デジタル大辞泉 「人種」の意味・読み・例文・類語
じん‐しゅ【人種】
2 人をその社会的地位・生活習慣・職業や気質などによって分類していう言い方。「仕事を生きがいとする
[類語]民族・種族
ひと‐だね【人種】
「声をかけ合って
2 精液のこと。
「おめえさんを女にすると、ほんに―が尽きいす」〈洒・遊冶郎〉
Sponserd by ![]()
翻訳|race
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
人種は,進化と並んで,ヒトの生物学的多様性に関する主要な概念である。しかし,しばしば意図的あるいは無意識的に,政治的迫害や社会的差別を合理化するために利用された歴史があり,それは現代においても,かつてのナチスによるユダヤ人の迫害や南アフリカ共和国のアパルトヘイトほどではないにしても,依然として続いている。そのため,人種という語の使用自体が問題視されることも多く,研究者の姿勢も分かれている。人種という概念を学問的に正しく定義し,差別が不合理であることを示すべきであるという意見がある一方で,人種という概念の科学的正当性を示すことは,結果的に人種差別の擁護に利用されることになるので,人種という語の使用をやめるべきだという意見もある。
人種という言葉を使用するかどうかは別として,人々が自分や他者の出自集団を認識する際に,言語その他の文化とともに,身体的特徴を手がかりとすることは否定しがたい現実である。出自集団の認識は,配偶者選択や生存戦略に大きく影響し,その結果,人類の集団遺伝学的構造を決める重要な要因になっている。つまり,人類は,自らの生物としてのあり方を,人種という概念を使って構造化している。人種による差別や迫害が生じるのは,配偶者選択や生存戦略のあり方に問題があるのであり,出自集団を身体的特徴で認識することや,構造化に出自集団という概念を使うこと自体に問題があるのではない。また,病気の罹患率や罹患した場合に耐性をもつ者の割合は,人種によって異なることが少なくない。したがって,保健医療において,人種の概念は一定の有用性をもっている。
ここでは,人種という語の使用が上述のような問題を含んでいることを念頭に置きつつ,人間の世界観および人類学における人種概念の歴史と意義について解説を行う。
ヒトという生物は,自身をとりまく世界を分類し命名することによって認識し,その認識を他者と共有する。人類の生物学的多様性に対する認識もこの方法で行われてきており,人種の概念はその延長線上にある。その最も古い例を,紀元前12世紀のエジプトの王墓の壁画に描かれた人物に見ることができる。エジプト人,セム人,黒人,リビア人が皮膚の色,髪の色と縮れ具合,眼の色,鼻の形,唇の厚さを明確に変えて表されており,当時のエジプト人がそれらの民族の違いを身体的特徴によって認識していたことがわかる。
人類一般からみると,上述のエジプト人の例のように,生物学的特徴だけによって異民族を分類することは例外的で,多くの場合,文化的特徴と生物学的特徴の両方が区別されずに用いられてきた。ヨーロッパにおいては,ギリシア時代からローマ時代にかけて,帝国の版図の拡大とともに,身体的特徴の多様性よりも言語や価値観の共通性が重視され,その傾向は,すべての人類はアダムとイブの子孫であると説くキリスト教によって強化され,中世まで続いた。
ヨーロッパ人が人類集団の多様性に興味を抱くようになったのは,15世紀に始まった大航海時代である。新大陸からヨーロッパにもたらされた物や知識は,聖書の記述には収まりきらず,人々の認識の変更を迫り,地理学や分類学,さらに諸科学の発達につながった。異民族の認識に関しては,アダムとイブの子孫が住みついたとは聖書に書かれていないアメリカ大陸に,先住民がいたことが議論を引き起こした。アメリカ大陸先住民は,聖書の記述に従えばアダムとイブの子孫ではなく,したがって人間ではないとするスペインの征服者によって,搾取や奴隷化が行われた。その有様を見かねたドミニコ会修道士の訴えにより,1537年ローマ教皇パウロ3世は,アメリカ先住民も真正な人間であるという勅書Sublimus Dei(〈神の気高さ〉の意)を発し,アメリカ先住民の自由と財産に対する権利を認め,奴隷化を禁じたが,それは1521年のコルテスによるアステカ帝国の征服や1533年のピサロによるインカ帝国の征服より後のことであった。また,この議論は,何をもって人間とみなすかに関するもので,文化的多様性や生物学的多様性に注目したものではなかった。
人類の生物学的多様性に関する科学的な考察のうち,ヨーロッパの中世以降で最初のものは,医師であり旅行家であったフランス人のフランソワ・ベルニエFrançois Bernierが1684年に出版した《人種による新しい世界の区分》に見られる。ベルニエは,類似した身体的特徴をもつグループを〈人種race〉と呼び,肌の色以外の点でヨーロッパ人に似た人々が北アフリカからインドまで分布することを指摘した。
リンネ(1707-78)は,生物の多様性を対象とする学問である分類学を集大成し,1758年に出版した《自然の体系Systema Naturae》第10版において,ヒトHomoを霊長目Order Primatesの四つの属の一つに位置付けた。ちなみに他の3属は,Simia(サル),Lemur(キツネザル),Vespertilio(コウモリ)であった。リンネが人間に与えたHomo sapiens(賢いヒト)という名称には,精神性を重視する文化的伝統の影響を認めることができる。彼は,Homo sapiensという種をさらにAfricanus niger(アフリカ人),Asiaticus fuscus(アジア人),Europaeus albescens(ヨーロッパ人),Americanus rubescens(アメリカ大陸先住民)に分類した。中世のキリスト教徒の間では,ノアの3人の息子であるハム,セム,ヤペテが,それぞれアフリカ人,アジア人,ヨーロッパ人の祖先であると信じられており,リンネの人種分類は,これら三つにパウロ3世の勅書Sublimus Deiで追加されたアメリカ先住民を加えたものになっている。また,これら以外にMonstrosus(化け物)といった分類まで設定している。このことからわかるように,リンネにおいては,ヒトや人種に学名を与え,自然界の一部に位置付けることにより形式上は科学の対象として取り扱われているが,内容はキリスト教の伝統や言い伝えに沿ったものであった。
具体的な身体的特徴に基づいた人種区分を初めて行ったのは,ドイツのブルーメンバハ(1752-1840)である。彼は,1775年の論文〈ヒトの自然的変種〉において,主に皮膚の色と頭型に基づいて,人類をコーカシア,モンゴリア,エチオピア,アメリカーナ,マレーの五つに分類した。コーカソイドやモンゴロイドといった名称はこれに由来する。ブルーメンバハは,これらの五つの自然的変種は連続的な変異であって,それらの間に明確な境界はないとする一方,コーカシアがその基本形で最良のものであるとした。
この例でもわかるように,近代における人種概念は,具体的なデータに基づいた実証的な概念になる一方,差別や支配を正当化する傾向が科学的な装いをこらして混入した。人類は古代から集団間の交渉を繰り返してきており,それが敵対的な関係や支配・被支配の関係である場合,集団の弁別的特徴は,身体的であれ文化的であれ,蔑視や崇拝の対象とされることが多い。19世紀から20世紀にかけての植民地をめぐる争いや支配・被支配の関係においては,人種概念がこの役割を果たした。白人が他の人種より能力,道徳性とも優れているというゴビノー(1816-82)らの《人種不平等論》は,この時代,軍事的,政治的,経済的に優勢であったヨーロッパ人が他の民族を支配するための根拠として利用された。それはナチスドイツでは優生学と結びついてアーリヤ人種礼賛やユダヤ人迫害へとつながり,また,アメリカ合衆国では奴隷制や人種差別を正当化する役割を果たした。アメリカ合衆国における人種概念は,国民を白人whiteと非白人coloredに区分し,祖先がすべて白人であるものだけを白人,それ以外をすべて非白人とするもので,出自に関する社会的な制度であり,生物学的な装いは一種の虚構といえる。
差別に利用された歴史に対する反省から,現在では,人種という言葉やコーカソイド,モンゴロイド,ネグロイド(ニグロイド)などの名称が科学的な研究や論考の中で使われることは少なくなってきた。ヒトの種内の生物学的多様性の研究が行われなくなった訳ではないので,その類別的記述としての人種の概念自体は存続しており,コーカソイド,ネグロイド,モンゴロイドにほぼ対応するものとして,ヨーロッパ人,アフリカ人,アジア人という地理的大集団名が使われることが多くなっている。
地理的大集団間の変異は,肌の色を除けば,集団内の変異に比べてはるかに小さく,ある個人がどの大集団に所属するかを,その遺伝的な身体的特徴から判定することは困難である。また,肌の色も大集団の区分とは一致しない。すなわち,身体的特徴から個人の所属集団は特定できない。現代の人種概念と古典的な人種概念は,この点で大きく異なっている。
生物の多様性には進化が反映されている。したがって,人種概念は人類進化に関する考え方の影響を強く受けている。現生人類の進化に関する学説は遺伝学の発展により1990年前後に大きく変化した。1980年代まで主流であった考え方は,主に化石の証拠に基づいて,人類は限られた遺伝的交流を保ちながら,各地域で原人から旧人,さらに新人すなわち現生人類へと進化したとするものであった。ヨーロッパ人はネアンデルタール人の直系の子孫であるとされ,人種間の差異は長期にわたる地理的隔離や環境への適応の結果生じたと考えられた(多地域進化説)。これが正しければ,人種は100万年以上の長い歴史をもつことになる。しかし,1990年代以降,ミトコンドリアDNAの分析から,現生人類は約20万年前のアフリカに共通祖先をもち,8万年から5万年前にその一部がアフリカを出て,アジアやヨーロッパに広がったとする説(出アフリカ説)が主流になってきた。この考え方によれば,地理的大集団への分化の歴史は浅く,集団差の生物学的な意味は限られたものになる。現在も二つの考え方の間の論争は続いているが,遺伝学的データは出アフリカ説を支持するものが多い。
人類の遺伝的多様性の大部分はアフリカ人の中の多様性であり,アジア人とヨーロッパ人は,アフリカ人の多様性の中に埋没し,地理的大集団間の差異は人類全体の多様性の10%程度にすぎない。したがって,人種すなわち地理的大集団は,人類の遺伝的多様性全体のニュートラルな表現ではない。おもに肌の色や鼻の形などに関する特徴に基づいており,結果として,気候への適応に関係した区分になっている。つまり,人種特徴は,アフリカを出た人類の旅の歴史を反映しており,人類がアフリカを出てから高緯度地方に移住していく間に経験した紫外線の強さや気温の低下による淘汰圧の変化に対応したものになっている。これは,人類の移動の歴史において,大部分の移動が徐々に行われ,その間に文化だけでなく身体的な変化による適応が生じたためである。しかし,大航海時代以降の移動は,非常に短期間に行われたので,旅の経験はほとんど身体には刻印されていない。つぎに,大航海時代以前の地理的大集団の分布と特徴を概説する。
アフリカ人はホモ・サピエンス発祥の地であるアフリカにとどまった人々の子孫であり,熱帯地方で進化した人類の一般的な特徴をもっている。すなわち,強い紫外線を遮るために皮膚は濃色で,高い気温に適応して鼻幅が大きい。また,遺伝子プールが大きいことを反映して身長的特徴は変化に富み,乾燥地帯に住む高身長のマサイ族や深い密林の生活に適応した低身長のピグミーなど,広い変異の幅がみられる。アフリカ人で鎌形赤血球遺伝子の頻度が高いのは,熱帯地方に多いマラリアに対してそのヘテロ接合型が耐性をもつからであると考えられている。なお,Y染色体上の塩基配列を用いた研究により,現在のアフリカ人の一部には,いったんアジアに向かった人々の一部がアフリカへ戻ったことによると考えられる遺伝的影響が認められることが明らかにされている。
サハラ砂漠以南のアフリカでは,約3000年前以降,農耕民・牧畜民の移動により,集団の分布が大きく変化したと考えられている。現在では農耕民が大部分の地域に広がり,狩猟採集民は砂漠や密林に残存するだけであるが,それ以前はまったく様相が異なっていた。東アフリカで出現した現生人類は,やがてサハラ以南全体に広がり,地域集団間の遺伝的な差を広げながらも,狩猟採集生活への適応からくる文化や身体的特徴の共通性を維持していた。湿地や密林で狩猟採集生活をしてきたピグミーや,砂漠に住む狩猟採集民のハッザやサン(ブッシュマン),サンから派生したと推定される遊牧民のコイ・コイン(ホッテントット)は,そのような狩猟採集民の残存であると考えられる。これらの集団は,遺伝学的には相互に非常に遠く隔たっているが,他のアフリカ人より比較的淡い黄褐色の皮膚や,毬状の頭髪,低身長傾向といった身体的特徴を共有している。このことは,これらの集団の共通の祖先が非常に古く,アジア人やヨーロッパ人が分かれる前の人類共通の祖先に近いことを示している。
アジア人は,ユーラシア大陸を東に旅した人々の子孫であり,熱帯から寒帯まで広い範囲に分布し,オーストラリア大陸,南北アメリカ大陸,太平洋の島々の先住民はすべてアジア人である。アフリカからアジアへの移動の回数や経路は,複数あった可能性がある。北アメリカへの移動は氷河期のベーリング海峡経由で,複数回あったと考えられている。アジア人の身体的特徴は多様である。多くの場合,熱帯地方に分布するアジア人は皮膚が濃色で鼻幅が大きく,比較的上下肢が長く,痩せ型であり,高緯度地方ではその逆の特徴を示す。しかし,いったん各地の気候に適応した後で起こった域内での移動の結果,気候と身体的特徴が対応していない場合がある。
遺伝学的にみると,アジア人の中では,東アジア人,アメリカ大陸先住民(アメリカ・インディアン),オーストラリア先住民(アボリジニーおよびトレス海峡諸島民)とニューギニア先住民からなるサフール人がそれぞれまとまりを作る。さらに,東アジア人の中では,内側眼瞼ヒダや比較的短い四肢などの寒冷適応を示す北方グループと,寒冷適応を示さない南方グループの間にやや違いがある。北方グループに多く見られる直毛の毛髪とシャベル状切歯とは,同じ遺伝子に支配されることが明らかにされている。また,ミクロネシア人やポリネシア人は,5000年前以降に移住した南方グループの農耕民の子孫である。大陸沿岸の島嶼部や半島に点在するネグリトは低身長や毬状毛,濃色の皮膚などの外見がアフリカのピグミーに似ているが,分子系統学的には明らかにアジア人で,サフール人に近く位置し,ユーラシア大陸の海岸に沿って広がった初期移住者の残存と考えられる。
ヨーロッパ人は,ユーラシア大陸の西部を北に向かって旅した人々の子孫である。遺伝学的な証拠により,その旅はアラビア半島からチグリス川・ユーフラテス川に沿ったルートをたどったと考えられている。中東からヨーロッパ全域に分布するとともに,東方にも移動し,南はインド亜大陸,北はシベリアまで達し,アフリカ北部にも進出した。皮膚と髪の色は連続的変化傾向(クライン)を示し,北に行くほど淡色である。皮膚色や毛髪の色は,同じ緯度に住むアジア人より一般に淡色である。また,虹彩の色は,アジア人は高緯度でも褐色であるのに対して,高緯度地域のヨーロッパ人では淡色のものが多い。これは,大陸の西部は東部に比べて曇りの日が多く,日照が少ないためと考えられる。ミトコンドリアDNAの証拠から,先住のネアンデルタール人との混血は生じなかったとされていたが,核DNAの研究により,ヨーロッパ人とアジア人の共通の祖先とネアンデルタール人との間に混血が生じ,1~4%の遺伝子を残していることが明らかにされた。なお,アフリカ人とヨーロッパ人の間の遺伝学的距離は,アフリカ人とアジア人の間の距離よりも小さい。これは,アフリカの周辺を離れた時期がアジア人よりも遅いことと,地理的にアフリカと近いため,一定の遺伝的交流があったことが影響していると考えられる。
混血も人種と同様,使用に注意が必要な言葉である。現生人類はホモ・サピエンス1種だけであり,どの集団も多かれ少なかれ他の集団と遺伝子の交換がある。したがって,人類の歴史において,純血の集団と呼べるものが存在したことは一度もなく,その存在を前提とした古典的な混血の概念は成立しない。
人類における混血は,地域集団間の遺伝子交換の一種である。ただし,混血と認識されるためには,二つの集団の身体的特徴に弁別可能な差があり,さらに混血の結果生まれた人々と元の集団の間にも弁別可能な差があることが期待されている。つまり,混血という概念は,生物学的であると同時に出自に関するものである。両親の集団が異なる場合,混血の結果生まれた子どもやその子孫は,アメリカ合衆国における〈有色人種〉のように,いずれかの集団に吸収される場合と,南アメリカにおけるメスティソのように混血者の集団を形成する場合とがある。
混血と認められる現象は,かなり大きい地理的集団間でのみ生じ得る。集団間の連続的な移行帯で持続的に生じる遺伝子の交換は,通常,混血には含めない。遠距離の交易に伴う小規模な混血は,北アフリカにおけるアフリカ人とヨーロッパ人との間や,東南アジアにおけるインド人とアジア人との間などで古くから生じていたが,大規模な混血は,主に大航海時代より後の大幅な分布域の変化によって出現した不連続的な境界で生じた。大航海時代以降,ヨーロッパ人が南北アメリカ大陸へ進出し,その際に労働力としてアフリカ人を移住させた。これによって,ヨーロッパ人,アフリカ人,アメリカ大陸先住民の間で混血が生じた。
南アメリカ大陸のうち,標高の高いアンデス地域では,ヨーロッパ人とアメリカ大陸先住民との混血(メスティソ)が多い。一方,ブラジルを中心とする東の低地では,ヨーロッパ人,アメリカ先住民,アフリカ人の間の通婚が進み,様々な程度の混血が見られる。ヨーロッパ人の割合とアフリカ人の割合は南北のクラインを示し,南の高緯度地域ではヨーロッパ人の割合が高く,北の熱帯地方ではアフリカ人の割合が高い傾向がみられる。
アメリカ合衆国では,ヨーロッパ人とアフリカ人との混血が進み,混血者はすべて〈有色人種〉として扱われた。一方,アメリカ大陸先住民とヨーロッパ人との混血では,どの程度の混血まで先住民とするかが問題になっている。いずれにしても,アメリカ合衆国における混血をめぐる問題は,支配権や所有権,先住民として保護を受ける権利などの継承に関連することから,出自の問題になっており,生物学的な概念ではとらえきれない。
オセアニアでは,18世紀以降,この地域に進出したヨーロッパ人とアジア人の子孫である先住民との間で混血が起こった。オーストラリアではアボリジニーとの混血が進み,ニュージーランドや南太平洋の島々では,ポリネシア人との混血が進んでいる。
南アフリカでは,17世紀以降オランダから,また18世紀末から19世紀にイギリスから入ったヨーロッパ人の移民とアフリカ人との間に混血が生じている。
執筆者:多賀谷 昭
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
現生の人類はホモ・サピエンスという単一種に属するが、多型的かつ多様的である。人種はその種内の変異の産物である。その変異は適応放散の結果であり、多分に地域と結び付く。人種は人類の生物学的・遺伝学的区分であるため、しばしば動物界の亜種に相当するとみられるが、それほど単純ではない。それは、人類には婚姻で代表されるような独自の繁殖制度があるからである。
人種は文化的・歴史的区分である民族、政治的・行政的区分である国民、宗教的区分である教徒などとは、別個の区分概念である。いかに巧みに英語を話し、アメリカ文化を身につけようとも、日系二世アメリカ人は身体的、人種的には日本人である。多くの人々はこれを十分心得ていながら、一方で、「人食い人種」などという言葉を口にしてきた。仮に、ある社会に人食いという慣習があったとしても、それは文化的行為であり、生物学的特性ではない。またユダヤ人種なるものを想定するが、それはおかしい。ユダヤ教に帰依(きえ)し、それゆえ独特の風俗習慣をもつ集団をユダヤ人とよぶことはできるが、それは人種ではない。金髪の多いポーランドのユダヤ人には金髪が多く、黒髪の多いスペインのユダヤ人には黒髪が多いということからでも、そのことは明白である。
本来人種は集団についていう。アメリカの人類遺伝学者スターンは「人種とは遺伝的に多少なりとも隔離された人類集団で、他のいかなる隔離集団とも違った集団遺伝子構造を有するもの」と定義した。したがって、人種は、統計学的思考によって理解されるものであり、一つの人種集団の成員はおおむね共通の人種特徴群をもつ。たとえば、金髪は北欧人のシンボルともいえる特徴であるが、その成員の4分の1は金髪ではない。しかし、その他の諸特徴によって北欧人集団に入れられる。また、ある種の血液型の頻度が二つの集団間で似ているからといって、ただちに両者を同一人種とみなすことはできない。他の血液型や人種特徴が異なる可能性があるからである。人種を考える場合、その集団の人口の大きさは重要である。人種をどの段階で区分するかは、多分に仮定的、恣意(しい)的である。なお、個人について某々人種であると一般にいわれるが、それはあくまで便宜的な称呼であり、厳密にいえば、某々人種型とよぶのがよい。
以上は従来の考え方であるが、最近、とくにアメリカ社会では人種という語が日本語におけるよりはるかに曖昧(あいまい)に用いられており、そのためその語の学問的使用が制限され、むしろ社会学的概念となっている。また人種差別問題との関係から、「人種」という語は自然人類学の教科書からは消え、そのかわりに「多様性」という語が登場している。多様性はまさしく人種概念の本質であるから、この意味で、本稿では使い慣れた「人種」という語をあえて用いることにする。これまでの日本の学界や教育界では、人種・民族・国民をきちんと使い分けてきたが、このほうがはるかに一般に理解されやすいであろう。
[香原志勢]
人種特徴には、遺伝すると考えられる形質があげられるが、その多くの遺伝様式は明らかでない。たとえば身長は人種により大いに異なり、また遺伝するものとされているが、一方、栄養や生活環境によって著しく影響される。その点、血液型や血清タンパクは遺伝様式がよくわかっているので、系統を論じるにはぐあいがよいが、とかく人類の認識は視覚に大きく頼るため、伝統的な人種概念は形態をもとにしており、人種特徴も目につきやすいものが多い。人種特徴として形態的なもののほか、生理的なもの、最近はとくに生化学的なものが広く採用されている。
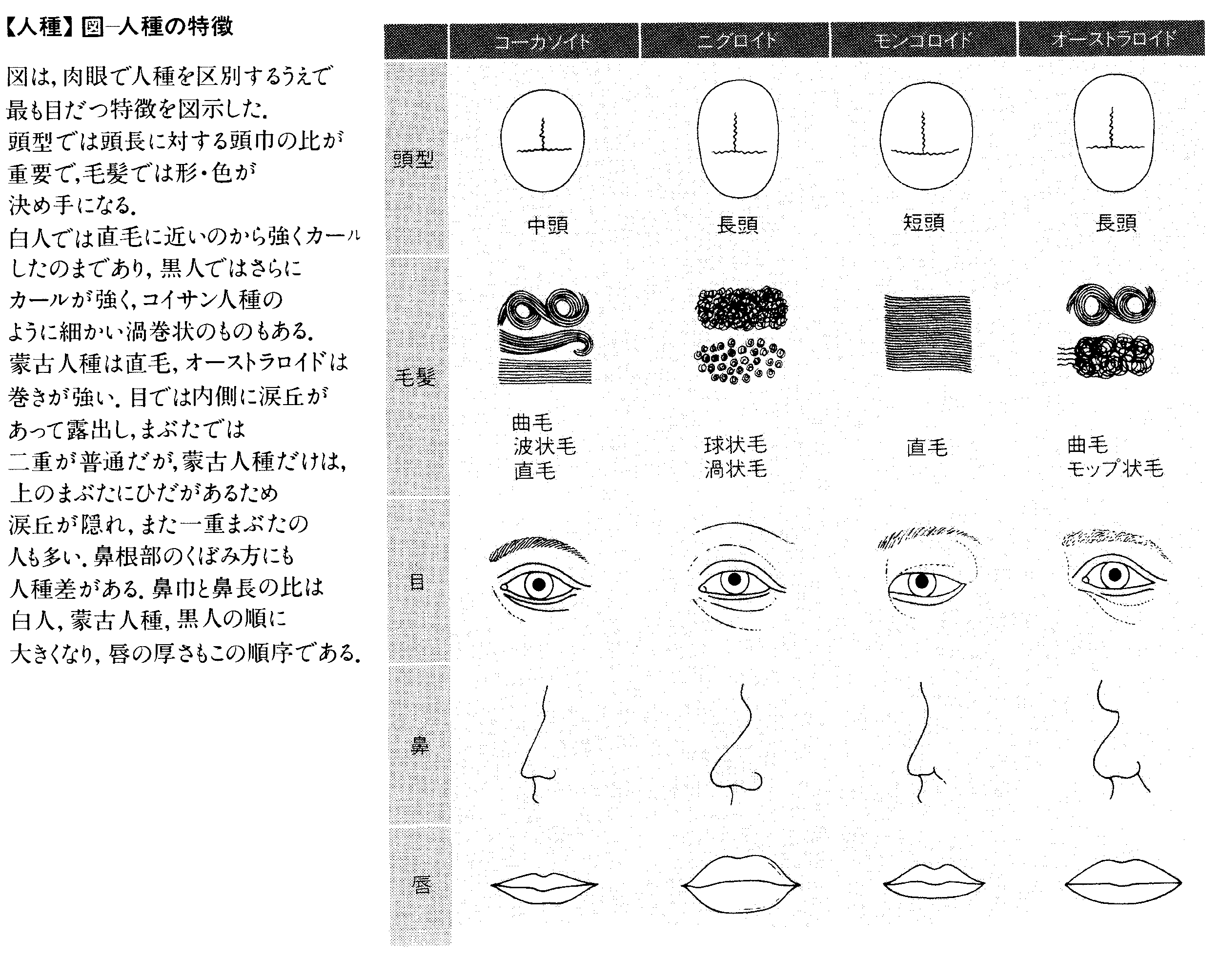
皮膚の色はもっとも目だつ特徴である。古くから用いられる白人や黒人などという俗称がその事実を裏書きしている。皮膚の色はメラニン色素の量によって決められており、それが多ければ黒色や濃褐色、中程度であれば褐色や淡褐色(いわゆる黄色)、ごく少なければ、毛細管の血液の色が透けて、赤みを帯びた白色になる。熱帯の人々は濃色の皮膚が多く、緯度が高くなるにしたがって明色の皮膚の個人の頻度が高くなる傾向がある。毛髪や目の虹彩(こうさい)の色も、メラニン色素の量の多少による。モンゴロイドの小児の尻(しり)の皮膚には青い斑紋(はんもん)がみられる。特別な機能はないまま、成人になると消失するので、小児斑という。コーカソイドではほとんどみられない。
頭髪の毛の形も直毛、波状毛、縮毛と、いろいろな段階がある。とくに著しい螺毛(らもう)は1本1本が独立に巻く。毛の横断面が円形ならば直毛であるが、楕円(だえん)の程度が増すにしたがって巻き方が強くなる。またコーカソイドの毛はモンゴロイドの毛に比べて細く、頭髪はふさふさとして柔らかくみえる。あごひげや体毛の量や生える部分は、人種によりかなりの差がある。若禿(わかは)げの出現度にも人種差があり、コーカソイド男子では出現度が高いが、アメリカ先住民(アメリカ・インディアン)ではきわめて低い。
頭の形も、頭最大長と幅、頭耳高、そしてこれらの値の間の示数、あるいは前・上・側面から見た頭の輪郭などについても、特定の人種に特有のものがある。顔や、鼻の幅や高さ、頬(ほお)骨の突出程度、頤(おとがい)の張り出し方、突顎(とつがく)=直顎の程度も重要な記載の対象であり、鼻根部付近の骨の形は人種によって特徴が出やすい。鼻や眼瞼(がんけん)(まぶた)の形、眼裂の方向、唇の厚さ、耳介や耳たぶの形など、頭蓋(とうがい)骨や顔面についても人種差がみられる。
身長、四肢(しし)や体幹の比率、体格の頑健(がんけん)度、姿勢、皮下脂肪、臀部(でんぶ)の形、乳房の形や大きさなどについても人種差がみられる。
以上のものは目につきやすいものであり、異人種意識をおこさせやすい形態学的特徴であるが、一方、目につきにくい特徴として、手足の指紋、掌紋の様式の頻度、歯の形や大きさ、骨の細部の形、筋肉や血管の変異など、多数のものをあげることができる。
コーカソイドやネグロイドは体臭が強く、モンゴロイドでは弱い。体臭はアポクリン汗腺(かんせん)の発達程度と関連する。わきががある場合、耳垢(みみあか)は湿性である。体力、基礎代謝量、耐寒性、耐暑性、耐光性も人種によりかなりの変異があるが、これらの機能は形態と多分に関連する。ネグロイドの優れた跳躍力はその細長い下腿(かたい)、突出したかかとなど、解剖学的見地からも十分予想できる。味覚障害や色覚障害の頻度も人種によって異なるが、これはそれぞれの人種集団の農耕経済、産業経済の採用の早い・遅いにかかわるところが大きい。一卵性双生児出産の頻度は全人種ほぼ同一であるが、二卵性は大幅に異なり、出生1000人につき、日本人は3人弱であるが、ネグロイドは20人である。身体発育の遅速にも人種差がある。知能や気質などにも人種差が予想されるが、それらの概念は明らかでなく、測定法も不完全なことが多く、また文化的要因の介入するところが甚だ大きく、現実には比較不能である。
生化学的な特徴には、遺伝様式が明快なものが多く、近年とみにその威力を発揮している。ABO式、MN式、Rh式など多くの血液型で集団内比率に人種差がみられる。とくにDNAによる検査は急速な発達をとげている。これらの諸特徴の総合化により、今日では各人種集団間の遺伝距離を相当程度数量的に表すことが可能となり、人種間の系統関係が明らかになりつつある。
[香原志勢]
古来、多数の学者がさまざまな人種分類表の作成を試みている。人種分類が非常に困難なのは、(1)古人類との関連が不明確であること、(2)人種特徴の評価が不定であること、(3)人種特徴、とくに目だつ特徴の遺伝関係が不明であること、(4)人種間の混血や移住が進み、近年では混住の傾向が広くみられ、集団の把握に苦慮すること、(5)一般動物と異なり、人類では人種形成の実験的研究が不可能であること、(6)人種というものの生物学的概念がかならずしも明確でなく、民族や国民と区別しにくい点があり、さらに研究者自身の世界観が投影されやすいこと、などがあげられる。とくに最近は混血、移住が激しく、重要な少数人種集団が絶滅に瀕(ひん)していることが、人種分類の作業のむずかしさに拍車をかけている。
今日よく知られている分類表としては、(1)毛髪を重んじたドゥニケ(1926)、(2)頭形、鼻示数を用いたディクソン(1923)、(3)毛髪に主点を置き、身長、頭示数、鼻示数を併用したハッドン(1930)、(4)三大人種を基幹とし、各種複合人種を想定して加えたフートン(1947)、(5)三大人種にそれぞれ主人種、副人種、特殊型、中間型を組み合わせた分類体系を作成したアイクシュテット(1939)、(6)三大人種に原始人種を加え、地理的分布に力点を置いたバロア(1948)、(7)人種形成論に主点を置くクーン(1950)、(8)遺伝様式の明確な形質を用いて分類に新分野を開いたボイド(1950)、(9)地理的人種、地域人種、微人種という概念を導入したガーン(1961)などがあげられる。
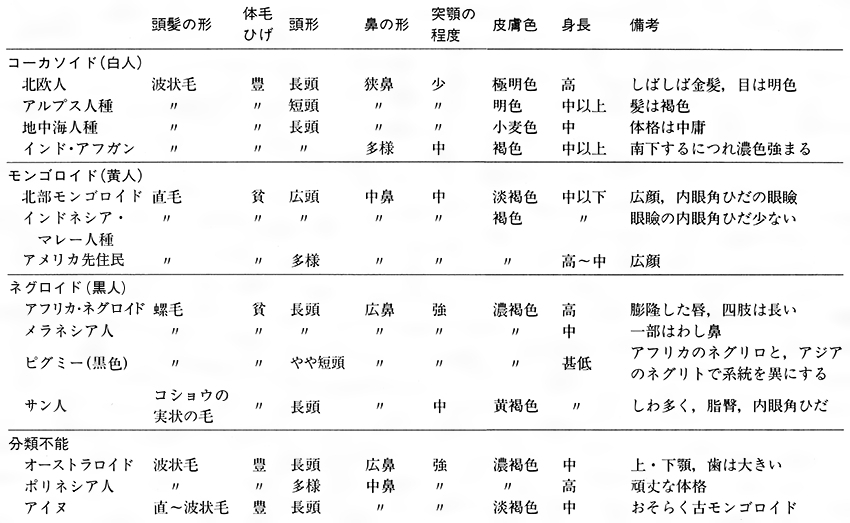
このような数多くの分類方法に共通するのは、ネグロイド、コーカソイド、モンゴロイド(語尾のオイドは「のようなもの」という意味)の三大人種である。なお、これらに対応して、体表の色彩に従った、黒人、白人、黄色人種という語があるが、俗称であり、適切なものではない。コーカソイドのなかには、白い肌の北欧人とともに黒い肌のインド・アフガン(いわゆるインド人)が入っている。両者の体表の差異は大きいが、骨格などはよく似ている。
ネグロイドの主体はアフリカ・ネグロイドであり、これに対してメラネシア人を大洋州ネグロイド、またはメラネソイドというが、最近の免疫化学的研究によれば、両者の間の系統関係はきわめて疎遠である。同様にコイサン人やピグミーもアフリカ・ネグロイドとはかなり疎遠である。
成人男子の平均身長が150センチメートルの集団をピグミー(ギリシア語で「こびと」の意)という。彼らはアフリカから東アジアに至る熱帯降雨林の奥深くに散在している。アフリカのそれをネグリロ、アジアのそれをネグリトとよぶが、両者の関係は浅い。短身は森林内での活動に適している。
三大人種に入れにくいものもあり、その一つはオーストラリア・アボリジニーである。これはインド南部やスリランカに住むベッダとともにオーストラロイドとよばれるが、比較的古代型を保っているとして独自な地位を与えられる。アメリカ先住民(アメリカ・インディアン)、ポリネシア人、ミクロネシア人をモンゴロイドから引き離し、三大人種という分類のなかに含めない見方もある。アイヌはこれまでコーカソイドの祖型とみなされたが、今日、日本の人類学者はこれをモンゴロイドの一つとみている。
[香原志勢]
人種の起源については多元論と単元論がしのぎを削ってきた。かつて、高名な人類学者のなかにも、「黒人と白人との差異は動物の種間の差よりも大きい」と唱える者がいたが、今日では、そのような考え方は完全に否定されている。
人類は精神活動が活発なため、異人種意識が強く、長い歴史の間では、似た者同士が集まって、目だつ形質を中心に一つの人種を形成したと考えられる。また、人類は種としてもっとも分布の広い動物であるため、さまざまな気候、地形に適応しなければならなかった。さらにその長い歴史は超過疎の状態にあったため、互いに隔離が進み、同時に遺伝的浮動を伴って、さまざまな人種に分かれたものと考えられている。
グローガーの法則によれば、温暖で多湿の地域の動物ほど、体表の色素沈着が著しいが、それは人間にもいえる。密林に住むネグリロは漆黒に近い。しかし、色素沈着は紫外線に対する反応である。紫外線の強い低緯度地帯の住民は濃色の皮膚をもつことで有害な紫外線の体内侵入を防ぐ。しかし、高緯度、とくに曇天の多い北ヨーロッパでは、白い肌のほうがビタミンD形成の補助をするに適した波長の紫外線を吸収しやすい。
ベルクマンの規則によると、寒冷地方の動物ほど体が大きい。また、アレンの規則によると、寒冷地方の動物ほど丸みを帯びた体をもつ。その点、エスキモーのずんぐりして、四肢の短く太い体格は酷寒の地によく適している。一方、ネグロイドやオーストラロイドの痩(や)せて、四肢が長く、すらりとした体格は、放熱作用を効果的にする。ネグロイドは手足の凍傷にかかりやすく、熱帯の日差しのもとに居住する皮膚の白い人々は皮膚癌(がん)にかかりやすい。なお、クーンその他は、モンゴロイドの顔や体は乾燥した酷寒の気候環境に適しているという。その顔は凹凸に乏しく、鼻は低く、頬骨は側方に高く張り、眼瞼は脂肪を豊かに含み、はれぼったくみえる。このような人種特徴をもつに至ったのは、その先祖が氷期の東北アジアの酷寒に適応して生き抜いたからだとし、その過程を十分に受けないでアメリカ大陸に渡ったのがアメリカ先住民(アメリカ・インディアン)である。
アフリカでは鎌(かま)状赤血球を遺伝形質としてもつ人々がいるが、正常状態では貧血しやすく、死亡率が高いにもかかわらず、その遺伝子の発現率は低下しなかった。それは、その形質保持者がマラリアに対する強い抵抗性をもっていたからである。その証拠として、非マラリア地域に移住した者の間では、この貧血症の出現率ははるかに低い。この貧血症はネグロイドに多く出現する。
[香原志勢]
諸人種の栄枯盛衰はそれぞれの人口によって維持されている。その増加率の差違と移住、混血の進展により、数世紀後の人種地図はいまと大幅に変わることであろう。
皮膚の色は紫外線に対する適応の結果であるから、これによる人種差別は無意味である。しかし、人種偏見は有形無形に現れており、これに対する反発も根強い。楽天的な人々は、未来の人類社会には、今日みるような人種はなくなり、すべてが混融するであろうと考えるが、人種差別の撤廃は、あくまで理念であって、現実には今後も人種および人種偏見は残る可能性がある。それというのも、人類は身体についての関心を強くもつからであり、そのことは正義の問題とは無関係である。しかし、科学技術の発達により、風土に対する不適応はかなり補われるであろう。問題は人種概念の不明確さであり、加えて政治的な曲解を恐れて、最近の人類学者は人種そのものに触れることを避ける風がある。一方、遺伝関係があいまいなところから、遺伝学志向の強い一部人類学者は、人種という概念を認めようとしない。しかし、従来の人類分類表をそのまま受け入れることはできないが、現実に身体上の類似性をもつ人類集団がおり、それが地域的に分布する現状を無視することはできないであろう。
[香原志勢]
『寺田和夫著『人種とは何か』(岩波新書)』▽『寺田和夫編『人類学講座7 人種』(1977・雄山閣出版)』▽『富田守編『人類学』(1985・垣内出版)』▽『NIRA(総合研究開発機構)編『世界民族問題事典』(1995・平凡社)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
従来は皮膚色などの外的な身体特徴から現代人を三大人種から200ほどの人種に区分することがあったが,遺伝学的研究の発達に伴って,現代人集団にみられる変異は連続的であり,集団差よりも個体差が大きく,さらには従来の人種区分と遺伝的変異は必ずしも相関しないことが認識されるようになった。したがって,人種を生物学的に定義することはきわめてむずかしい。一方,旧来的な外的基準と文化・社会的類型化の双方にもとづく人種概念は存続しており,生物学的根拠のない差別意識の原因ともなっている。
出典 山川出版社「山川 世界史小辞典 改訂新版」山川 世界史小辞典 改訂新版について 情報
Sponserd by ![]()
出典 旺文社世界史事典 三訂版旺文社世界史事典 三訂版について 情報
Sponserd by ![]()
…これはレース新生のためであることが多い。レースraceとは形態が同じでありながら病原性の異なる菌で,日本には今16以上のイネいもち病菌レースが知られている。 いもち病の発生程度は年次によっても異なり,ほぼ10年周期で大発生が記録されている。…
…優秀性,均一性,永続性が遺伝的に保たれ,固有の特性によって他と区別されるような一群の農作物や家畜をいう。英語では作物の場合cultivarまたはvarietyといい,家畜の場合はraceまたはbreedという。農林水産業でとくに重視される実用的概念である。…
…これに対し,左右対称に発生する器官は,呼吸器と泌尿生殖器である。
[男女差と人種差]
男性と女性の体には性差がある。内外の生殖器の違いを一次性徴といい,それ以外で思春期以降に現れるものを二次性徴という。…
…すなわち,シャンスラード型のエスキモーとの類似がテスチューによって強調され,グリマルディ型とネグロとの系統関係の親近性がベルノーにより示唆された。しかし今日では,これら3型間の形態的相異は,ヨーロッパの化石現生人類のいちじるしい変異を反映するものと理解され,ヨーロッパ,西アジア,北アフリカの後期旧石器時代人は現存するコーカソイド大人種の原型とみなされている。 化石現生人類は,ヨーロッパ,西アジア,北アフリカだけではなく,それ以外の旧大陸各地,また旧人段階までは人類の足跡が印されたことのないアメリカ大陸やオーストラリアからも発見されている。…
…すでに死滅したヒト科の成員をも併せて広義に人類と呼び,また現存する人類以外を一括して古人類と呼ぶこともある。現在,地球上に生息する人類を,形態学的な差異あるいは遺伝学的特性に基づいていくつかのグループに分けて人種と呼ぶが,人種の起源や人種間の系統的類縁については明らかにされていない点が多い。また人類を文化の差異によって分け,同じ文化を共有する人々をまとめて民族と呼ぶ。…
※「人種」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
夏の暑さに体が慣れること。数日から数十日間で起こる短期暑熱順化と、数年または数世代にかけて起こる長期暑熱順化とがある。→寒冷順化[補説]近年では、冷房設備の普及にともない短期暑熱順化が起こりにくくなっ...