長大な時間経過に伴い生物が変化していくことをいう。生物の形質(形態・生理・行動など)が生息する環境に、より適合したものになる、既存の種から新しい種が形成される、単純な原始生命から複雑多様なものへ変化する、などがその変化の内容である。対象とする変化の内容により、小進化、大進化と区別することも一般に行われている。なお、宇宙の進化といったように生物以外の現象に対して適用されることもある。
生物進化の観念は、現存する個々の生物の種が、天地創造以来、不変のままに存続している(特殊創造説)とか、地表でたびたび大異変がおこり、そのつど新しい生物が創造された(天変地異説)といった、神による生物の創造の観念を排除したところに成立しており、生物の連続性と可変性に対する認識が根本にある。進化論の確立者であるC・R・ダーウィンが、「変化を伴う由来」ということばで進化を表現した理由もこのあたりにある。進化についての先駆的な考えは、18世紀以来繰り返し出現している。これは、生物学的事実の蓄積に伴い、神による創造では説明できないほど著しい生物の多様性が明らかになってきたことや、その一方で、その根底に斉一性(共通性)が存在すること、さらに、物事が変化するとの観念が普遍的になってきたことが認識されていき、そのような状況をつくりだしたのであろう。似ているけれども違うという事実は、生物の連続性と可変性という両面の表現にほかならず、進化論とは、生物の多様性を連続性と可変性という二つの基本仮定のもとに説明しようとする立場にほかならないからである。体系的な進化論は、用不用説や獲得形質の遺伝で有名なフランスのラマルクが初めて提唱し、19世紀なかば、イギリスのダーウィンに至って確立された。ダーウィンは著書『種の起原』(1859)において多くの品種改良の例をあげて生物の可変性を示すとともに、人為的選択によってその変化の方向を左右しうることを示し、ついで、この人為選択に似た仕組みが自然界にもあるに違いないと類推、自然選択説を展開している。
すなわち、彼は、まず生物の多産性という事実に注目し、それは生物の無限の増加をもたらすはずなのに実際にはそうではないことを指摘、数の増加を抑える仕組みとしての生存競争を想定する。この概念を生物の変異性というもう一つの事実と結合させることによって、生存競争の結果、生き残り、子孫を残すのは、わずかでも有利な遺伝する変異をもった個体であるという、自然選択説が引き出されるわけである。『種の起原』の残りの部分では、それまで明らかになった生物学的・古生物学的事実が、自然選択による進化という仮説で、いかに合理的に説明できるかを説いている。進化論が扱うのは、すべて過去におこった現象であり、実験や観察によって直接証明しうる性質のものではない。したがって彼は、生物が進化するとの仮定によって、さらにはその仮定によってのみ合理的に説明しうる事実を多数示すことによって、進化がおこったことを証明しようとしたのである。この事情は、もちろん現在においても変わらない。
一般に進化を証明する事実とされるものには、次のようなものがある。
(1)二つの分類群の中間的形態をもった化石や現生生物が存在する。
(2)変化の道筋を示すような化石系列、現生生物のグループがみいだされる。
(3)分岐をもった階段のように生物を分類することができる。
(4)共通の祖先からの由来を示すような相同器官や痕跡(こんせき)器官が多く存在する。
(5)地域ごとに特有の生物が分布している、などである。
生物が進化するものであるとの考えは、その後の生物学の進歩によってますます確からしいものとなり、一部の信仰的立場の人々を除けば、もはや疑いのない事実となっている自然選択説も、その後の遺伝学の進歩などにより一部修正されたところもあるが、その基本的な考え方は、現代の進化論においても、大ぜいの人の認めるところとなっている。ただし、自然選択説のみでは、進化のすべての面が説明できないとして、新しい仮説の模索も行われている。ここで注意しておく必要があるのは、進化がおこる要因、あるいは仕組みを説明するものとしての自然選択説と、生物は進化するということとは別のものであり、前者がかりに否定されるような事態になっても、後者が否定されるわけではないということである。
[上田哲行]
現在の地球上には、200万を超える生物の種が存在するが、過去に存在し、すでに絶滅したものも含めれば、少なくともその10倍以上(100倍ないし1000倍という推定もある)の生物種が存在していたともいわれる。種というのは、大まかにいえば、相互に区別できないほどよく似た個体の集合であり、ある程度以上の相違があるものは異種とされるわけであるから、実に莫大(ばくだい)な数の異なった生物が存在していることになる。しかし、このような種における多様性は、それらが無秩序に存在することを意味するわけではない。これらの種の間には、その差異性や類似性においてさまざまなものがみられる。たとえば、ウマとシマウマは、その色彩を別とすれば非常によく似ているが、それらはキリンやゾウとはかなり異なっている。しかし、その違いは、ウマとスズメの違いほどは大きくない。さらにウマとスズメの違いも、タコとの違いを前にすれば小さくなってしまう。このように類似性や差異性の程度に基づいて生物を段階的に分類することができるのは、どうしてであろうか。
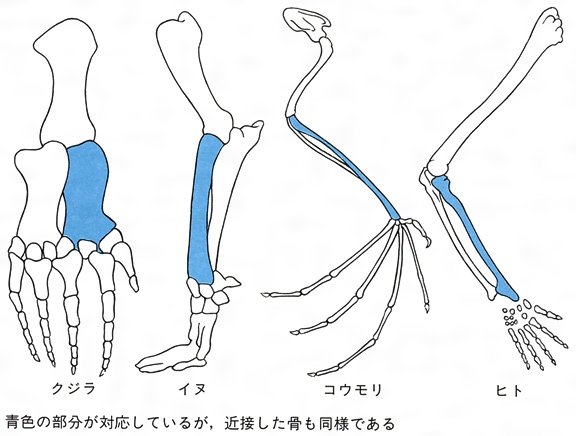
イルカとサメのように外見的類似にかかわらず、体のつくりにかなりの違いがみられる場合もあるが、一般に外見的によく似た生物は、その体のつくりも似ていることが多い。また、外見的にかなり異なったものでも、体のつくりが驚くほど似ている場合もある。たとえば、ヒトの手とチンパンジーの手は、その外見においても骨格においてもよく似ているが、それらは、外見的にはまったく異なるコウモリの翼やクジラのひれとも骨格やその位置において驚くほどの共通性がみられるのである。骨格を詳しくみれば、これらの動物の前肢の外見的相違は本質的なものではなく、特定の部分の骨の相対的長さの違いや、ごく一部の骨の欠落・付加といった部分的な相違にほかならないことがわかる。この例のように、異なる生物において、構造や体における位置関係などで類似が認められる器官を、互いに相同であるという。相同な器官は、胚(はい)発生の過程においても共通しており、同一の胚葉から出現してくる。これら相同器官の類似性は、それらが共通の祖先の器官に由来するためだと考えられている。つまり、共通の祖先に由来する器官が、それぞれの生物の生活上の役割に応じて部分的に変化することによって形成されたとみるわけである。もしも、それらが共通の祖先の器官に由来しないのであれば、ヒトの手とコウモリの翼のように、機能も形態も違う器官が同じような骨格をもつ必然性を説明しなければならないが、それは非常に困難であろう。
相同器官の例としては、肺と硬骨魚類のうきぶくろ、多様に変化した昆虫の口器の相同性などが有名であるが、それ以外にもさまざまなものが知られている。また、相同器官のなかには、クジラの後肢などのように成体では痕跡的に残っているにすぎないものや、魚類のえらに相当する哺乳(ほにゅう)類の鰓裂(さいれつ)のように、成体では完全に消失し、発生の初期にのみ一時的に出現するものもある。また、ヒトの盲腸の虫垂や耳殻筋のように、ほかの哺乳動物では有用だが、ヒトにおいては不用と考えられる相同器官もある。このような痕跡器官や不用器官の存在もまた、生物が進化の産物であることの証拠となりうるであろう。
相同器官の存在は、このようにそれらの生物の共通の由来を示すものと考えられる。哺乳類にも、一時的にえらに相当する器官が出現するということは、哺乳類と魚類の共通祖先の存在を示唆していると考えられる。また、両者の間には、うきぶくろと肺、背骨など多数の相同器官がみいだされる。相同器官が共通の祖先に由来するものであれば、それをより多く共有するものほど類縁が近く、それが少ないほど類縁が遠いと考えることは自然であろう。このように相同関係をもとにして生物の系統をたどることができる。すなわち、哺乳類は爬虫(はちゅう)類から生じ、爬虫類は両生類から、両生類は魚類から生じたというぐあいに、進化の道筋をたどることができるわけである。もちろん、魚類は現在も多数存在するのであるから、魚類のすべてが両生類に変化したわけではなく、魚類の一部の種が両生類の祖先になったという意味である。長い進化の過程においては、祖先種から多数の新しい種が分かれ、分かれた種からまた新しい種が分かれるということが繰り返されてきたに違いない。こうして生物の進化を、時間の経過にしたがって図に表せば、それは1本の木に似たものになるであろう。それを系統樹とよぶ。植物や動物は、その木のそれぞれの太い枝であり、それぞれがさらに細かく枝を出し、脊椎(せきつい)動物、哺乳類、霊長目といったように、より小さな分類群を生み出していくと考えるわけである。
爬虫類から鳥類への移行を示すといわれる始祖鳥のように、異なった分類群の中間的形態をもった化石がみつかれば、両者の系統関係はより直接的に示されることになる。現生生物でも同様な例がある。たとえば、軟体動物に属するネオピリナNeopilinaには体節構造が残っており、軟体動物と環形動物の類縁の近さを示す証拠と考えられている。しかし、このような直接的証拠が得られる例は、むしろ少ないのが実状である。
なお、共通の祖先に由来する生物に相同器官がみられるのは、その器官を生じさせる遺伝子の一そろいが、すこしずつ変化しながらではあるが、そっくり受け継がれているためと考えられる。同様のことは、さまざまな形質を支配する遺伝子についてもいえるわけで、遺伝子組成の類似度から系統を構成する試みも行われている。
[上田哲行]
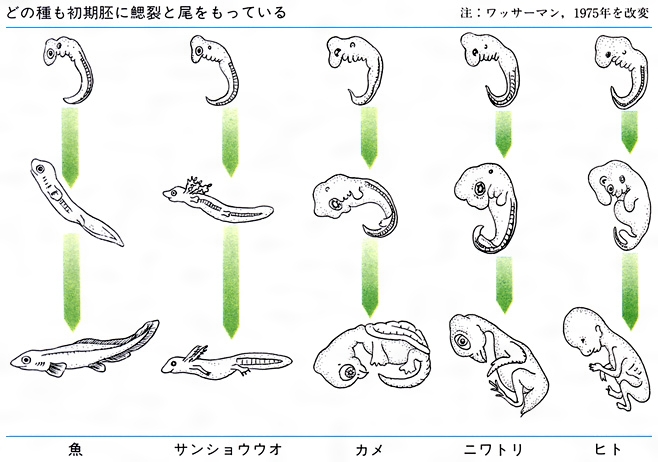
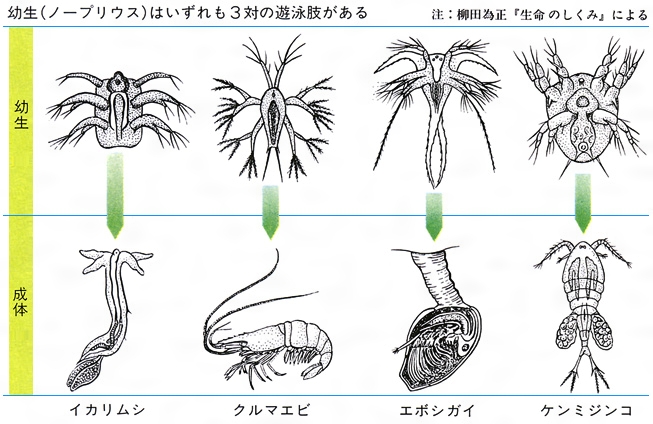
生物の示す時間的変化には2通りある。一つはいままでみてきたような、種の進化的変化であり、系統発生とよばれる。これに対して卵から成体になる過程を個体発生という。進化で新しい種が生じる場合には、個体発生が変化するわけであるから、個体発生と系統発生の間には密接な関係がある。哺乳類の個体発生の途中に鰓裂がみいだされるように、幼生形に祖先動物の形態が残っていると思われる例が少なくない。また、いろいろな動物の個体発生の過程を比較してみると、成体における著しい相違にもかかわらず、その幼生が非常に似ている場合があって驚かされる(図A・図B)。このような事実からE・H・ヘッケルは、1866年に「個体発生は系統発生の短縮された急速の反復である」という生物発生原則あるいは反復説とよばれる仮説を提唱した。そして、この仮説に基づいて、現生生物の祖先型を類推することも行っている。
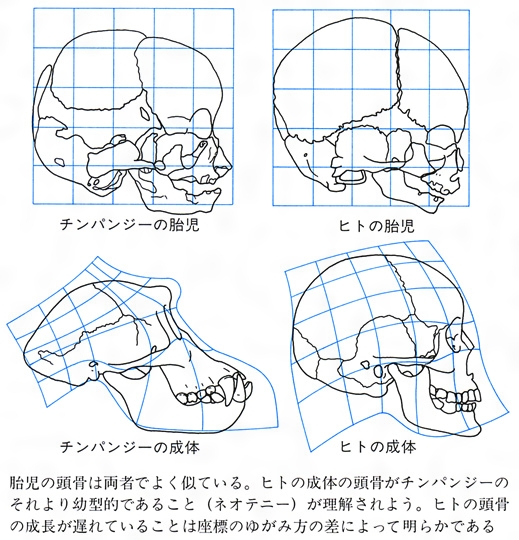
しかし、この原則が成立するためには、つねに新たな変化が個体発生の先へ先へと付け加わっていく形で進化がおこる必要がある。しかし、発生の途中で成熟して成体になり、その先の過程が失われるという形の進化を考えねばならない場合もあり、個体発生と系統発生の関係は、ヘッケルがいうほど単純ではない。なお、後者のような変化をネオテニーneotenyといい、昆虫類は多足類の、脊索動物は棘皮(きょくひ)動物のネオテニー形より進化したとの説も有力である。ヒトがサルや類人猿の胎児型として生じたという胎児化説も、ネオテニー的変化を想定したものである(図C)。
[上田哲行]
新しい種はほかの種に由来するという考え方を推し進めていくと、究極的には、全生物の共通の由来という問題に至る。では、それを示す証拠はあるのだろうか。いろいろ知られているが、もっとも重要なものは遺伝コードの共通性であろう。生物の遺伝情報は、核酸分子中の塩基の配列という形で伝達されるが、それを意味あるものとして読み取るための暗号規則、すなわち遺伝コードは、これまで研究された全生物に基本的に共通しているが、ミトコンドリアDNA(デオキシリボ核酸)ではすこし違うことがわかっている。したがって、たとえば人間のホルモンであるインスリンの遺伝子を大腸菌の遺伝子の中に組み込んで、インスリンを大量生産するなどといった遺伝子工学の手法が成立しうるのである。さらに重要なことは、遺伝コードの決定における偶然性という点である。たとえば、リボ核酸(RNA)においてウラシルという塩基が三つ並んだものは、フェニルアラニンというアミノ酸を意味するコードであるが、それがフェニルアラニンを意味する必然性はいまのところどうしてもみいだせないのである。いいかえれば、それがほかのアミノ酸を意味するコードであってもすこしの不思議もないのである。したがって、遺伝コードはまったく偶然に決定されたとしか考えられず、そのような偶然の一致がすべての生物で生じる可能性は皆無であろう。結局、現生生物のすべては、そのような対応関係を偶然につくりあげた単一の生物を共通の祖先とし、そこから派生してきたものだと考えざるをえないのである。
では、最初の生物はどのようにして出現したのか。いわゆる生命の起源の問題であるが、これについてはさまざまな仮説が乱立しているというのが現状である。原始生命は地球外から飛来したとの説もあるが、それは直接の解答にはならない。地球上における生命の起源を最初に本格的に考察したのはオパーリンである。原始地球上において、もっとも簡単な炭素化合物から、アミノ酸、核酸塩基、炭水化物などがまず生成され、ついでこれらの物質から、生命現象にもっとも基本的であるタンパク質や核酸が生成される。そして多くのタンパク質や核酸が寄り集まって、外部から相対的に独立し、自己複製能力をもった原始生命が誕生したというものである。このような大筋は多くの研究者によって認められ、実験室内で原始地球の大気成分に似た状態からアミノ酸や尿素などが合成されたりもしている。単純な化学物質から複雑な分子が形成され、やがて生命の出現に至る過程は、とくに化学進化とよばれている。地球はおよそ45億年前に誕生したと考えられているが、最初の生物化石は約30億年以前の地層にみいだされている。地球上においても約15億年にわたる化学進化の時代があったということになる。
[上田哲行]
進化の様相には分岐(クラドゲネシスcladogenesis)と向上(アナゲネシスanagenesis)の二つの側面がある。分岐というのは、種が二つ以上の新しい種に分かれることであり、そのもっとも顕著な例は適応放散とよばれている。向上というのは、たとえば、脊椎動物の系統において、体制や機能の面で、より遅く出現した群に内温性がみられるというように、段階を画する(グレード)新たな傾向がみられることをいう。
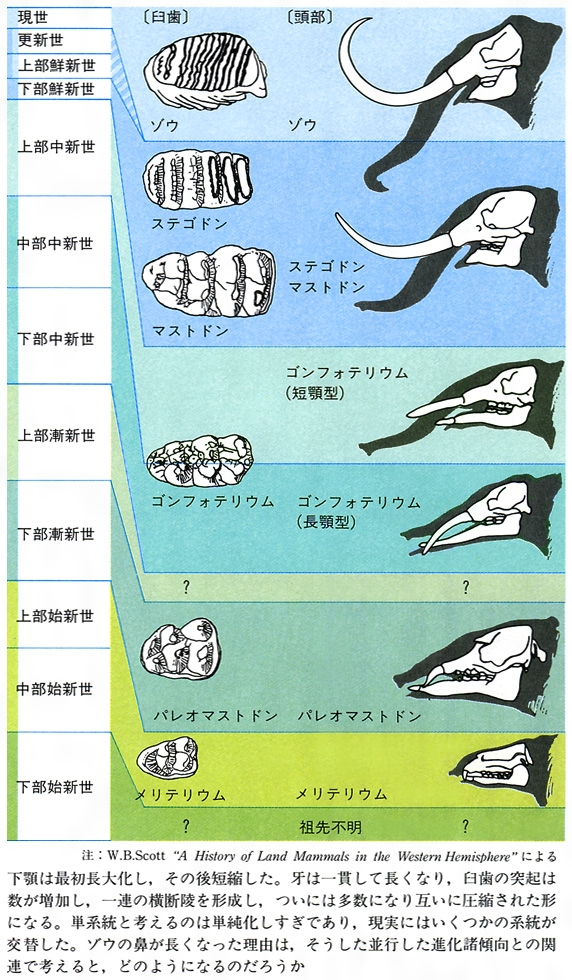
脊椎動物における脳の大きさについても、その体重に対する相対的重量の指数でみる限り、一般に増大する傾向を認めることができる。爬虫類、鳥類、哺乳類と配列したときその傾向は認められるし、哺乳類においても草食獣と肉食獣のそれぞれで、新生代第三紀から現世にかけてその平均相対値も増加する。しかも、どの時期でも肉食獣の脳が大きい。このような向上がどのようにして生じたのかはむずかしい問題であるが、ウマの進化系列においてよくいわれてきた体の増大傾向(コープの規則あるいは一般に定向進化といわれる現象)についてなら、ある種の説明はいくらか可能である。ある個体群に大きさの遺伝的変異があり、その平均値より大きい個体に有利な選択が恒常的に働けば、体の増大化は定着するからである。この場合もいくつかの可能性が考えられる。ただし、より大きな体が生存と繁殖につねに有利だという条件とは、いったいどのようなものかは自明ではない。脳の相対的増大化傾向については、つい知能との関係を想定しそうになるが、肉食獣が狩猟技術の面でより発達した神経系を必要としたという程度の説明なら可能である。ゾウの系統における、下顎骨(かがくこつ)や牙(きば)、さらには鼻や歯にみられる変化の並行的諸傾向の例を図Dに示したが、この説明はむずかしい。
このような変化を記述するのに、トンプソンW. D.Arcy Thompson(1860―1948)は、1917年に相対成長図(図C)を使って瞠目(どうもく)すべき変形理論を提出したが、相対成長として記述される体のある部分と、ほかの部分の相関的なゆがみの進行が何を意味するかは依然として説明できない。それは何かある部分の変化のみが適応的に意味があり、それを「動機」としてほかの部分は単にメカニカルに、力学的安定化を支えるために変化しているのかもしれない。また、ネオテニーの現象と同じく、個体発生上の調節プログラムの別の展開ルートが開けて、その帰結として体のある方向への展開が生じたのかもしれない。それが結果としては生存と繁殖のより成功した方法として採用されたはず、ということになる。
しかし、これは新しい体制や機能の付加については何も説明していない。機械でいえば、部品の改良や大型化を説明できても、モデルチェンジ、つまり、ある種の不連続がどうしてできたのかが問題なのである。これは、F・ジャコブがいうブリコラージュ、あるいはG・G・シンプソンのいう便宜主義がどのように生じたのかという問題でもある。それは、新しい機能を担える素材がたまたま使える形で身辺に「変異」として存在したということなのであるが、その素材が、体の経済学の観点から、さして維持費用のかからないものであったか、即座の転用であったかでなければならないし、あれば便利であるが、現行の体制では獲得しえない拘束についても考えねばならない。適応がどのように「完成」されたかという問題は、変異と自然選択という「制約」のもとに、とりあえずの初期条件を仮定したうえでの説明ならば、たとえば、進化的に安定な「戦略」としてゲーム理論風の説明も可能であるが、新しい条件設定がどのように生まれてくるかという問題なのである。遺伝子レベルでさして費用のかからない変異がどのように生じており、それと新しい表現型との関連が、具体的にとまではいわないまでも、もうすこし実質的に言及されない限り、この問題は依然として宙づりのままである。
[遠藤 彰]
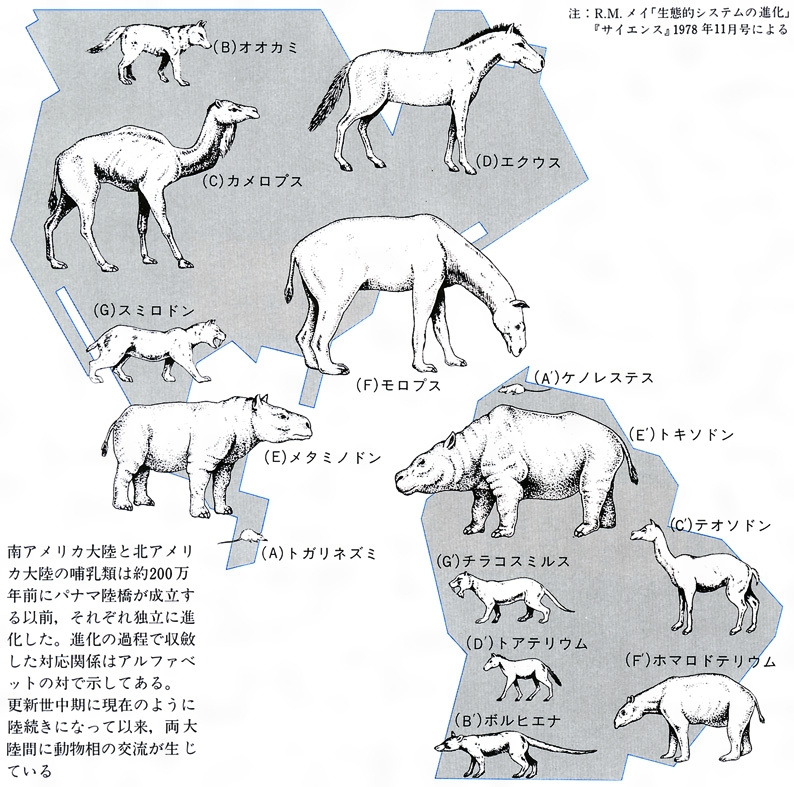
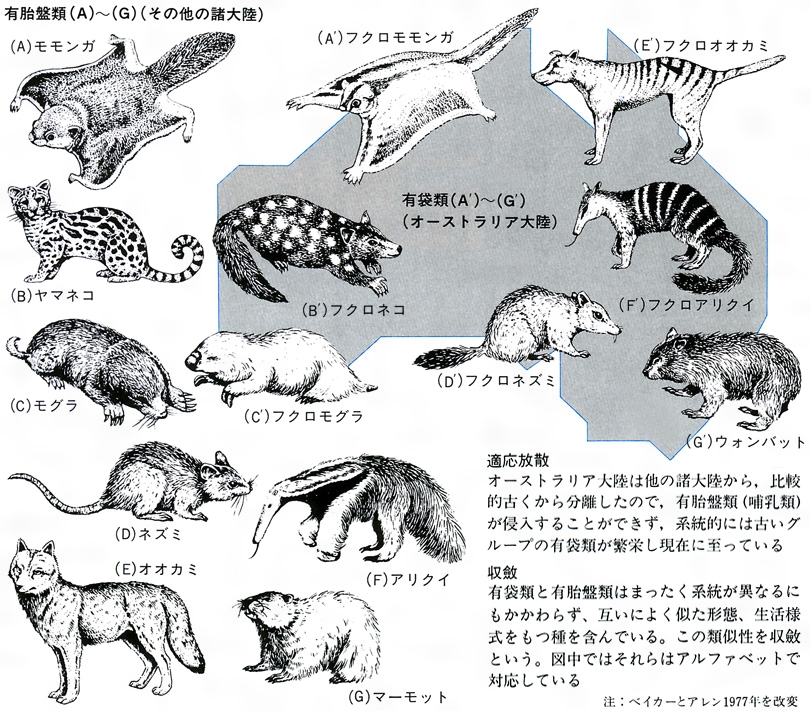
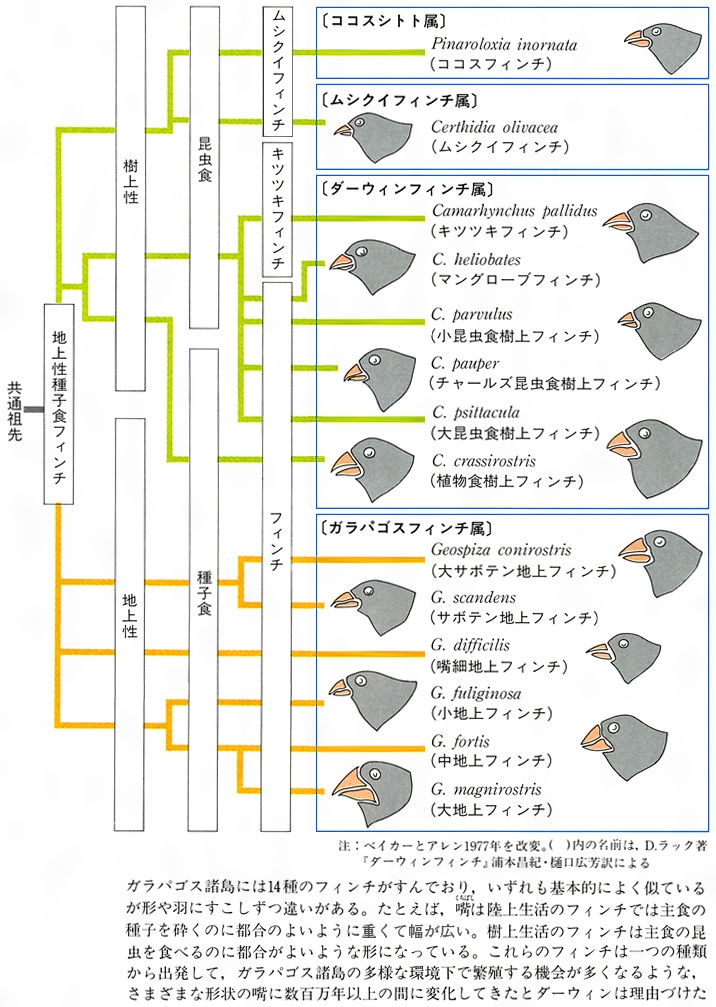
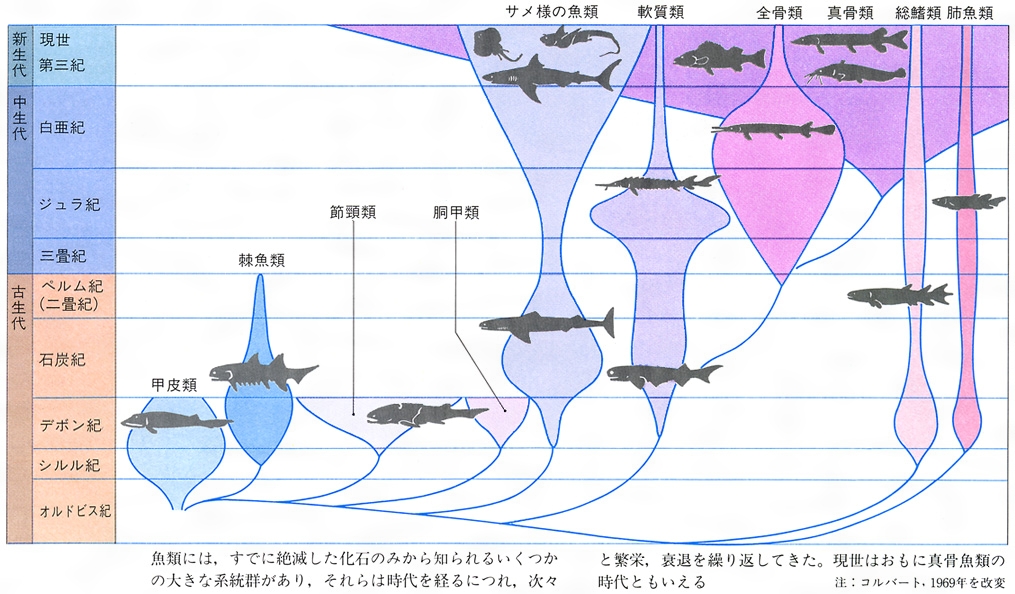
生物の多様性という場合、いわゆる適応放散という現象を抜きには考えられない。この現象は、長い系統の歴史上数多くの例をみいだせる。オーストラリアの有袋類とそれ以外の大陸の有胎盤類の放散との対応関係(図F)はあまりにも有名、また南北アメリカの哺乳類もそうである(図E)。そこには、系統とは関係なく、形態や生活様式の著しく類似した種の対応関係をみいだせる。この類似性は相似といってもよいが、一般に収斂(しゅうれん)現象とよばれている。ガラパゴス諸島のダーウィンフィンチ類(図G)、ハワイ諸島のショウジョウバエ類も著しい多様化を示している。アフリカのタンガニーカ湖のカワスズメダイ類は、少数の祖先種からせいぜい数百万年のうちに多様な種が形成されてきた際だった例である。さらに、魚類の化石は、古生代シルル紀の甲皮類からデボン紀の魚類、節頸(せっけい)類、ペルム紀(二畳紀)から中生代三畳紀にかけての軟質類、中生代ジュラ紀から白亜紀の全骨類、そしてその後現世に至る真骨類の繁栄といった適応放散が繰り返し生じたことを示唆しているとみられている。中生代白亜紀から新生代第三紀の爬虫類の絶滅後の哺乳類の多様化についても同様の進化パターンがみられるだろうし、ほとんどの動物の門(もん)の段階に属するものはこうした適応放散によって出現したとみられている。
では、このような適応放散は、どのようにして生じたのだろうか。これは生物学の超一級の難問である。そうした多様性の記述には、生態的地位(ニッチniche)の概念が有効である。つまり、環境のあらゆる要因とある生物の生活の仕方を、多次元空間における位置で示すことにより、生活様式の多様な分化(分割)の状況を、たとえば、食物源や捕食者、生息場所や運動様式などの生態的・生理的諸要因の関係で差異を明確にして記述するものである。あるものは陸上生活を、またあるものは空中あるいは水中生活をし、さらにあるものは草食に、また肉食になる。そして草食にも、地上、地中、樹上、空中、水中と生活空間は分かれるし、地上生活の草食でも、草食一般から果実食とか若葉食とかに分かれるといったぐあいである。
シンプソンは、適応放散とは、可能性をもつ時と所をたまたま同じくする生物のグループが既存の特徴を再結合し、突然変異させて、可能な生活様式を求めていくことであり、あるグループはある基本的な適応型から出発し、その子孫は分岐する傾向をもち、広くさまざまな習性を身につけるようになる、と述べている。それは、自然選択による秩序づけの原理、すなわちどのように選択圧がかかってきたかをある程度まで推測することができることを意味する。このような生態的地位の分化は、基本的には生態的な競争や協同などの相互作用によってもたらされてきたと考えられている。たとえば形質置換として知られている現象は、そのような相互作用の基本過程とも解釈されている。つまり、近縁な個体群が同所的に分布するとき、その食物のレパートリーや生息場所だけでなく、さまざまな形態的特徴までも変化してしまうことがある。このような形質置換がさまざまに積み重なると、多くの個体群間に表現型のうえでの不連続が生じることになろう。ただし、その適応放散の出発点においては、いわば生態的地位は空白であったわけで、そこにはさまざまな生態的地位に分割される可能性のみが存在していたのである。どう分けるかは気の遠くなるような相互作用の結果として想定されるものである。その結果の一端をわれわれは知っているが、その過程はまだわれわれの想像力の及ばないかなたにある。
生物の示すさまざまな現象は、何から何まで適応しているというのではない。それは分子レベルでの中立的突然変異にとどまらず、体の経済学上の費用、つまりエネルギー配分の問題がさして障害をもたらさないなら、相対成長においてある部分が他の部分の変化の結果として単に変化したにすぎない場合も含まれよう。しかし、そのことと仮説としての適応プログラムをもつこととは矛盾しない。適応にはもちろんいくつかの解答があるわけであり、それは初期条件の単なる違いによって決まっていることもある。時と場所の違いによって異なった自然選択の帰結に至るということである。たとえば、見るとか、泳ぐとか、あるいは飛ぶという機能についての形態的適応を疑うことはできまい(図H)。
しかし、ルウォンティンRichard Charles Lewontin(1929―2021)が指摘したように、サイの角(つの)が1本か2本かという問題になると慎重でなければならない。捕食者に対する適応として、アフリカでは2本がよく、インドでは1本がよかった理由はみつけがたい。これは、たまたま両方の可能性があっただけだというわけである。確かにそうであるかもしれないが、ややこしいことに、これには、角が捕食者に対するものではなく、たとえば種内の威嚇や性的魅力のためのものとすれば、多数派のスタイルに応じて、1本が有利になる場合や2本が有利になる場合をともに想定することもできる、というドーキンスRichard Dawkins(1941― )の反論もある。結局、ある表現型形質をどのような文脈でとらえるのか、これが適応を論じる際に重要な問題となる。つまり、何に対しての適応かということと、後述するように何にとっての適応かという問題がそこにはある。
[遠藤 彰]
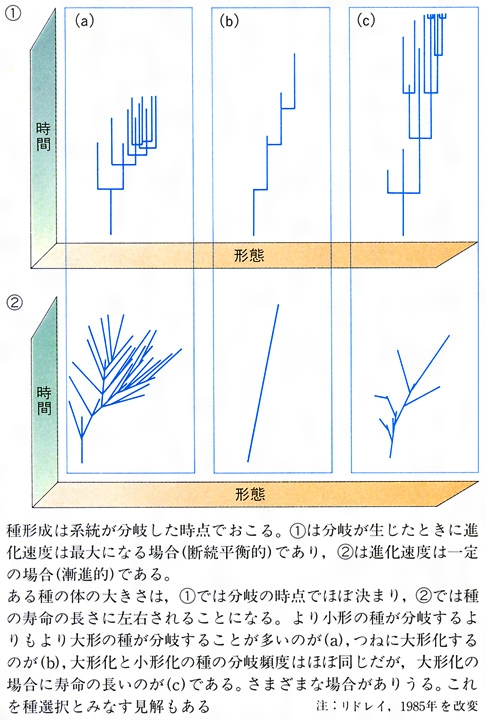
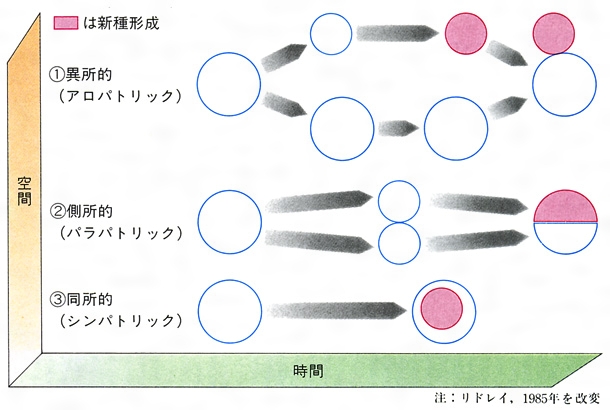
現生生物の数多くの種が共通の祖先から進化してきたとすれば、祖先種は二つあるいはそれ以上の種に分岐したという過程を頻繁に繰り返してきたはずである。その分岐過程を種形成もしくは種分化という。種はどのようにして生じたのだろうか。これは生物進化の基本過程を理解するうえで避けて通れない問題である。新しい種の形成については図Iに示すような三つの基本様式があると考えられている。図Iの①はもっとも広く受け入れられている様式で、異所的種形成(アロパトリック種形成)といわれている。ある生物集団(個体群)が地理的に連続して分布していた段階から、なんらかの原因で、地理的に分集団に隔離された(遺伝子交流を断たれた)状態になり、それぞれの地域で別の自然選択圧がかかり、ふたたび分布域が接することがあっても雑種形成(交配)が不可能なほどに分岐してしまった場合である。集団を分断する障壁にはさまざまな地理的要因、大陸移動や島嶼(とうしょ)化や氷河形成や大洋島への移住などが考えられる。
それに対して、図Iの③は同所的種形成(シンパトリック種形成)といわれ、集団の地理的な分断なしに分岐の生じる場合である。これは同一個体群内になんらかの形で生殖隔離がおこらなければならない。生息場所が異なったり、生殖時期が季節的にずれたり、行動的な多型が配偶関係に影響を与えたりすると、このような分岐が生じる。もうすこし一般的にいうと、同じ集団内に二つの型に有利に働く自然選択が十分強くて、相互の交配を妨げるか、あるいは同系的な交配によって生じた子孫の生存率が、異系交配による子孫の生存率より高ければ、このような種形成も十分可能である。
図Iの②は側所的種形成(パラパトリック種形成)といわれ、前二者のいわば中間的な様式である。個体群がほぼ連続的に接して分布している場合がこれにあたり、多くの地理的勾配(こうばい)のみられるグループの種形成は、これで説明されることが多いと考えられるようになっている。
[遠藤 彰]
分子レベルでの遺伝機構の解明が進んだことから、分子レベルでの進化は、DNAの塩基配列の変化や置換として理解されるとともに、進化における有性生殖の意義の重要性はよりいっそう深く認識されるようになったし、中立的な突然変異の事実も理解され、いわゆる適応万能論に対して、木村資生(もとお)(1924―1994)らによる中立説からの限定が付されることにもなった。この中立突然変異を利用した分子時計が系統分岐の推定に有効なことも認められてきている。ただし、中立説そのものは、表現型の適応そのものを説明するものではなく、遺伝的変異を豊富化する可能性を示すものであろう。
中立説に依拠するとして進化の偶然性のみが強調されることもあるが、それは適応万能論への警告の意義はもつものの、適応の説明とはまた別の問題である。それはともかく、真核生物では遺伝子の重複構造や動く遺伝子(トランスポゾン)の存在も確かめられ、分子遺伝学はいま新たな展開期を迎えつつあり、進化について、豊富な知見をもたらすだろうことは間違いない。
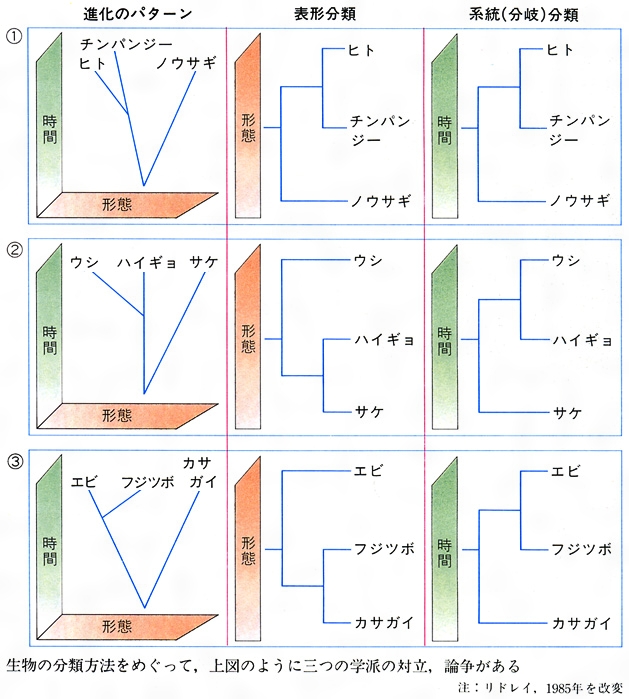
また、系統分類学の方法をめぐって三つの学派の論争が現在も続いている。その一つは、任意に選んだ形質による分類を主観的であるとして排し、数量分類の立場から、可能な限り多くの形質を量的に扱い、類似度を指標にしてグループ分けをする表形(ひょうけい)分類学(フェネティックス)の学派である。
この立場が推論に基づく系統的類縁関係を認めないと主張するのに対して、ヘニッヒW. Hennig(1913―1976)らが唱えた系統分類は、単系統に基づいた分類を主張する。つまり、相同形質について、その形質状態からもっとも新しく形成されたもの――派生形質(子孫形質)をみいだせれば、それを共有する群を単系統の枝(クレード)として位置づけられる。それを繰り返して、より高次の系統分類が可能になるとする(クラディスティックス)。また、系統群により進化過程での分化の程度に差のあることを重視して分類する立場もあり、進化的分類学といわれる。単純化して図Jに示す。表形分類と系統分類は、場合によっては一致するが(図Jの①)、著しい形態差が比較的短期間に生じた場合には異なってくる可能性がある。著しい分岐が生じたり(図Jの②)、収斂(しゅうれん)が生じると(図Jの③)、系統群と表形群に差が出てくる。さまざまな異論があるが、表形分類は、近縁なグループにはそれなりに有効であるが、諸形質をどう測定するか、差異の距離をどう計算するかに問題が残るし、系統分類は、派生的形質の見極めがかならずしも簡単ではない、などの批判もある。といって、従来の分類方法の欠陥は明らかであり、さまざまな折衷も試みられている。
大進化をめぐっては、漸進的変化では説明しにくい現象が化石記録によって示され、種の急激な分岐が生じたのちに長い停滞期が存在するという断続平衡説(区切り平衡説)が、古生物学者エルドリッジNiles EldredgeとグールドStephen Jay Gould(1941―2002)によって1972年に提出され大きな論争をよんでいる。ただし、地質学的時間尺度でいう急激な変化は遺伝学的時間尺度ではかなり長期にわたることから、このことが事実としても、当初主張されたほどには進化の漸進的説明を否定するものではないとも反論されている。それよりも長期の停滞がなぜ生じるかが漸進説には難問かもしれない。種形成(分化)のおこり方については、周辺小個体群における新しい変異の定着が仮定されており、マイアE. Mayrの主張した異所的種形成説への地質学的証拠として位置づけられたものであったことも留意しておいてよい。
ところで、種形成機構については、総合説の打ち出された当時、可能性が低いとされていた同所的種形成も、ランダムな交配ではなく、同系的交配が生じるならば、従来考えられていた以上におこる可能性が高いとみなされてきている。種形成の基本過程は、単に外的要因による隔離だけでは説明しえない現象であることも確かであろう。どのようにして種が分化したのか。これは進化過程を理解するうえでは不可避の論点に違いないが、遺伝子の組換え、中立的なものも含めた突然変異、遺伝的浮動、さらには遺伝子重複などさまざまな変異が総体としてはランダムに生じたうえで、自然選択のさまざまな方向性あるいは規定性が関与したものと考えられている。それはおそらく抽象的にではなく、より具体的に解かれねばならない問題でもあるだろう。
選択(淘汰(とうた))の単位、つまり何にとっての適応かという問題は、ダーウィン以来、漠然と個体にとっての適応が問題にされてきたし、ある意味では、それは当然に思える。しかし、利他的行動(社会的行動)の進化について、1962年にウィン・エドワーズV. C. Wynne-Edwardsがそれを群淘汰(集団選択)によって説明して以来、大論争がおこった。操作上は集団についての平均的適応度を想定することも簡単にできるし、それは限られた状況において意味論的には意義をもつし、理論上も個体選択とかならずしも矛盾するものではないが、大勢としては不要な仮説として退けられることが多い。しかし、では個体が選択の単位としてまったく問題がないかというと、かならずしもそうではない。
ハミルトンW. D. Hamiltonは1964年、利他的行動の進化を説明するものとして、群淘汰説にかえて包括適応度の概念を唱え、いわば同祖遺伝子の適応度をぎりぎりのところで個体の適応度として救い出した。そして、ドーキンスR. Dawkinsは1976年、適応は遺伝子にとっての利益として考えるのが論理的である、つまり、選択されるのは遺伝子すなわち自己複製子であるとの説を唱えた。個体や個体群選択(集団選択)は、ドーキンスによると遺伝子のビィークル(媒体・乗り物)の選択であり、それは遺伝子の選択とは区別されねばならない。適応について語るのであれば、個体ではなく遺伝子にとっての利益として明示的に論じるべきである。包括適応度の概念は、実はそういうことを意味していたのである、という。
生物学的な個体は独特の遺伝子型と表現型をもっている。遺伝子型というのは、その個体の属している個体群の遺伝子プールの構成部分である。そして、表現型が、繁殖成功をめぐってほかの表現型と競争、相互作用する。この成功度すなわち適応度は、さまざまなほかの表現型効果との作用の結果、相対的に決まるものである。つまり条件しだいで、ある表現型が有利になることもあれば、不利になることもある。自然選択は、まさしく外からの秩序づけの原理であり、そうした表現型効果の差異を規定しているが、なんらかの遺伝的差異そのものに働くことになる。そうして減数分裂・生殖を経て、有効な遺伝子の組合せの適合性が高まり、表現型の「進化」がさまざまに進行する。
[遠藤 彰]
『J・モノー著、渡辺格・村上光彦訳『偶然と必然』(1972・みすず書房)』▽『S・L・ミラー、L・E・オーゲル著、野田春彦訳『生命の起源』(1975・培風館)』▽『D・ラック著、安部直哉・樋口広芳訳『進化――ガンカモ類の多様な世界』(1976・思索社)』▽『G・G・シンプソン著、平沢一夫・鈴木邦雄訳『進化の意味』(1977・草思社)』▽『R・ドーキンス著、日高敏隆・岸由二・羽田節子訳『生物=生存機械論』(1980・紀伊國屋書店)』▽『S・M・スタンレー著、養老孟司訳『進化の新しいタイムテーブル』(1983・岩波現代選書)』▽『S・J・グールド著、浦本昌紀・寺田鴻訳『ダーウィン以来』上下(1984・早川書房)』▽『J・チャーファス編、松永俊男・野田春彦・岸由二訳『生物の進化 最近の話題』(1984・培風館)』▽『木村資生編『分子進化学入門』(1984・培風館)』▽『L・マーギュリス著、永井進監訳『細胞の共生進化』上下(1985・学会出版センター)』▽『徳田御稔著『生物進化論』(講談社学術文庫)』▽『C・パターソン著、磯野直秀・磯野裕子訳『現代の進化論』(岩波新書)』▽『西田誠著『陸上植物の起源と進化』(岩波新書)』


 、皆良
、皆良
 天、太(はなは)だ
天、太(はなは)だ ならず
ならず  年
年
 無くんば 大地も合(まさ)に沈淪すべし
無くんば 大地も合(まさ)に沈淪すべし