デジタル大辞泉 「藻類」の意味・読み・例文・類語
Sponserd by ![]()
翻訳|algae
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
藻類は、系統分類学的には一つのグループ(単系統)ではなく、異なる祖先をもった複数のグループ(多系統)からなる生物群の総称である。古代より「藻」は肉眼的な大きさを有する光合成生物(広義の植物)のうち水中にすむものをさしていたが、現代の生物学では、「藻類」は酸素分子を発生するタイプの光合成を行う生物のうち、陸上植物もしくは有胚(ゆうはい)植物(コケ植物、シダ植物、種子植物)を除いたものと定義される。約6万5000種が知られている。そのなかには植物界のみならず、黄色生物や原生生物、細菌までもが含まれ、大きさもマイクロメートル(1000分の1ミリメートル)単位のピコプランクトンから数十メートルに達する巨大海藻まで幅広く、色も形態も生殖方法も生活環も生育場所も著しく多様である。
[北山太樹 2025年4月15日]
地球上に最初に現れた藻類は原核生物prokaryotesの藍藻(らんそう)(藍色細菌、シアノバクテリアcyanobacteria)であった。現在知られているすべての生物の最終的な共通祖先(LUCA(ルカ):Last Universal Common Ancestor)が誕生したのは約40億年前であるが、最初の10億年間に光合成を行っていたのは紅色細菌や緑色硫黄(いおう)細菌のようなバクテリオクロロフィルをもつ酸素非発生型光合成細菌のみで、そのため原始大気には酸素分子がほとんど含まれていなかった。そのような嫌気的環境では、生物が大型化や多細胞化するのに充分なエネルギーを得ることは望めない。約30億年前に出現した藍藻は、クロロフィルを獲得し、水を分解して酸素を発生するタイプの光合成を行う能力を獲得した最初の生物である。この酸素発生型光合成は、緑色硫黄細菌がもつ反応系に似た光化学系Ⅰと紅色細菌のそれに似た光化学系Ⅱが連結されたもので、遺伝子の水平移動によって成立したと思われる。藍藻の登場は、大気と海洋の酸素濃度を徐々に高め、オゾン層を形成して紫外線を遮り、海水を好気的にするなど地球環境を劇的に変え、約20億年前に原核生物のなかから出現した真核生物eukaryotesの生存とその多様化を可能にした。世界各地に産するストロマトライトstromatoliteは、真核生物が現れる前に繁栄した藍藻の化石である。また、多様な単細胞性真核生物の細胞内に藍藻が色素体(葉緑体)として共生したことで、多種多様な真核藻類が生まれることになった。葉緑体に含まれるDNAも、それがかつて独立した生物(藍藻)に起源をもつことを示している。
[北山太樹 2025年4月15日]
現代の生物分類は形態のみならず遺伝子の系統解析に基づくようになり、真核藻類の分類学的位置も大きく変動している。生命樹を古細菌(アーキアArchaea)、真正細菌(バクテリアBacteria)、真核生物(ユーカリヤEukarya)の3ドメイン系で構成する分類体系によると、ドメイン真核生物は、ドメイン古細菌のアスガルド古細菌Asgard archaeaに由来し、ドメイン真正細菌のプロテオバクテリアproteobacteriaを細胞内共生させて成立したミトコンドリアmitochondriaと、核膜で包まれた細胞核をもつ生物である。かつて真核生物全体の分類は、細胞学的あるいは生態学的観点に基づき、界kingdomのレベルで行われた時代が長く続き、20世紀中、真核藻類は植物界Kingdom Plantaeや原生動物界Kingdom Protozoa、あるいは原生生物界Kingdom Protistaのなかに置かれてきた。分子系統解析が盛んになる以前、もっとも有力な定説として扱われたリン・マーギュリスLynn Margulis(1938―2011)の五界説(1982)においても、藍藻(らんそう)(原核生物界Kingdom Moneraに分類)以外のすべての真核藻類が原生生物界Kingdom Protoctistaに押し込められた。しかし、近年の分子遺伝学的解析技術の進歩は、真核生物間の系統関係を深いレベルで明らかにしつつあり、界を超えるランクのスーパーグループsupergroupで議論されるようになっている。その結果、原生生物界は解体・消滅し、真核藻類はいくつものスーパーグループに離散することになった。
今日、真核生物の分類は九つ前後のスーパーグループ、すなわちアメーボゾアAmoebozoa(アメーバ生物)、オピストコンタOpisthokonta(後方鞭毛(べんもう)生物)、エクスカバータExcavata、アーケプラスチダArchaeplastida(一次植物)、クリプチスタCryptista、ハプチスタHaptista、ストラメノパイルStramenopiles(不等毛生物)、アルベオラータAlveolata、リザリアRhizariaなどに再編され、真核藻類は7スーパーグループ12門46綱に所属する(表 藻類の高次分類)。この分類体系も定説というわけではなく、毎年のように修正が試みられ、いまだ不安定である。真核藻類の主要なグループを例にあげると、ミドリムシはエクスカバータ、紅藻・灰色藻・緑藻・車軸藻はアーケプラスチダ(一次植物)、クリプト藻はクリプチスタ、ハプト藻はハプチスタ、褐藻・珪藻(けいそう)・黄緑藻はストラメノパイル、渦鞭毛藻(うずべんもうそう)はアルベオラータ、クロララクニオン藻はリザリアに分類される。ただし、エクスカバータ、ストラメノパイル、アルベオラータ、リザリアには藻類でないものが数多く含まれる。なお、藻類の高次分類の表では、ストラメノパイル、アルベオラータ、リザリアの3系統群が SAR(Stramenopiles+Alveolata+Rhizaria)にまとめられている。
[北山太樹 2025年4月15日]
古くから知られる主要な藻類には、伝統的に色を冠した分類群名をもつものが多い。1836年、ウィリアム・ハーベイWilliam Henry Harvey(1811―1866)は体色に基づき海藻を「紅藻Rhodospermeae」、「褐藻Melanospermeae」、「緑藻Chlorospermeae」の3綱に分類した。当時は人為分類と批判されたこともあったが、その後、藻類の体色が光合成色素の組成によるもので系統進化を反映していることが認識され、この形質は現在の分類体系のなかに生かされている。
藍藻(らんそう)blue-green algaeは基本としてクロロフィルaのみを有し、藻種によってはクロロフィルb(原核緑藻、プロクロロン)やクロロフィルd(アカリオクロリス)、クロロフィルf(ストロマトライトから発見)をあわせもつ。紅藻red algaeや灰色藻のクロロフィルはクロロフィルaのみで、補助色素のフィコシアニンやフィコエリスリンも藍藻と類似することから、それらの色素体と藍藻との関係が早くから議論されていた。今日では紅藻や灰色藻の色素体(葉緑体)は、前方に2本の鞭毛(べんもう)をもった原生動物が藍藻の1種を細胞内共生させたものであると考えられている。実際、紅藻の色素体は二重の膜をもち、それらは藍藻と紅藻の細胞膜に由来する。灰色藻の色素体は2枚の細胞膜の間に細菌の細胞壁が残っており、藍藻だったときの名残(なごり)と考えられている。このような原始的な色素体はシアネレcyanelleとよばれる。こうした藍藻の最初の細胞内共生(一次共生)によって色素体が成立した生物、すなわち灰色藻と紅藻、そしてクロロフィルaに加えてクロロフィルbを有する、緑藻green algaeから陸上植物までの緑色植物が一次植物(植物界)である。ただし、ケルコゾア門有殻糸状根足虫綱のパウリネラ・クロマトフォラPaulinella chromatophoraもシアネレをもつことが知られており、色素体の一次共生が実は一度限りの現象ではなかったことを示している。
一方、褐藻brown algaeや珪藻(けいそう)diatomsに代表されるストラメノパイルや、クリプト藻を含むクリプチスタ、ハプト藻を含むハプチスタ、渦鞭毛藻(うずべんもうそう)を含むアルベオラータは、色素体が黄色でクロロフィルaとクロロフィルcを有することから、かつてこれらを一つにまとめるクロミスタ界Chromistaが設けられたことがあった。その多くがクロロフィル以外にもフコキサンチンなど細胞内で赤色を呈する補助色素をもち、体色が黄色になる点で共通するので黄色生物ともよばれる。なお、クロミスタ界からクリプチスタとハプチスタを除外し、リザリアを加えたのが前述のSARである。リザリアのクロララクニオン藻chlorarachniophyteはクロロフィルaとクロロフィルbを含む色素体をもつ。ところで、一次植物の色素体(葉緑体)が二重の膜をもつのに対し、クロミスタ界やSARのほとんどの生物が四重膜の色素体をもつ(アルベオラータは三重膜あるいは色素体をもたない)。クロララクニオン藻の色素体も四重膜である。このことは黄色生物が紅藻を、クロララクニオン藻が緑藻を、細胞内共生させることによって色素体を獲得したことを意味している。その傍証とされているのが、クリプト藻やクロララクニオン藻の色素体内部にみられるヌクレオモルフnucleomorphの存在である。ヌクレオモルフは色素体の内側から二番目の膜(紅藻または緑藻の色素体膜由来)と三番目の膜(紅藻または緑藻の細胞膜由来)の間にみられる細胞小器官であるが、DNAを有していることから、細胞内共生(二次共生)で取り込まれた単細胞性藻類の細胞核の痕跡(こんせき)と推定されている。ミトコンドリアの共生が生物の歴史上一度だけだったのに対し、色素体の共生は複数の系統で何度も起きていることになる。
渦鞭毛藻に至っては、二次共生生物であるクリプト藻、ハプト藻、珪藻を取り込んだ三次共生の例が知られている。また、一次植物の緑藻を二次共生させた渦鞭毛藻もある(渦鞭毛藻自体が二次共生生物なので「連続的二次共生」とよばれている)。細胞内共生の繰り返しが、藻類の光合成色素の多様性を生んでいるといえる。
[北山太樹 2025年4月15日]
藻類の体は多様性に富む。原核生物の藍藻(らんそう)は、真正細菌と同じペプチドグリカンを含む細胞壁をもち、核膜、ミトコンドリアや色素体などの細胞小器官を欠いている。それに対し、真核藻類の細胞は核膜、ミトコンドリア、色素体を含み、大部分がセルロース繊維の細胞壁をもつ点で共通する(分類群によっては、細胞壁の組成やその外被構造に特殊なものがある)。紅藻の細胞壁には寒天やカラギーナンcarageenan、褐藻ではフコイダンfucoidanやアルギン酸が埋め込まれている。また、紅藻、緑藻、褐藻などには炭酸カルシウムの外層が形成される分類群(石灰藻)がある。ハプト藻の細胞を覆う炭酸カルシウムの鱗片(りんぺん)は古来、円石coccolithとよばれている。珪藻(けいそう)は珪酸質の被殻frustuleを上下2個組み合わせた外被をもつ。
藻類には、鞭毛(べんもう)を運動させて泳ぐ多様な遊走細胞がみられる(ただし、藍藻と紅藻の細胞は鞭毛をもたない)。緑藻植物は遊走細胞の前方に等長の鞭毛をもち、平泳ぎのように進む。褐藻などストラメノパイルは、側面から出る不等長の2本の鞭毛をもつ(珪藻は1本)。前方を向く長鞭毛にはマスチゴネマmastigonemaとよばれる構造が羽根状について推進力を生んでいる。クリプト藻は前方に両羽根と片羽根の不等長2本の鞭毛をもつ。ハプト藻は前方に等長2本の鞭毛と1本のハプトネマhaptonemaとよばれる構造をもつ。渦鞭毛藻(うずべんもうそう)には、横溝に体を1周する横鞭毛と縦溝に沿って伸びる縦鞭毛をもつ。ユーグレナ藻は、前方に長い鞭毛と痕跡(こんせき)的な短鞭毛の2本をもつ。
藻類の多くが単細胞性(あるいは群体性)であるが、多細胞性の分類群も複数の系統にみられ、単細胞から多細胞への進化が系統ごとに独立して起こっていることがわかる。とくに緑藻、紅藻、褐藻に多細胞性の種が多く含まれる(褐藻には単細胞性種が知られていない)。褐藻には大型化に成功した種が多く、北海道産のナガコンブは最長で20メートルの長さに育ち、北米でジャイアントケルプとよばれているオオウキモは60メートルに達する海洋最大の生物である。
巨視的生物のほとんどが多細胞生物であるが、藻類のなかには細胞分裂を行わずに単細胞のまま巨大化に成功した生物がいる。ミルやイワヅタ類などの海産管状緑藻は、核分裂のみを繰り返して成長する多核細胞体coenocyteで、数メートルの体をつくる世界最大の単細胞生物である。
[北山太樹 2025年4月15日]
藻類には、生物の生殖方法と生活環のほぼすべてのパターンがみられる。後生動物metazoaや陸上植物land plantsが、大きくて不動の卵と小さいが鞭毛(べんもう)をもち運動性のある精子(精核)によって受精を行う卵生殖oogamyに限られるのに対して、藻類には卵生殖だけでなく、雌雄の配偶子gameteが同形同大で運動性がある同形配偶isogamyや、大きさや形に著しい差異があるものの雌雄ともに運動性のある異形配偶anisogamyが観察される。たとえば、クラミドモナス(緑藻)とシオミドロ(褐藻)などが同形配偶、ミル(緑藻)とムチモ(褐藻)などが異形配偶、シャジクモ(車軸藻)やワカメ(褐藻)などが卵生殖を採用しており、配偶様式の進化が各系統群で独立して起きていると考えられる。
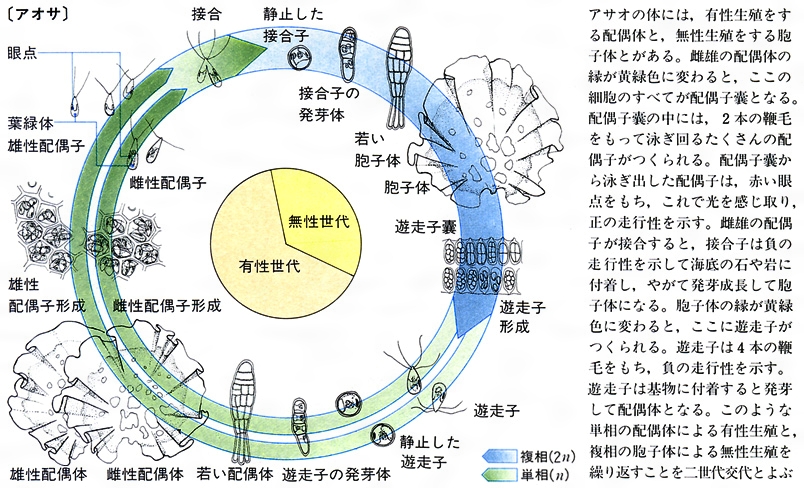
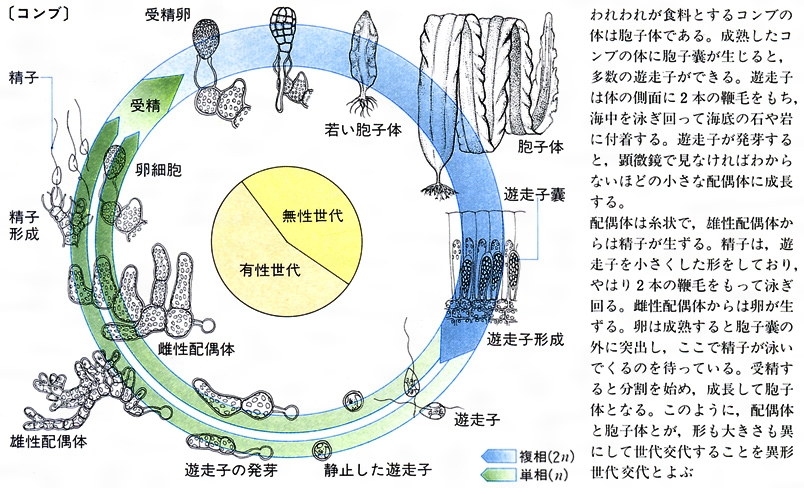
生活環life cycleは生物の一生を環状につないだ概念図で、二倍体の胞子体sporophyteで減数分裂が起きて無性の遊走子(n)を生じ、遊走子が発芽後、一倍体の配偶体gametophyteに発生・成長して雌雄の配偶子(n)が形成され、両配偶子が接合・受精すると二倍体の胞子体(2n)に戻るのが基本パターンである。後生動物では、すべての分類群で減数分裂後に配偶子がつくられるので、二倍体(2n)の配偶体だけが現れ、世代交代がみられない。種子植物の配偶体は非常に小さな体になり(花粉と胚(はい))、見かけ上は世代交代を欠いている。これらに対し、藻類ではあらゆるパターンの生活環をみることができる。アナアオサ(緑藻)やアミジグサ(褐藻)では、同形同大の胞子体(2n)と配偶体(n)が交互に現れる同形世代交代alternation of isomorphic generationsが行われる(図 藻類の生活環〈アナアオサ〉)。マコンブ(褐藻)やヒトエグサ(緑藻)では、胞子体(2n)と配偶体(n)の大きさが著しく異なり、かつては別の生物と考えられたケースが少なくなかった。マコンブのように胞子体が大きく数メートルに達し、配偶体は数ミリの糸状体になる場合と、ヒトエグサのように配偶体が大きい場合があるが、どちらも異形世代交代alternation of heteromorphic generationsとよばれる(図 藻類の生活環〈マコンブ〉)。また、ミル(緑藻)やヒジキ(褐藻)は配偶体を欠き、減数分裂直後に雌雄の配偶子を放出するので後生動物と同じパターンの生活環をもつ。さらに、マクサで代表される紅藻は、鞭毛がなく運動性のない不動精子spermatium(n)と卵細胞である造果器carpogonium(n)が受精した直後に個体発生を行わず、受精卵をそのままの状態で大量に複製して果胞子体carposporophyte(2n)を形成する。その後、第三の世代ともいうべき果胞子体から放出された果胞子carpospore(2n)が発芽すると四分胞子体tetrasporophyte(2n)が発生する。そして、減数分裂後に放出される四分胞子tetraspore(n)からは雌雄の配偶体(n)が発生する。この特異な生活環は、鞭毛を欠き、遊走細胞をもたない紅藻特有のもので、受精卵のクローンをつくることで受精確率の低さをカバーする効果があると推測されている(図 藻類の生活環〈マクサ〉)。
[北山太樹 2025年4月15日]
藻類は海と陸のほぼ全域に分布し、生育場所も太陽光が届くかぎり、海底、海中、海表面、河川、湖沼にとどまらず、汽水、湿地、温泉、氷雪、流氷、土壌、岩石、大気や、コンクリートなどの人工物にも生育する。さらにはほかの生物の体内からもみつかる。
海には、大型の底生藻類、いわゆる海藻benthic marine algae、seaweedsと、浮遊性の植物プランクトンphytoplanktonがすむ。海藻は、おもに潮下帯subtidal zoneの太陽光が届く深さまでの海底に生育するが、干潮時に陸となる潮間帯intertidal zoneにも多く生育する。岩盤や小石、貝や亀などの動物の体表面、ほかの海藻やコンクリートのような人工物などの上にも着生する。イワヅタ類(緑藻)などは、砂地にストロンstolonを這(は)わせて生育する。また、ほかの海藻の体内や動物体内に内生する藻もいる。造礁サンゴの多くは褐虫藻zooxanthellaeを細胞内に共生させて光合成を行わせているが、その正体は渦鞭毛藻(うずべんもうそう)である。海洋に浮遊する植物プランクトンは、大きさで分類される。0.2~2マイクロメートルのピコプランクトンpicoplanktonに分類されるプロクロロコッカス(藍藻(らんそう))は原核緑藻で、地球上の光合成生物として最大の生物量と推定されている。2~20マイクロメートルのナノプランクトンnanoplanktonには、ココリスで知られる円石藻(ハプト藻)が含まれる。20~200マイクロメートルのマイクロプランクトンmicroplanktonには、珪藻(けいそう)や渦鞭毛藻が多い。20センチメートルを超えるメガプランクトンmegaplanktonには、アカモク(褐藻)などホンダワラ類の流れ藻が該当する。大西洋のサルガッソー海(藻海)に浮遊する主要種も、ホンダワラ属の一種Sargassum natansである。また、河口など汽水域を好む藻類も少なくない。そこにはアオノリ類(緑藻)、アヤギヌ類(紅藻)などが生育する。
河川や湖沼などには、淡水に適応した藻類が生育する。大型の藻ではカワノリ、シャジクモ、アオミドロ、マリモなどの緑藻のほか、カワモズク、タンスイベニマダラ、チスジノリなどの紅藻や褐藻のイズミイシノカワなどが知られている。微細な淡水藻類では、アオコをつくる藍藻、オオヒゲマワリ(ボルボックス)、ミカヅキモ、クンショウモなどの緑藻、ハネケイソウ、タルケイソウなどの珪藻や渦鞭毛藻などが生育する。温泉からは紅藻のイデユコゴメなどが発見されている。雪や氷の上からは、赤雪をつくるクラミドモナス(緑藻)などの氷雪藻がみつかる。流氷の下に繁茂するアイスアルジーice algaeは藍藻や珪藻が主である。また、池に生息する繊毛虫ミドリゾウリムシの細胞内には、クロレラ(緑藻)が共生する。
これら水環境以外にも、大気中(気生藻)や岩石中(岩石藻)からも藻類はみつかる。地衣類の体内には藍藻やトレボウクシア藻などが共生し、藻類層を形成している。後生動物でもイヌやネコ、ウシ、人体に寄生して人獣共通感染症を引き起こす病原性の藻類として、トレボウクシア藻のプロトテカProtothecaが知られている。
[北山太樹 2025年4月15日]
日本には、およそ1500種の海藻が分布する。おもに紅藻約900種、褐藻約340種、緑藻約260種からなり、そのうち60種以上が食用とされている。日本の海藻の水平分布は海流の影響を強く受けており、たとえば親潮寒流(千島海流)が支配する北海道太平洋沿岸にはマコンブやナガコンブなどのコンブ類(褐藻)がつくるコンブ藻場が優占し、エゾイシゲ、ヒバマタ、マツモ(褐藻)やダルス、フジマツモ(紅藻)などが生育する亜寒帯性海藻相である。それに対し、本州・四国・九州の黒潮暖流(日本海流)に洗われる太平洋沿岸と、対馬暖流が流れる日本海沿岸には、アカモク、オオバモク、ホンダワラ、ヒジキなどのホンダワラ類(褐藻)やアラメ、カジメ、ワカメ(褐藻)などによる海中林が優占し、ウミウチワ(褐藻)、マクサ、ヒラクサなどのテングサ類やムカデノリ類、コトジツノマタ、トサカノリ(紅藻)、アオサ類、ミル類(緑藻)などがよく生育する温帯性海藻相である。一方、日本海沿岸にはツルアラメやフシスジモクやスギモク(褐藻)などといった日本海特産種が知られている。また、南西諸島や小笠原諸島などは、ラッパモク(褐藻)やコナハダ類(紅藻)などのほか、カサノリ、イワヅタ類、サボテングサ類などの緑藻が多く分布する亜熱帯性海藻相である(図 紅藻植物のおもな種類①・②、褐藻植物のおもな種類①~③、緑藻植物のおもな種類)。
[北山太樹 2025年4月15日]
『堀輝三編『藻類の生活史集成 第2巻(褐藻・紅藻類)』(1993・内田老鶴圃)』▽『堀輝三編『藻類の生活史集成 第1巻(緑色藻類)』(1994・内田老鶴圃)』▽『吉田忠生著『新日本海藻誌――日本産海藻類総覧』(1998・内田老鶴圃)』▽『千原光雄編『バイオディバーシティ・シリーズ3 藻類の多様性と系統』(1999・裳華房)』▽『井上勲著『藻類30億年の自然史――藻類から見る生物進化・地球・環境』第2版(2007・東海大学出版会)』▽『渡邉信監編『藻類ハンドブック』(2012・エヌ・ティー・エス)』▽『矢﨑裕規・永宗喜三郎・島野智之ほか編著『原生生物学事典』(2023・朝倉書店)』
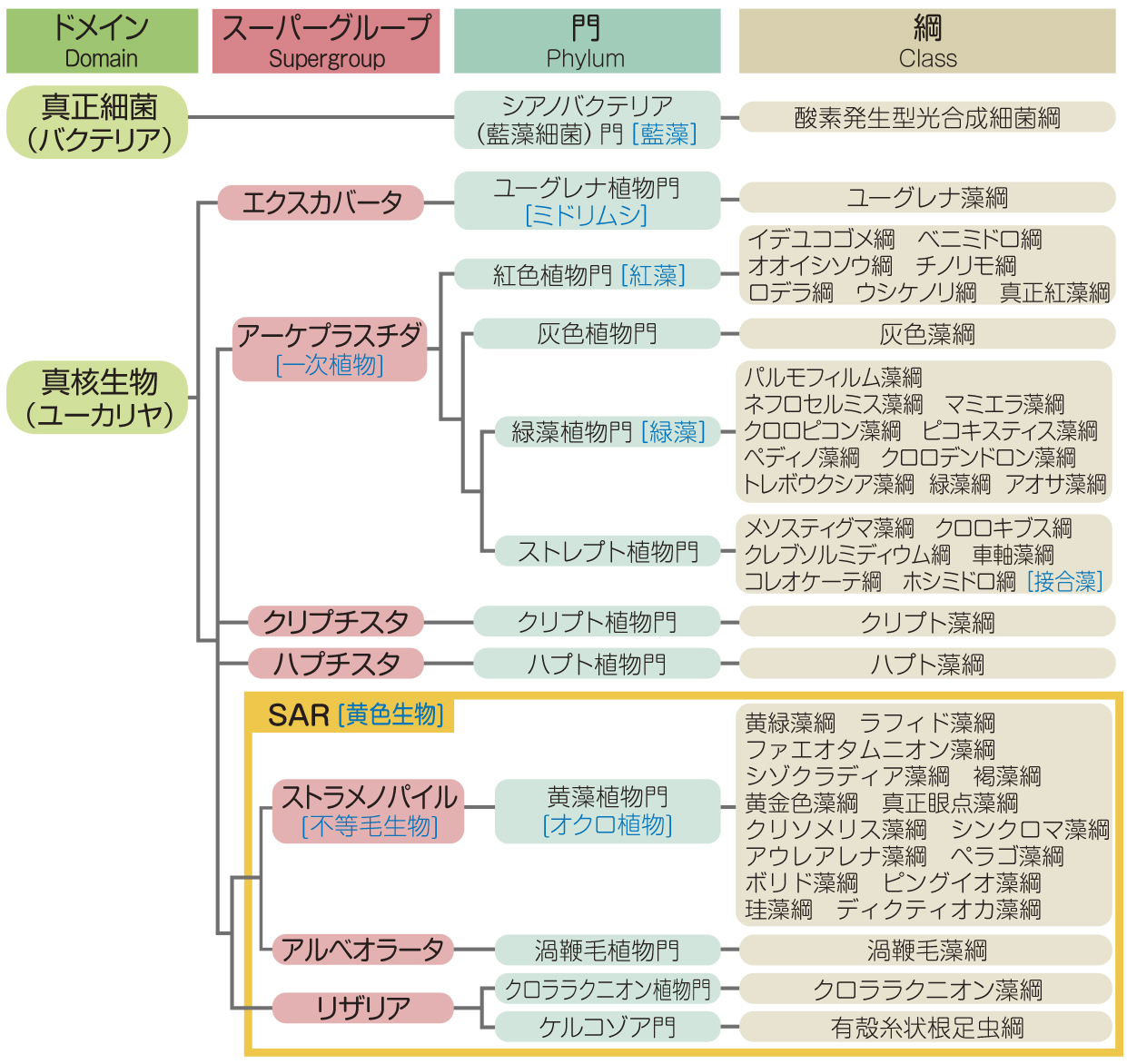
藻類の高次分類
藻類の生活環(アナアオサ)
藻類の生活環(マコンブ)

藻類の生活環(マクサ)

紅藻植物のおもな種類①〔標本画・標本写…

紅藻植物のおもな種類②〔標本画・標本写…

褐藻植物のおもな種類①〔標本画〕

褐藻植物のおもな種類②〔標本画・標本写…

褐藻植物のおもな種類③〔標本画〕

緑藻植物のおもな種類〔標本画〕
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
一般に藻類というと,水中生活を営む体制の単純な植物を指すが,系統的には多様な生物群の集りで,厳密に定義すると,光合成の過程で酸素を放出する光合成生物からコケ植物と維管束植物を除く残りのすべてということになる。しかし光合成色素を欠くものでも体制や生殖方法が類似していれば,藻類として扱われる。
藻類の生育場所はほとんどが水中,つまり海,汽水,および淡水である。海では太陽光の届く大陸棚の深さまでであり,それ以下の深海では育たない。ちなみに海藻で知られた最深の生育記録は199m(褐藻ツルアラメ)である。緑藻のクロレラや黄緑藻のフウセンモのように,土壌中や土壌の表面に生育するものや,緑藻のスミレモやクロロコックムのように,岩上や樹木の表面に生育するものもある。またラン藻のネンジュモのある種のように,ソテツやツノゴケなど他の植物の組織内に生育するものもある。地衣類が,子囊菌と緑藻類またはラン藻類との共生体であることはよく知られている。緑藻類をもつ地衣類はやや緑色がかり,かたい手ざわりであるが,ラン藻類をもつものは黒色がかり,やわらかい手ざわりである。藻類の中には80℃以上にもなる高温の温泉水が流れるところに好んで生育するものがあり,またこれとは対照的に雪の中や氷の割れ目などに生育するものもある。前者は温泉藻と呼ばれ,ラン藻のシネココックス属やユレモ属などのある種や紅藻イデユコゴメがその仲間であり,後者は氷雪藻と呼ばれ,緑藻のクラミドモナスがその代表である。
水中の藻類には,岩や石またはその他の基物について生育するものと,水中に漂って生育するものとがある。基物について生育するものを底生藻または着生藻といい,漂って生育するものを浮遊藻という。浮遊藻はふつう植物プランクトンと呼ばれる。
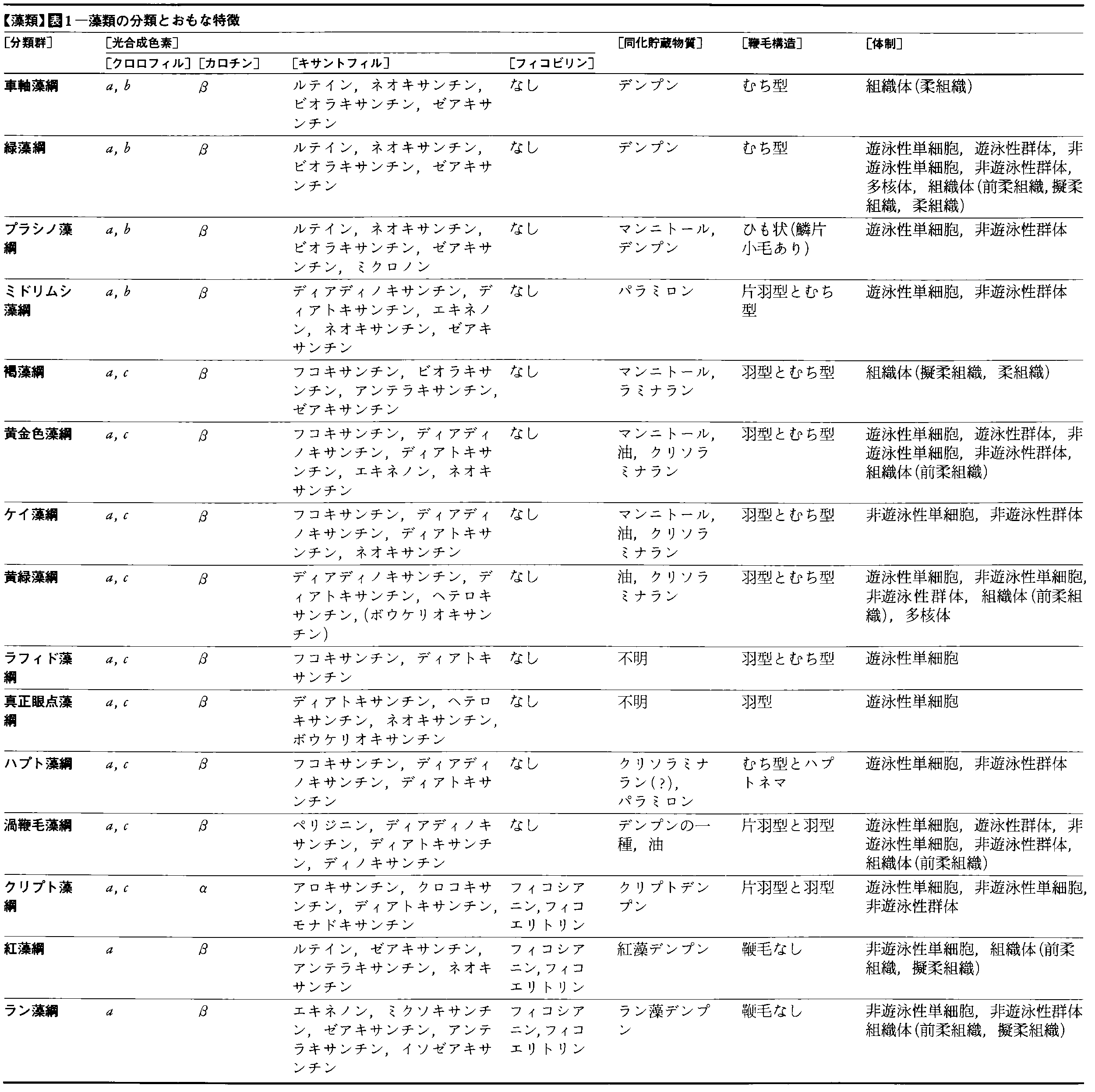
藻類の分類には,最も重要な物質代謝である光合成に関する特徴が,第1の基準となっている。この分類の歴史は古く19世紀の前半までさかのぼる。当時の海藻研究の第一人者であったアイルランドのハーベーW.H.Harveyは,海藻の体の色の違いが体の構造や生殖方法の違いと対応することを知り,色を基準にして海藻を(1)Rhodospermeae,(2)Melanospermeae,(3)Chlorospermeaeの3群に分類した(1836)。これらの(1)は現在の紅藻綱Rhodophyceae,(2)は褐藻綱Phaeophyceae,(3)は緑藻綱Chlorophyceaeにそれぞれ相当する。色は視覚で容易にとらえられる一種の形態的特徴であるせいか,当時この分類は人為分類であるという批評もあった。しかし,20世紀に入ると,色の違いは色素体に含まれる光合成色素組成の違いに起因することや,色が違うと光合成産物も異なることなどがわかり,ハーベーの分類は優れた自然分類としての地位を固めるに至った。さらに,光合成色素組成や光合成産物の基本的な違いは,生物の重要な営みである生殖に関与する細胞の構造の違いともよく対応することが明らかとなってきた。このことを少し詳しく説明すると次のようである。クロロフィルaのほかに主要な補助色素としてフィコエリトリンやフィコシアニンなどの色素タンパク質をもち,光合成産物として紅藻デンプンを貯蔵する紅藻類は鞭毛をもたない生殖細胞をつくるのに対し,クロロフィルaとcのほかにフコキサンチンをもち,ラミナランやマンニトールを貯蔵する褐藻類は,長短2本の鞭毛を側部にもつ先のとがった卵形の泳ぐ生殖細胞を形成する。この2本の鞭毛のうち,前方に伸びるものは両側に小毛を並列する,いわゆる羽型鞭毛であり,後方に伸びるものは表面に付属物のないむち型鞭毛である。またクロロフィルaとbをもち,デンプンを貯蔵する緑藻類では,泳ぐ生殖細胞は先のとがった卵形が基本型で,前端から前方に等長,むち型の鞭毛が伸びる。現在,藻類の分類体系は研究者により多少扱い方が異なるが,光合成と生殖にかかわる特徴を主要な基準にとりあげ,次の15綱に分類する場合が多い(表1参照)。ラン藻綱Cyanophyceae,紅藻綱Rhodophyceae,クリプト藻綱Cryptophyceae(=褐色鞭毛藻綱),渦鞭毛藻綱Dinophyceae(=炎藻綱,橙藻綱),ハプト藻綱Haptophyceae,褐藻綱Phaeophyceae,黄金色藻綱Chrysophyceae(=ヒカリモ綱,黄色鞭毛藻綱),ケイ藻綱Bacillariophyceae,黄緑藻綱Xanthophyceae(=不等毛綱),ラフィド藻綱Raphidophyceae(=緑色鞭毛藻綱),真正眼点藻綱Eustigmatophyceae,ミドリムシ藻綱Euglenophyceae,プラシノ藻綱Prasinophyceae(=異型緑藻綱),緑藻綱Chlorophyceae,車軸藻綱Charophyceae。
陸上のコケ植物,シダ植物,種子植物は,上記の光合成や生殖細胞の特徴が基本的には車軸藻類や緑藻類と同じである。このことからコケ植物,シダ植物,および種子植物などは,車軸藻綱や緑藻綱などとともに緑色植物の1亜門として扱われることがある。これらの植物はすべて系統的には同じ系列に所属するもので,区別点は,生殖器官が単細胞であるか多細胞であるか,受精卵が親の体内で発生を始めるか,それとも体外でするか,すなわち,胚をもつかもたないか,植物体に維管束があるかないか,そして繁殖の手段として種子をつくるか,つくらないかなどである(表2参照)。
最近,電子顕微鏡による研究の結果,緑藻類には,細胞分裂の際の隔膜形成の様式が大別して三つあり,しかもそれらの違いは生殖細胞の鞭毛基部装置の構造の違いともよく対応することが明らかとなり,これらの事実に基づいて,緑藻綱をコレオケーテ綱Coleochaetophyceae,緑藻綱Chlorophyceaeおよびアオサ綱Ulvophyceaeの3綱に分ける分類系が提案されている。このうち,コレオケーテ綱は陸上植物につながる系統群であるのに対し,狭義の緑藻綱はもっぱら淡水に,そしてアオサ綱は海にそれぞれ生育の場をもつ系統群である。
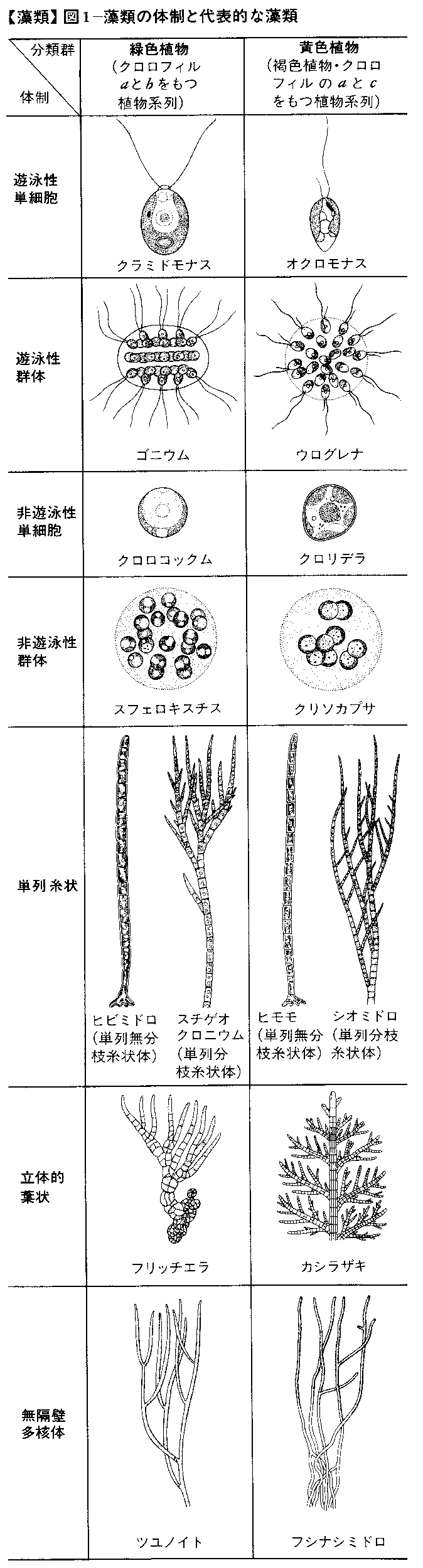
藻類は陸上植物に比べて体制ははるかに単純であるが,多様性に富んでいる。次におもな体制と代表的な藻類名を記す(図1)。
(1)遊泳性単細胞体制 クラミドモナスChlamydomonas(緑藻),オクロモナスOchromonas(黄金色藻)。(2)遊泳性群体体制 ゴニウムGonium(緑藻),ウログレナUroglena(黄金色藻)。(3)非遊泳性単細胞体制 クロロコックムChlorococcum(緑藻),クロリデラChloridella(黄緑藻)。(4)非遊泳性群体体制 スフェロキスチスSphaerocystis(緑藻),クリソカプサChrysocapsa(黄金色藻)。
次に記す(5)~(7)は組織をもつ体制で,ここには細胞が1平面だけに分裂してできる単列糸状体制,2平面に分裂してできる平面的葉状体制,さらに3平面に分裂が起こる立体的葉状体制がある。(5)単列糸状体制 ヒビミドロUlothrix,スチゲオクロニウムStigeoclonium(緑藻),シオミドロEctocarpus(褐藻),ヒモモTribonema(黄緑藻)。(6)平面的葉状体制 ヒトエグサMonostroma(緑藻),アマノリPorphyra(紅藻)。(7)立体的葉状体制 フリッチエラFritschiella(緑藻),カシラザキHalopteris,コンブLaminaria(褐藻),ヒジキHizikia(褐藻),ホンダワラSargassum(褐藻)。
以上のほかに特殊な体制として多核体制があり,これには体が単一細胞からなるもの,隔壁をもつ細胞からなるもの,および変形ともいうべき無隔壁単核体制がある。(8)無隔壁多核体制 ミルCodium(緑藻),ツユノイトDerbesia(緑藻),フウセンモBotrydium(黄緑藻),フシナシミドロVaucheria(黄緑藻)。(9)有隔壁多核体制 シオグサCladophora(緑藻),ジュズモChaetomorpha(緑藻)。(10)無隔壁単核体制 カサノリAcetabularia(緑藻)。
これらの体制のうち(1)は最も単純なもので,おそらく生物の始原的な形態に近いものであろうと考えられている。最も複雑な形態は(6)に見られ,褐藻ホンダワラ属の体は藻類の中で最高度に分化して,根,茎,葉の区別が外見上明りょうであり,陸上植物の体を思わせる。
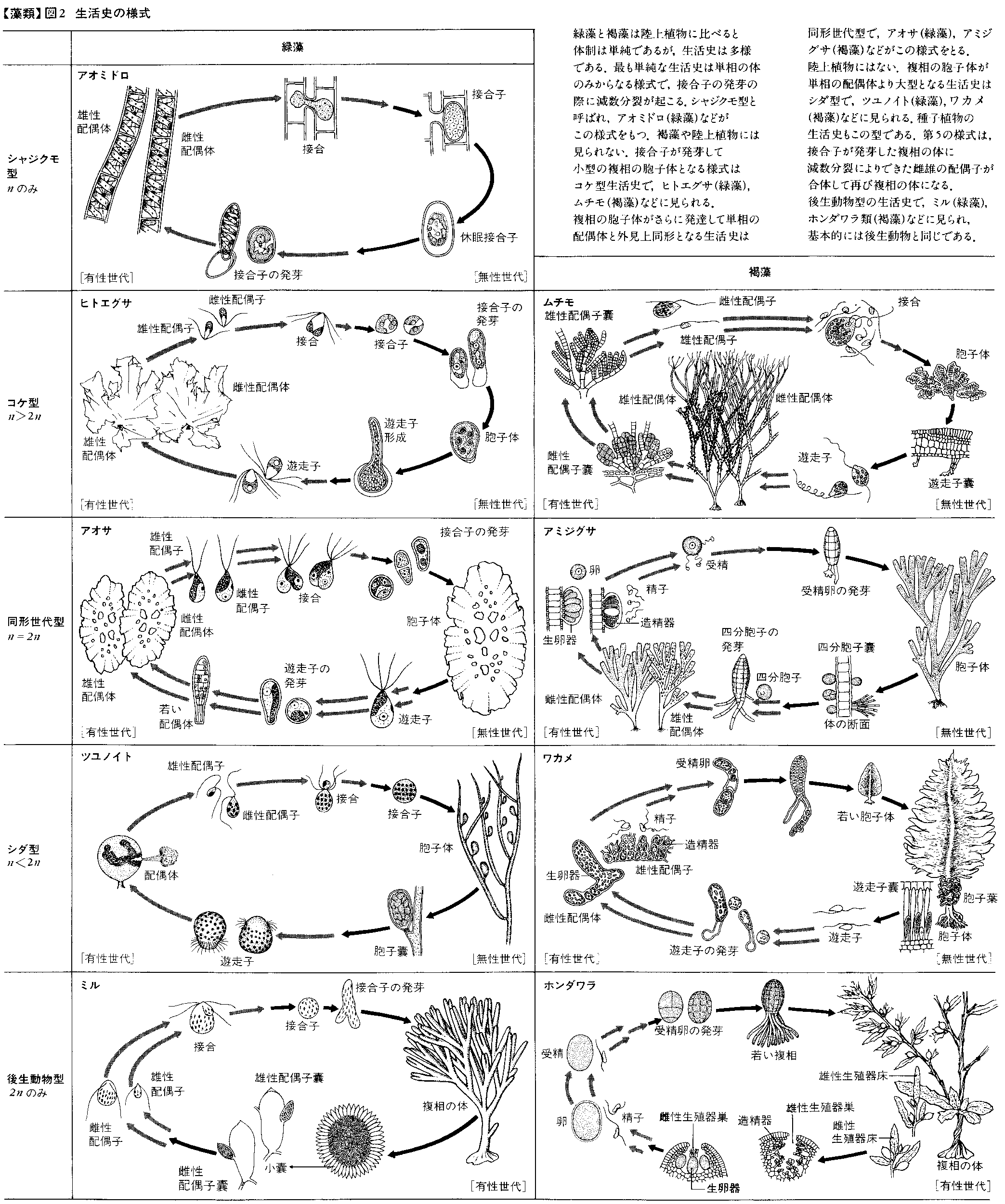
藻類の世代交代には大別して三つの様式がある。第1は外見上同形の配偶体と胞子体の間で世代交代をする。いわゆる同型世代交代で,褐藻のアミジグサ,緑藻のアオサやシオグサなどに見られる。第2は大型の配偶体と小型の胞子体の間の世代交代で,褐藻のムチモやカヤモノリ,紅藻のアサクサノリ,および緑藻のヒトエグサなどに見られる。この様式は基本的にはコケ植物の世代交代と同じである。第3は,逆に小型の配偶体と大型の胞子体の間の世代交代で,褐藻のコンブやワカメ,緑藻のツユノイトなどに見られる。この様式は基本的にはシダ植物の世代交代と同じである。上記のいずれの世代交代の場合も,配偶体は卵と精子あるいは同形または異形の配偶子を形成し,これらの合体によってできた接合子が発芽すると胞子体となる。胞子体は後に成熟して減数分裂を行って遊走子や四分胞子をつくり,これらが発芽して配偶体にもどる。したがって,配偶体の核相は単相(n),胞子体は複相(2n)である。すなわち,世代交代に伴って核相交代も行われていることがわかる。
藻類には世代交代をしない生活史をもつものもある。そのような生活史には大別して二つの様式がある。一つは緑藻のアオミドロや車軸藻のシャジクモとフラスコモに見られるもので,ふつうに見る藻体は単相の配偶体で,ここにつくられた雌雄配偶子の合体により生じた接合子は,減数分裂を行って配偶体の始原細胞をつくる。したがって,これらの藻類の生活史は単相の配偶体の世代だけからなり,複相は接合子の時期のみである。この生活史とは逆に,褐藻のホンダワラや緑藻のミルの生活史は複相の世代のみからなる。この藻体で減数分裂によりつくられた卵や精子,あるいは異形の雌雄配偶子が合体してできた接合子が発芽すると,親と同形の体に生育する。この生活史の様式は,基本的には後生動物のそれと同じである。
以上に述べた生活史の様式をまとめると,次のようになる。(1)単相の体のみ,(2)単相の体>複相の体,(3)単相の体=複相の体,(4)単相の体<複相の体,(5)複相の体のみ(図2)。
紅藻のテングサやオゴノリの世代交代は,一見,上述の同型世代交代に似るが,実際にはさらに複雑である。配偶体にできた卵と精子の合体により生じた接合子は,母体内で発芽して果胞子体と呼ぶ微細な体の世代となる。後にここに果胞子が形成され,これが体外に放出されて発芽すると,外見上配偶体と同形の四分胞子体に発達する。四分胞子体には減数分裂によって四分胞子がつくられ,発芽すると,そのうちの2個は雌性の,他の2個は雄性の配偶体にそれぞれ発達する。すなわち,テングサやオゴノリでは,配偶体,配偶体に内生する微細な果胞子体,および配偶体と同形の四分胞子体の順序で,三つの世代が循環する。紅藻のカギノリやフサノリも三世代植物であるが,四分胞子体が極端に小さい生活史をもつことで特徴づけられる。
化石の記録によると,藻類は約30億年前の先カンブリア時代にはすでに地球上に出現していたらしい。この時代の地層から発見された直径約20μmの球形のラン藻様の微化石Archaeosphaeroides larbertonensisが最古の藻類化石と考えられている。年代が下り約19億年前の地層からは現生のユレモなどに似たラン藻の化石が多数得られている。真核性の藻類の出現はそれよりかなり遅れたらしい。約9億年前の地層からは核やピレノイドをもつ細胞分裂中の緑藻と思われる化石が得られ,Glenobotrydionと名づけられている。紅藻のサンゴモ類,緑藻のカサノリ類,および車軸藻類は炭酸カルシウム(石灰)を,またケイ藻類はケイ酸をそれぞれ体に沈着するので化石として残りやすい。化石の記録から,サンゴモ類やカサノリ類は4億~5億年前のオルドビス紀に,車軸藻類は約4億年前のシルル紀にすでに生育していたことがわかる。興味あることに,ケイ藻類は単細胞性であるが,出現は遅く,白亜紀以降である。ケイ藻の化石はケイ藻土として多量に出現するが,これが見られるのはさらに年代の遅い第三紀と第四紀の新生代である。すなわちケイ藻の出現は今より約1億年前であり,3億~4億年前に出現した陸上植物よりケイ藻類ははるかに後出の生物であるといえる。また本来,南方系である緑藻類は北極にも生育し,緑藻類のうち10%が海水に,残り90%が淡水にみられる。この多様な生育場所をもつ緑藻類から,古生代オルドビス紀からシルル紀のころに,陸上植物が出現したといわれる。
藻類は有機物の生産者として水界の生態系において重要な役割を占めている。地球上の全植物の年間純一次生産量は170×109t(乾量)で,そのうち水界の植物の生産量は約1/3の55.5×109tであるという(ホイッタカーR.H.Whittaker,1975)。水界には種子植物も生育することを考慮に入れても,藻類が水界で果たす生態的役割がいかに大きいかがうかがい知れよう。なお,海藻の生産量はプランクトン性藻類のそれの約1/10であるという(ライザーJ.H.Ryther,1963)。しかし,海藻は沿岸帯に海中林や海中草原ともいうべき群落をつくり,沿岸性の魚貝類の生息場としてまた餌料植物として水産上重要な役割を果たしている。飼育実験によると,アワビが肉1kgを増量するのに褐藻カジメを約15kg摂取するという。プランクトン性の藻類や定着性の微細な藻類は稚魚貝や動物プランクトンなどの餌料として食物連鎖のうえで重要な位置を占める。ところで,ラン藻,ケイ藻,渦鞭毛藻などのプランクトン性藻類は環境が変化すると突然異常に大発生して,海では赤潮を,湖や池では水の華と呼ぶ現象を起こすことがある。赤潮や水の華は,水界の物理化学的性質をさらに急変させ,魚貝類を死滅させるなど,しばしば大きな被害をもたらす。
(1)食用 藻類はタンパク質と無機質を比較的多く含み,またビタミン類や脂質も含むので,健康保持の食品としての価値は高い。しかし,乾燥重量の約半分を占める多糖類の消化吸収率が,陸上植物に比べるとはるかに低いので,カロリー価は低い。藻類が整腸作用に効果があり,また美容食として優れた食品であるのはこのためである。最近,欧米でも海藻をsea vegetable(海の野菜)と呼ぶなど,健康食品として見なおす傾向にある。食用とするおもな藻類に紅藻のアサクサノリ,褐藻のモズク,コンブ,ワカメ,ヒジキ,緑藻のヒトエグサ,アオノリなどがある。
(2)寒天,アルギン酸,カラギーナン 紅藻のテングサ類,オゴノリ類からは寒天,褐藻のカジメ,アラメなどのコンブ科植物からはアルギン酸,紅藻ツノマタやスギノリ類からはカラギーナンがとれる。寒天はようかん,ゼリーなどの原料,ジャム,マーマレードなどの安定剤に使うほか,細菌の固形培養基の基質に使われる。アルギン酸はアイスクリーム,ゼリー,チョコレートミルクなど,多くの食品の安定剤に使われ,また薬品,乳化剤,洗剤,化粧品などの基剤にも使われ,用途が広い。このほかに織物用の経糸(たていと)のりや捺染のりにも多く使われる。欧米で生産の盛んなカラギーナンもアルギン酸と同じように,食品などの安定剤に使われるなど,広い用途をもっている。
(3)タンパク質資源 淡水藻の緑藻クロレラとラン藻スピルリナはタンパク質の含有量が多く,また生産性も高いので,最近では大量に培養され,粉末食品が作られている。
(4)駆虫剤 カイニン酸を含む紅藻マクリと,ドウモイ酸を含む紅藻ハナヤナギは優れた駆虫の効能をもつ。
(5)ケイ藻土 ケイ藻の化石の堆積物であるケイ藻土はろ過器のろ材,研磨材,断熱煉瓦,絶縁材料および緩下剤などに使われる。
(6)肥料,飼料 海藻は田畑の肥料として用いられたが,最近は少ない。ネンジュモやアナベナなど,ラン藻には空中窒素固定能をもつものがある。そこで,これらのラン藻を大量培養して水田の肥沃化を図る事業が,インドや中国で行われている。ヨーロッパでは古くから家畜の飼料として海藻を利用してきた。用いられる海藻はおもに褐藻コンブ科とヒバマタ科のもので,これらを粉末とし一般飼料に混ぜて与える。この結果,鶏卵や牛乳のヨード含有量の増加が認められている。
(7)エネルギー源 新しい試みとして,オオウキモを発酵させメタンガスを採り,これをエネルギー源にしようという研究がアメリカで行われている。
執筆者:千原 光雄
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
algae
隠花植物(葉状植物)に属し,緑藻・褐藻・紅藻・ラン藻・珪藻,さらに広範な分類範囲を含んでいる。形態も単細胞の微小なものから数mに及ぶ大型のものまで多様である。化石としては,これらのなかで石灰質や珪質の硬組織を有するものが残りやすく,石灰藻および珪藻とまとめて呼ばれ,多くの研究がなされている。しかし褐藻類のような軟組織の化石も若干報告されている。日本では中新統から褐藻類カイフモク(Cystophyllum sp.)の化石が報告されている。コダイアマモは藻類でなく水生の顕花植物の化石である。
執筆者:堀口 萬吉
出典 平凡社「最新 地学事典」最新 地学事典について 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
Sponserd by ![]()
…アルプスなど山岳地帯の人々は,春の山野に萌える山菜を好んで利用している。 藻類は,アサクサノリのように栽培的に生産されるもののほかは,利用はすべて採集による。緑藻類(ヒトエグサ,スジアオノリ,カワノリ,クロレラなど),褐藻類(モズク,マツモ,マコンブ,ミツイシコンブ,トロロコンブ,ワカメ,ヒジキなど),紅藻類(アサクサノリ,スサビノリ,チシマクロノリ(岩海苔),フクロフノリ,マフノリなど),ラン藻類(スイゼンジノリ,カワタケなど)がある。…
…土壌中に存在する微生物で,細菌,放線菌,糸状菌,藻類,原生動物などをいう。肥沃な表土には,土壌1g当り細菌数が数十億,糸状菌の菌糸長が数百m,微生物の生体重が土壌有機物量の数%に達することがある。…
※「藻類」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
夏の暑さに体が慣れること。数日から数十日間で起こる短期暑熱順化と、数年または数世代にかけて起こる長期暑熱順化とがある。→寒冷順化[補説]近年では、冷房設備の普及にともない短期暑熱順化が起こりにくくなっ...