質で作られており,糖脂質が多いこと,脂質を構成する脂肪酸の組成では不飽和脂肪酸が多く82%に達すること,リノレン酸が多いことが特徴である。
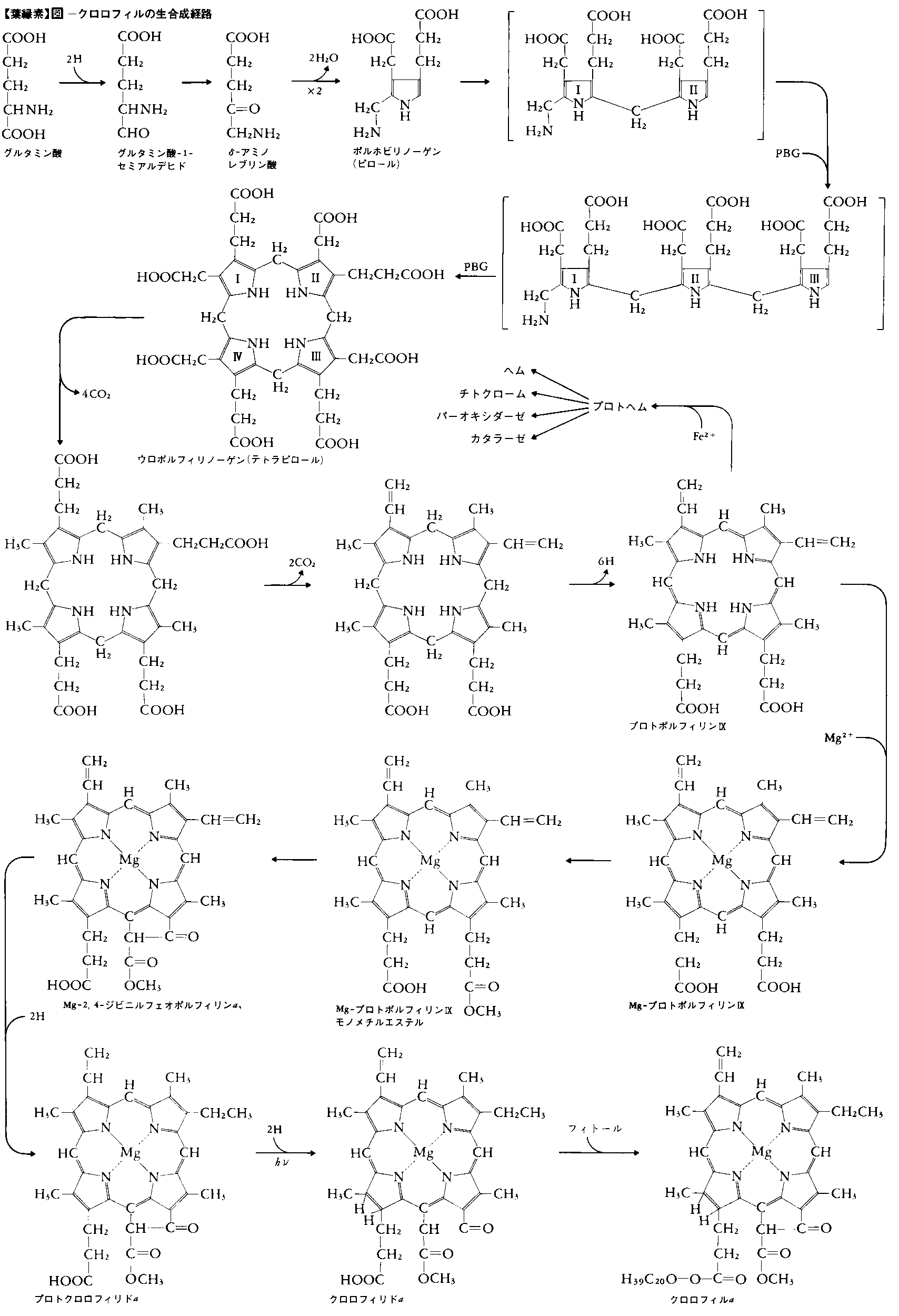
高等植物において,茎の生長点など未分化な細胞の中には,将来葉緑体に発達する未分化な小さな(0.5~1.0μm)色素体(前色素体proplastid)が観察される(図6)。前色素体は0.5μmより小さいプラスチド始原体plastid initialから発達したものといわれ,前色素体の段階になると包膜の内膜の陥入により生じた少量の小胞構造vesicleや未分化な内膜primary thylakoidをもっている。前色素体の形成は明・暗どちらの条件でも進行する。(1)前色素体は光のもとでは,葉緑素の合成と並行して小胞の数が増加し,それが融合しあって大きくなりチラコイドになり,色素体も大きくなり葉緑体となる。(2)植物が暗所に置かれた場合は,葉緑素の合成はプロトクロロフィリドの段階で止まり,内膜構造の発達も小胞の段階でとどまり20~30nmの小胞が集合した直径約1μmの結晶状構造体(前ラメラ体またはラメラ形成体prolamellar body)を形成する。このような色素体をエチオプラストという。これに光が当たると,葉緑素の合成とともに前ラメラ体から放射状に伸びるようにチラコイドが形成されていき,これに伴って前ラメラ体はしだいに消失していき(この状態の色素体をエチオクロロプラストetio-chloroplastという),さらにチラコイドが発達して正常な葉緑体となる。(1),(2)いずれの場合も,グラナ構造は光を受けて短時間後の発達初期の葉緑体にはなく,光化学系Ⅱ活性が出現する時期に形成される。前色素体,エチオプラスト,若い葉緑体は盛んに分裂して増殖する。
緑葉の老化に伴い葉緑体は退化する。すなわちRNA,葉緑体リボソーム,ストロマのタンパク質,葉緑体の厚みが減少し,葉緑素の分解,チラコイドの崩壊が起こり,プラスト顆粒が増加・増大し,葉緑体の収縮,球状化が進行する。葉緑体の発達,退化の制御のしくみはよくわかっていないが,サイトカイニンは葉緑体の発達を促進し,退化を抑制する。有色体chromoplastを含む果実の場合には,果実の成熟時に葉緑体のチラコイドが崩壊し,多量のカロチノイドを含むプラスト顆粒またはカロチノイドの結晶で満たされ有色体となる。
光を受けて光合成を行っている葉緑体は(1)細胞質でのショ糖合成の基質としてジヒドロキシアセトンリン酸を細胞質へ放出するほか,(2)ジヒドロキシアセトンリン酸と3-ホスホグリセリン酸およびオキサロ酢酸とリンゴ酸のシャトルshuttleを使って,間接的にATPを細胞質へ運び出す(図7)。光合成で生じた3炭酸が葉緑体内のデンプン合成に使われるか,細胞質でのショ糖合成に使われるかの割合を制御するしくみとして,最近細胞質の信号代謝産物signal metaboliteすなわちフルクトース-2,6-二リン酸が重要な役割を果たし,葉緑体からジヒドロキシアセトンリン酸(DHAP)の出る量が調節されている(図8)。
葉緑体内で行われる反応は光合成だけでなく,光合成に必要な色素やタンパク質の合成,核酸の合成のほかにも多くの重要な合成反応が行われている。亜硝酸レダクターゼは葉緑体に局在し,窒素同化に必須の過程である亜硝酸還元は光合成の電子伝達の過程で還元されたフェレドキシンを利用して葉緑体内で行われる。脂肪酸およびガラクト脂質の合成系は,葉緑体に局在し,実際に葉緑体内で合成が行われる(図9)。脂肪酸合成は完全な光依存性を示し,光合成反応の過程で生じた3-ホスホグリセリン酸,NADPH,ATPが利用される。アセチルCoAカルボキシラーゼの3成分中,ビオチンカルボキシラーゼ転移タンパク質はチラコイド膜に結合しており,ガラクト脂質合成系は包膜に局在する。また,葉緑体膜系は脂肪酸の多不飽和化の反応系をももち,葉緑体の脂肪酸の多不飽和化は葉緑体自身で行われる。このほか葉緑体はジベレリン,アブシジン酸などイソプレノイド系のホルモンの合成系をももっている。
葉緑体は光のもとでペルオキシソーム,ミトコンドリアと共同で行う光呼吸の最初の部分を分担し,O2消費を伴うグリコール酸の生成を行う。このほか暗所では葉緑体内でのグルコース分解から生じたNADPHを図10のような経路で酸化する系が葉緑体に存在することが明らかにされ,光呼吸と区別して葉緑呼吸chlororespirationと呼ばれている。ただし,光呼吸も葉緑呼吸もATP生成を伴わないと考えられている。
葉緑体を単離してその働きを調べる研究は古くから行われてきたが,包膜はこわれやすく,初期に行われたヒル反応の研究や後に行われた電子伝達反応,光リン酸化反応などの研究には,葉緑体単離操作中に包膜がはがれてしまったもの(クラスIIの葉緑体という)が使われていた。上記の諸反応はクラスⅡ葉緑体でも進行するが,CO2固定反応は包膜をもった完全な葉緑体(クラスⅠの葉緑体という)でないと起こらない。現在では単離方法が改良され,クラスⅠ葉緑体を用いて単離葉緑体によるCO2固定だけでなく,葉緑体DNAの転写,翻訳に関する研究が盛んに行われている。
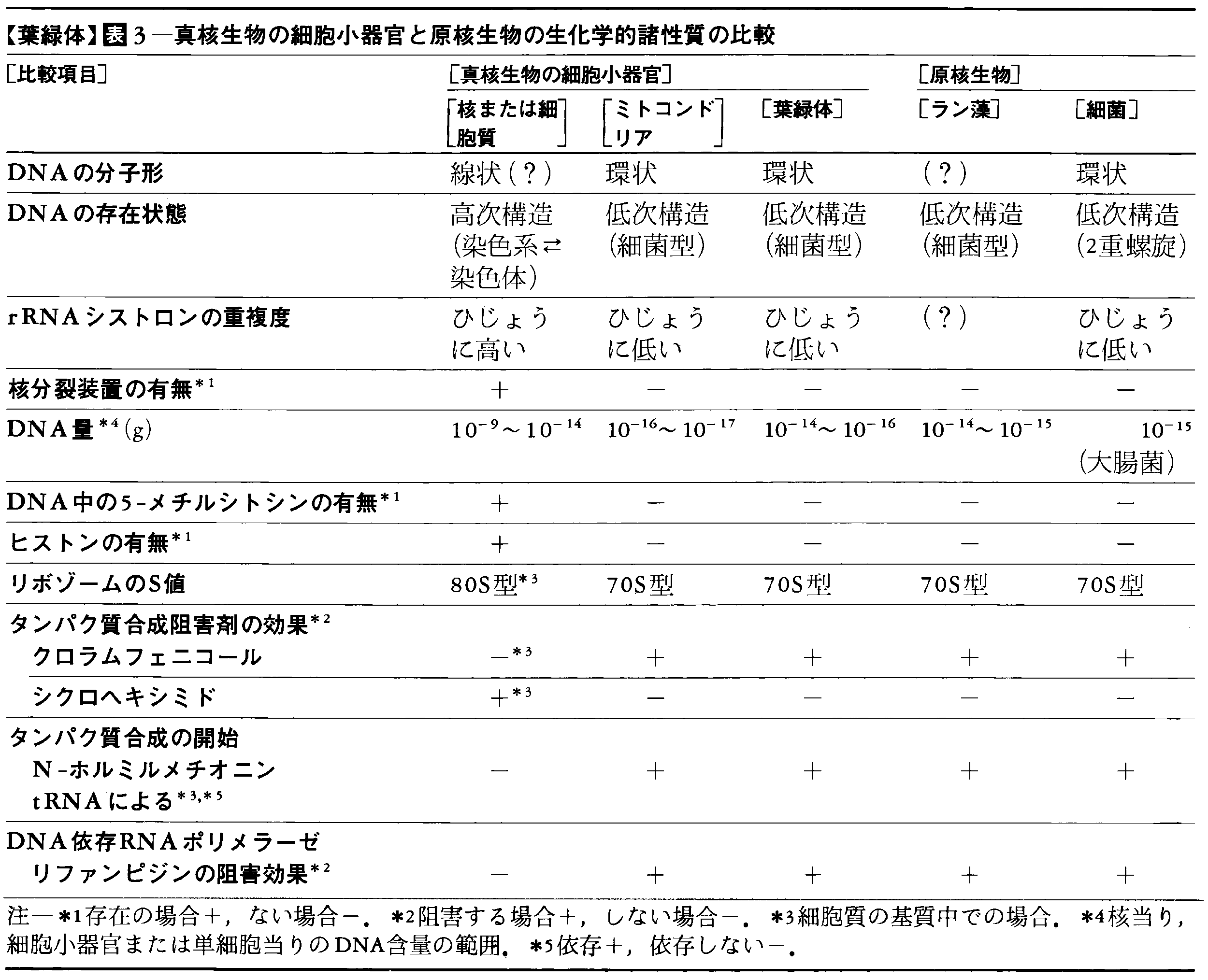
葉緑体DNAは分子量108ドルトン(1ドルトン=1.661×10⁻27kg)で長さ40μmの環状の二重鎖分子で,150~200個の遺伝子をもつことが可能であり,T4ファージのDNAとほぼ同じ大きさである。1個の葉緑体に含まれるDNA量はホウレンソウでは3.8~6.9×10⁻14g,クラミドモナスでは0.5~1.2×10⁻14gで,細菌やラン藻の細胞1個中のDNA量と同じレベルであり,ミトコンドリアのDNA量(10⁻16g)よりずっと大きい。葉緑体1個のDNA含量として10⁻14gという値を使うと,葉緑体1個にDNA分子が約70個(この数をコピー数という)含まれる倍数体である。藻類や高等植物ではコピー数20~100という値が報告されている(表2)。葉緑体DNAの性質は表3に見られるように原核生物のDNAに似ている。現在では葉緑体DNAの物理地図の作られた植物はかなりの数に上っている。葉緑体の遺伝子工学的利用の範囲は今後もますます広がることが予想され,最近では葉緑体DNAを,植物細胞を宿主とするベクターに改変する試みが行われている。
葉緑体は独自のDNAをもち分裂増殖して,細胞の分裂とともに細胞から細胞へと引き継がれていくので,核外自己増殖系の一つに数えられるが,核の支配から完全に独立しているわけではない。葉緑体DNAは葉緑体自身の成分に対する遺伝情報をすべてもっているのではなく,表4に示すように多くの重要な成分は核にコードされている。核DNAにコードされているリブロース-1,5-二リン酸カルボキシラーゼの小サブユニットは葉緑体の包膜を通過して葉緑体内へ入り,葉緑体DNAにコードされているこの酵素の大サブユニットと会合してホロ酵素となる(図11)が,この現象は葉緑体と核の関係を調べるかっこうの例として多くの研究が集中的に行われている。
葉緑体のDNA依存RNAポリメラーゼやリボソームの性質,リボソーム上でのタンパク質合成に関する特徴は原核生物に似ており(表3),また葉緑体のリボソームRNAの塩基配列は細胞質のリボソームRNAよりもラン藻のリボソームRNAに似ている。
進化の過程での葉緑体の起源については,現在二つの説(1)葉緑体の重要な性質の多くが原核生物に似ていること,ラン藻を含む共生体cyanelleが現に存在することなどから,光合成細菌やラン藻の祖先が,もともとは葉緑体をもたなかった宿主細胞と共生した結果生じたとする説(共生説)と,(2)葉緑体の重要な形質の多くが核にコードされていること,原核生物の細胞膜が多くの機能をもつのに比べ,真核生物の膜系は分業化してこれらの機能を別々にもっていること,原核生物としての特徴を一部残したまま,葉緑体またはクロロフィルbをもつ移行型とも考えられる生物が現に存在することなどから,原核生物の細胞膜の陥入によって生じたひだが分化して葉緑体となったとする説(細胞内分化説)とがある。
執筆者:辻 英夫
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponsored by ![]()
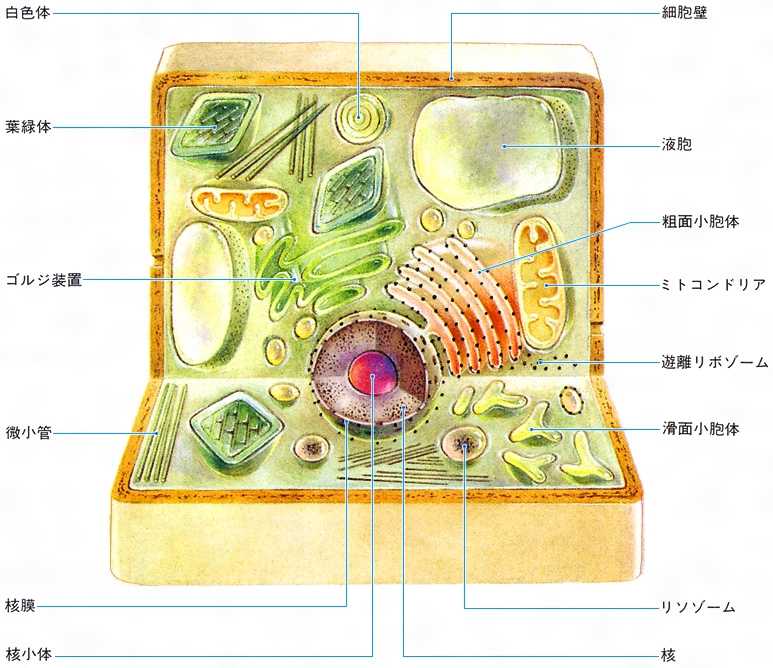
緑色植物の緑色部分の細胞に特有の構造体。大きさが光学顕微鏡で見える水準にあるうえ、クロロフィル(葉緑素)を含んで緑色をなしているのでよく目だつ。陸上植物では直径5マイクロメートル、厚さ1~3マイクロメートルぐらいの凸レンズ形(円盤状)をしているのが普通であるが、緑藻の仲間ではベル形、螺旋(らせん)形、星形、網状のものがあり、分類の目安となる。褐藻類、紅藻類の葉緑体はそれぞれ褐色、紅色にみえるが、これはクロロフィル以外にフコキサンチン、フィコエリスリンなどの色素を含んでいるためである。葉緑体の一細胞当りの数は、種子植物では一定していない場合が多く、だいたい数十個から数百個に及ぶ。原始的な体制の植物ほど一細胞当りの数は少なく、また一定している例が多い。なかには1細胞当りに1個という例もある。
もやしのように、光の当たらない条件下で育った植物では、葉緑体にクロロフィルが形成されず、淡黄色を呈している。これはエチオプラストとよばれる黄色の色素体のためであるが、光の照射を受けると速やかにクロロフィルが合成されて緑化する。
種子植物の根や茎の内部、あるいは斑(ふ)入りの葉の白色部には、エチオプラストのように緑色を欠き、しかも光を与えても緑化しない色素体がある。これを白色体(はくしょくたい)とよぶ。デンプンを貯蔵する器官の細胞では盛んにデンプンの合成が行われる。このような貯蔵デンプンの大きな粒を含む色素体をアミロプラストまたはデンプン形成体とよぶ。ある種の植物の花弁や黄葉では、濃い黄色ないし橙(だいだい)色を呈し、クロモプラストまたは有色体とよばれる色素体がみられるが、これはクロロフィルが失われ、そのかわり黄色、橙色などのカロチノイドが蓄積されたものである。また形成層のように、盛んに分裂を繰り返している未分化の細胞には、白色体に似ているが小形で、分裂によって増殖するものが含まれている。これはプロプラスチド(原色素体、前色素体)とよばれるもので、細胞の分化に伴って葉緑体、白色体、アミロプラスト、クロモプラストなどに分化してゆく能力をもっている(これら全体を総称してプラスチドとよぶ)。
[佐藤七郎]
葉緑体の構造をみると、外表面には二枚の薄い膜(単位膜)からなる包膜がある。包膜に囲まれた内部には、同じ単位膜でできたチラコイドとよばれる扁平(へんぺい)な袋があり、葉緑体の全長にわたって、ほぼ平行に並んでいる。陸上植物の葉緑体では、この大きなチラコイドのところどころに小さいチラコイドが何枚も重なって付着している(これをグラナとよぶ)。藻類などでは、グラナ部分のチラコイドの大きさが不ぞろいであったり、グラナがなく、チラコイドが2~3枚ずつ全面にわたって接着して重層チラコイドをつくっていたりする。このチラコイドの配列の様式は系統と関連があり、分類の指標となる。
チラコイドの周囲を埋めている物質はストロマとよばれ、比較的構造に乏しいが、リボゾーム(RNA‐タンパク質複合体)があり、いろいろの脂質が液滴となって散在している。ストロマには、太さ2.5ナノメートルぐらいのDNAの分子がみられることもある。種子植物と緑藻植物では同化デンプンがこのストロマの中に形成される。
[佐藤七郎]
葉緑体の生理的機能は光合成である。光合成の前段階は、光エネルギーでATP(アデノシン三リン酸)と還元力としてのNADPH(還元型のニコチン酸アミドアデニンジヌクレオチドリン酸)を生成する反応で、これを明反応とよぶ。明反応はもっぱらチラコイドで行われる。生成されたATPとNADPHはストロマ中に放出される。ストロマ中には二酸化炭素をリブロース二リン酸に結合させたうえで、これを二分子のホスホグリセリン酸にする酵素がある。これが炭酸同化作用であり、このときに明反応でつくられたATPとNADPHが消費される。ホスホグリセリン酸はその後やや複雑な経路をたどって六炭糖となり、一部はショ糖として細胞質中に送り出される。他の六炭糖はいったん同化デンプンとなって蓄積され、夜間になって明反応が進行しなくなるとショ糖に変えられて細胞質中に放出される。二酸化炭素が固定されて糖がつくられる一連の反応を暗反応とよぶ。
葉緑体内では、炭酸同化だけではなく、窒素の同化も行われる。根から吸収された硝酸イオンは、葉の細胞の細胞質内で亜硝酸イオンに変えられて葉緑体内に入り、ここで明反応で生成されたATPと還元型フェレドキシンを使ってアンモニアを経てグルタミン酸などのアミノ酸が合成される。このようなアミノ酸合成は白色体やプロプラスチドでも盛んに行われている。この場合はATPとNADPHは明反応以外で生成される。
このように、葉緑体は無機の炭素と窒素を有機化合物に変える役割を果たして、地球上の生命へのエネルギーおよび必要元素供給の元締めをなしている。葉緑体にはDNAがあり、分裂によって増殖し、単細胞の藍(らん)藻に類似した性質があるので、葉緑体は、藍藻が共生して遺伝的に定着したものではないかという「細胞共生進化説」が注目を集めている。
[佐藤七郎]
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponsored by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponsored by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponsored by ![]()
クロロプラストともいう.高等植物および藻類の細胞中に存在する光合成器官であって,その形は星状,環状,板状などの特殊なものもあるが,球状のものでは直径は約数 μm である.内部は光合成サイクルの酵素類を含むストロマ(stroma)の部分と,光合成色素タンパク質および電子伝達系を含む膜状のラメラ(lamella,またはシラコイド,thylakoid)を積層したグラナ(grana)の部分よりなっている.ラメラは厚さが約10 nm,直径約500 nm の袋状のもので,そのなかで光合成の初期光化学反応が行われている.
出典 森北出版「化学辞典(第2版)」化学辞典 第2版について 情報
Sponsored by ![]()
Sponsored by ![]()
…緑色植物の細胞に含まれる葉緑体と,その類縁で色素を含むことで特徴づけられる細胞小器官の総称。動物細胞・菌類に欠けているが,分裂組織など未分化な植物細胞にみられ,大きさ1~3μmの桿(かん)状または長楕円体のクロロフィルをもたない原色素体proplastidから発達する細胞小器官である。…
…この構造はさらに複雑に折りたたまれて,いわゆる染色糸を形成するが,高次の折りたたまれ方はまだ十分に解明されてはいない。真核生物細胞には,核外にもミトコンドリアや葉緑体中に小さな環状DNAが存在する。また細菌のDNA中には,IS(insertion sequence,挿入断片)や二つのISで挟まれたトランスポゾンtransposonという特殊な塩基配列があって,これらは低頻度でDNAの上を飛び移り,挿入や欠失などの突然変異を起こしている。…
…藻類の葉緑体にある核様の小体で,タンパク質としての性質を示し,合成されたデンプンが貯蔵デンプンとなる際その核となる。核様体,デンプン核などともいわれる。…
…藻類の葉緑体(クロロプラスト)に存在する色素。この色素と結合したタンパク質は,ビリタンパク質またはフィコビリタンパク質と呼ばれる。…
※「葉緑体」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponsored by ![]()
政府首脳が外国を訪問した際の会談内容や合意事項を記した外交文書。法的拘束力は持たないが,その内容は両国を事実上拘束する。類似のものに共同発表 joint statementがあるが,これはより記録的な...
7/28 化学辞典 第2版(森北出版)を更新
6/26 日本大百科全書(ニッポニカ)を更新
4/17 デジタル大辞泉プラスを更新
4/17 デジタル大辞泉を更新
2/17 日本大百科全書(ニッポニカ)を更新