デジタル大辞泉 「昆虫」の意味・読み・例文・類語
こん‐ちゅう【昆虫】
Sponserd by ![]()
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
動物分類上、節足動物門昆虫綱Insectaに属する小動物の総称である。俗に虫(むし)とよばれる場合は昆虫以外の小動物も含まれる。また、鱗翅(りんし)目のチョウはガを含めて昆虫であるが、虫あるいは昆虫と区別して扱われることがある。別にハエ、アブ、ハチ、アリ、セミ、トンボなど、昆虫の一部をさす語もいろいろある。
昆虫は世界中至る所の陸地および淡水にすんでおり、ごく一部であるが海水中にすむものもある。海にすむものは海岸に近い浅所に限られ、半翅目のウミアメンボ属Halobatesだけが遠く岸を離れた洋上で発見される。熱帯の密林、温暖な森林や山野は多数の多様な昆虫がすんでいるが、極地の荒野、高山、砂漠、洞穴内のような厳しい環境もすべて昆虫の生活場所となっており、氷雪上に現れるセッケイカワゲラ(セッケイムシ)Takagripopteryx nivalisのような種類もあり、それぞれ環境に応じた適応を示している。現在までに記録された昆虫の種類はおそらく80万を超えており、全動物の種数のほぼ4分の3を占めている。また、一地区にすむ昆虫の種類と個体の数から推算すると、地球上の昆虫の全種数は約300万、総個体数は10の18乗にも達するという研究もある。
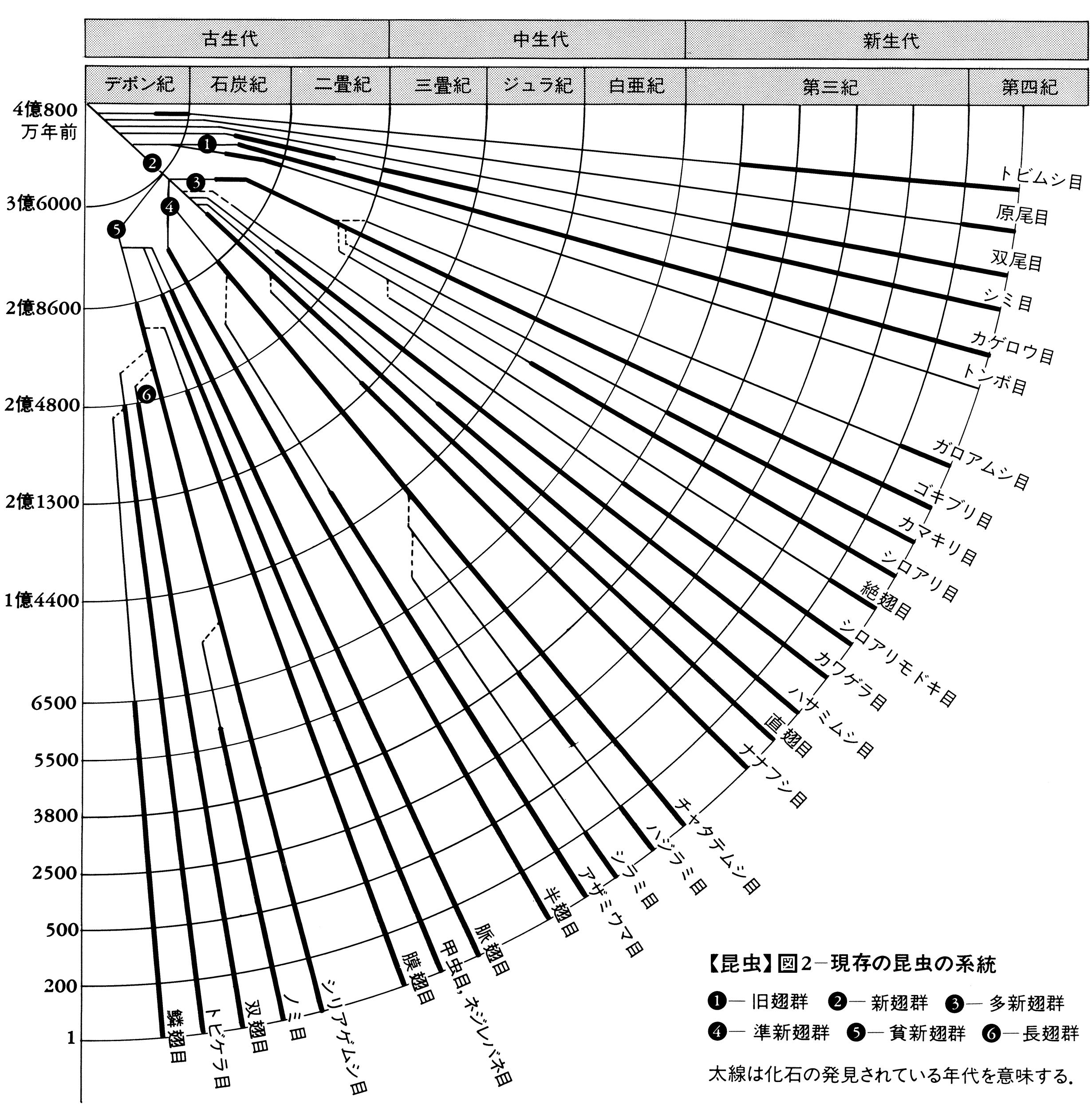
昆虫が地球上に現れたのはおよそ3億5000万年から4億年前の古生代デボン紀のころと考えられ、トビムシ類の1種リニエラ・プラエクルソーRhyniella praecursorの化石が発見されている。次の石炭紀に入って、初めてはねのある昆虫が現れ、2対のはねのほかに前胸両側にも膜状の葉片をもったムカシアミバネムシStilbocrocis heeri(古網翅目Palaeodictyoptera)や、体長30センチメートル、はねの開張75センチメートルに達するコダイトンボMeganeura monyi(原トンボ目Protodonata)などの類があり、ゴキブリ類などもこの時代に出現した。これら有翅昆虫の出現は、翼をもつ爬虫(はちゅう)類や鳥類が現れるのに先だつこと数千万年であって、後者の出現まで昆虫は空中を独占し発展したのである。次のペルム紀(二畳紀)の後期になると、以前に栄えたいくつかの類は滅びたが、現在生存している主要な類がそろって姿をみせ、以後およそ2億年の間、地球上に繁栄してきた。その後、新生代第三紀になると現在に近い昆虫の種類が多くなり、第四紀以後には現存する昆虫とほぼ同じ種類や近縁の類が現れている。現在いる昆虫のなかには比較的近世に種が分化したと思われるものもあるが、大部分は100万年以上前にすでに生存していたと思われる。
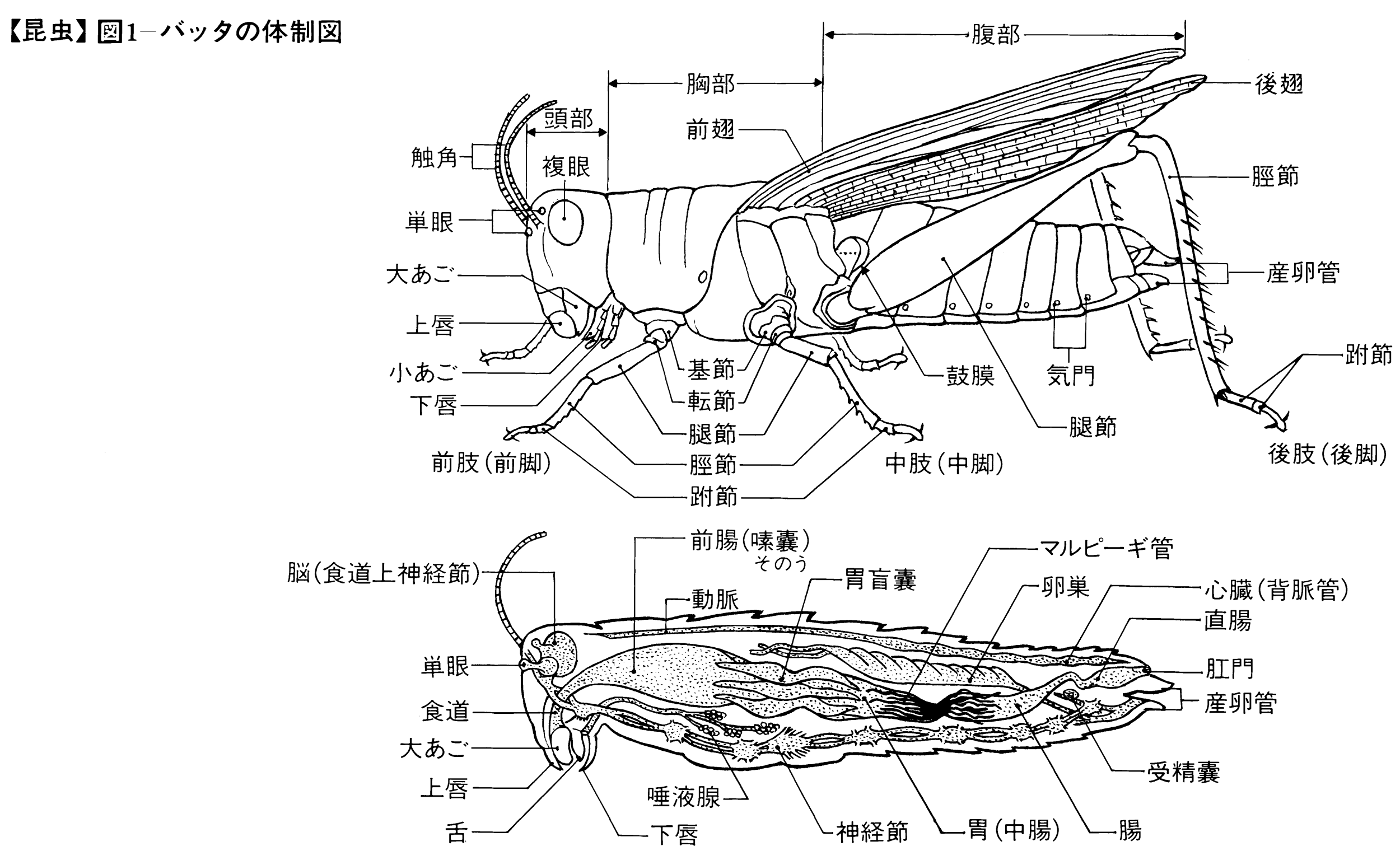
昆虫の体は一連の環節からできており、頭、胸、腹の3部に分かれ、頭には1対の触角それに口器を、胸には3対の肢(あし)と多くの場合2対のはねをもち、腹部には普通末端に消化管と生殖器の開口があり、しばしば尾角(尾毛)や産卵管、あるいは付属器を備えている。昆虫の属する節足動物は体が環節からなっており、付属肢(し)も節からできているが、これらはおそらく環形動物のミミズのように同じ環節が連結した体をもつ動物から進化し、まず各節に1対の付属肢ができ、前端にあって運動や摂食に主導的な部分の付属肢が触角や口器の大あご、小あごに変化し、環節も数個が合着して頭をつくり、後端の肢は尾角や産卵管になり、胸部は発達して運動の中心となり、腹部の肢が退化した結果、昆虫の原形ができあがったのであろう。したがって、昆虫の直接の祖先として考えられるのは唇脚(しんきゃく)類(ムカデ、ゲジゲジなど)や結合類(コムカデなど)に近い形をもつ小動物である。
[中根猛彦]
昆虫のおもな形態と機能について述べる。
[中根猛彦]
昆虫の体は前述のように環節からなっているが、節間部の皮膚は軟らかく膜状で内方へ折れ込んでいることが多い。体節は、背板、腹板と両側にある側板の4部分に分かれ、部分的に互いに接着しているが、節間に硬化部があることもある。このような体節が前後に集合し、頭、胸、腹の3部を形成している。
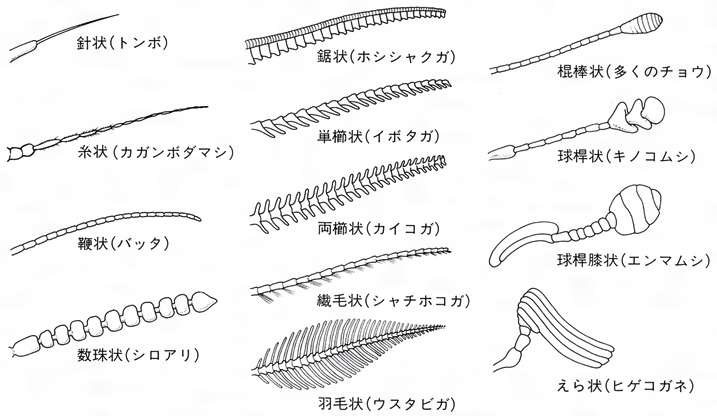
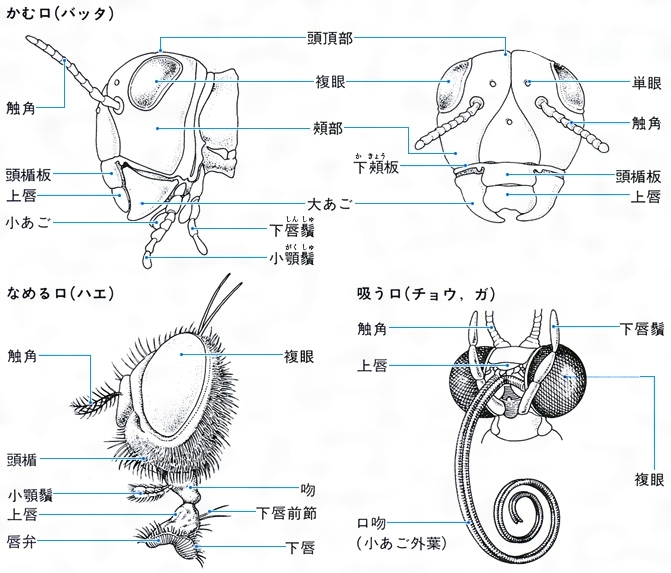
(1)頭 頭部は強く融合した骨片で頭蓋(とうがい)がつくられ、上唇、各1対の触角、大あご、小あご、下唇(左右が合一している)、複眼、それにしばしば1~3個の単眼を備え、頭蓋背部にはY字形の会合線があるのが原則で、背部後方は頭頂、両複眼間は前頭または額(がく)、その前方は頭楯(とうじゅん)とよばれ、前端に上唇があって口器を覆っている。複眼の側方ないし下方は頬(ほお)、頭頂と頸(けい)部は後頭とよばれ、目の後方はこめかみまたは側頭というが、とくに界線はないことが多い。頸部はしばしばくびれている。触角は普通多くの節からなり、糸状、鞭(むち)状、数珠(じゅず)状、鋸(のこぎり)状、櫛(くし)状、羽毛状などと変化があり、先端の節が膨れるか、または広がっているものは球桿(きゅうかん)状といい、先端へ向かって太まる棍棒(こんぼう)状などもある。口器も食性によっていろいろに変化し、かむ口、吸う口、刺す口、なめる口などがある。複眼は細長い円錐(えんすい)形の個眼が密に集合してできている。
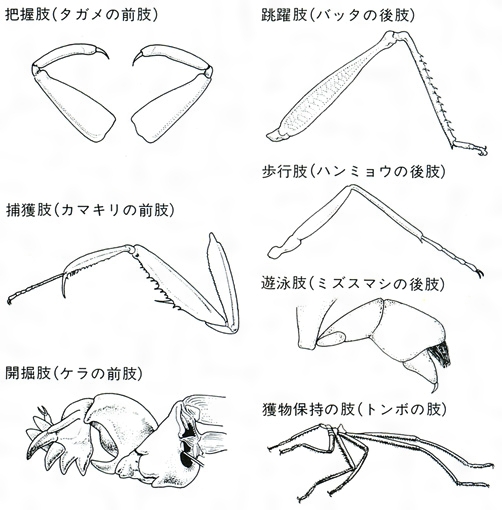
(2)胸 胸部は、前胸、中胸、後胸の3節からなり、各1対の肢(あし)をもち、中胸、後胸の背方側部に多くは各1対のはねを備え、両胸は多少とも密に連合していて、その発達ははねの大きさと活動に関係がある。肢は腹面両側につき、基節、転節、腿節(たいせつ)、脛節、跗節(ふせつ)の5部に分かれ、跗節だけはさらに2~5節に分かれることが多く、先に1~2本のつめと、ときに付属片を備えている。下等なトビムシ、シミなど無翅(し)類のものははねがまったくないが、有翅類の大部分の種類でははねが発達し、一部の寄生性のもの、シラミ、ハジラミ、ノミ、その他では二次的に退化している。また、甲虫、ハサミムシ、カメムシなどでは前ばねが肥厚し硬くなり、双翅類では後ろばねが縮小して平均棍になっている。チョウやガでは鱗粉(りんぷん)が瓦(かわら)状に密にはねを覆っており、アザミウマ類のようにはねが棒状で両側に長い毛を列生するものもある。はねにある翅脈は分類上重要な特徴になるが、長軸にほぼ平行して走る主要な脈と、それらを結ぶ横脈とからなっており、一般に高等な類では脈が減少する傾向がある。
(3)腹 腹部は、後胸に続く後方の部分で、基本的には10~11節からなるが、前後の一部の節は退化したり変形して内部に隠されて外面の節数は少ないことが多い。後端には2本、ときに3本の尾角をもつことがあり、交接の際の把握器など付属器や、雌では突出した産卵管をもつこともある。なお、シミ類では腹節に付属肢の痕跡(こんせき)があり、鱗翅類、シリアゲムシ類、ハバチ類の幼虫は二次的な腹脚をもち、カゲロウ類、カワゲラ類など水生の幼虫には腹部両側または尾端に気管鰓(えら)をもつものが多い。
(4)外骨格 昆虫の体は、外骨格とよばれるじょうぶな硬い皮膚に覆われ、それは外側の表皮(クチクラ)、真皮、内側の薄い基底膜の3層からなっている。表皮はおもに真皮を形成する1層の上皮細胞から分泌され、新たに生成されるときは白くて柔軟であるが、普通はまもなく硬化し着色し、節間の硬化しない膜状部でつながれる。皮膚はこのようにして堅固さと柔軟さを兼ね備えて、筋肉の付着点ともなり、水分の蒸散の防止にも役だつ。表皮の外層は4マイクロメートル以下の上クチクラであって、セメント、ワックス、ポリフェノール、クチクリンの4層からなり、この下に薄層が多数重なって構成された外クチクラ、内クチクラの2層がある。クチクラはおもに窒素を含む多糖類であるキチン質とタンパク質からなっており、両者とも鎖状の分子構造をもち、互いに結合して糖タンパク質をつくっている。少量のポリフェノールがキノンに変化して外クチクラのタンパク質、アルトロポジンをなめしてスクレロチンをつくる。この物質はきわめて堅く、大あごなどはこれによって硬化し、堅い木材や金属にさえ孔(あな)をあけるナガシンクイ、キクイムシなどの甲虫もある。真皮からクチクリン層までは多数の螺旋(らせん)状の微細管が貫通しているが、水分の透過はワックス層で防ぎ、上皮細胞の間には腺(せん)細胞や感覚器の細胞があり、脱皮のときに内クチクラ層を溶かす脱皮液も真皮から分泌する。体表にある突起や毛には、表層からのもの、皮膚の凹凸によるもの、真皮から表皮を貫いて出るものがある。
(5)体色 昆虫の体色はさまざまで、金属色の種類も少なくないが、皮膚の色は色素色、構造色、両者の関係した色の3通りがある。色素色にはメラニン(黒)、カロチノイド(黄、赤、緑など、植物に由来する)、プテリン(白、黄、赤など、シロチョウ類のはねにある)、インセクトルビン(橙赤(とうせき)色、ハエの目など)、アントキサンチン、アントシアン(ともに植物の花に由来する)などがある。構造色には4種類あり、散乱反射による白、波長と同程度の粒子による短波光の散乱によるチンダル青、多層の薄膜や稜(りょう)の反射による干渉色(虹(にじ)色)、多数の密接する細線による回折色がこれである。色素と構造の両方による色にはトリバネチョウなどの金緑色やジンガサハムシの金色などの例がある。
[中根猛彦]
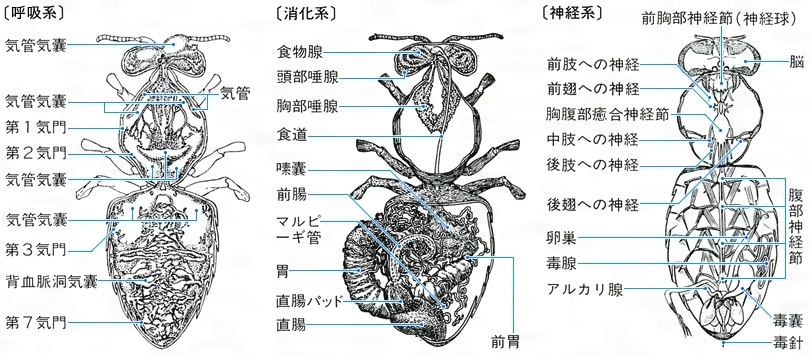
(1)循環系 昆虫の体の内部は広く空所になっていて体液(血液)に満たされ、その流動は背面中央を前後に走る背脈管がポンプの役を果たし、管の前半は大動脈で前端は開き、後半は心臓で数室に分かれ、各両側に心門をもち、後端は閉じている。心臓両側には翼状筋がついていて、収縮すると心臓は膨らみ心門から体液が吸い込まれ、環状筋の波状の収縮で後ろから前へ押しやられる。体液は、栄養物、老廃物、ホルモンなどの移動に役だち、数種の血球を含んでいるが、赤血球はない。
(2)呼吸系 昆虫の呼吸器は気管で、胸腹部の各節両側にある気門から内部に入り込み前後左右に連絡し、また内部の諸器官へ細かく枝分れして達している。気管は表皮が体内へ陥入した形で、内壁は螺旋状に厚くなった上クチクラで補強され、よく飛行する昆虫では各所が膨らんだり、球状の気嚢(きのう)を多数もつものがある。気門は筋肉により開閉できる弁があり、類により一部が消失したり変形したものもある。水生では半翅類のタガメなどのように呼吸管を尾端にもつものがあり、幼虫類はカゲロウ、カワゲラ、トンボなどのように葉片状や房状の気管鰓で呼吸するものが多い。この鰓は内部に細く分かれた気管が入り込んだもので、ここでガス交換を行う。無翅類や水底にすむユスリカなどの幼虫、寄生性のハチやハエの一部の幼虫のように皮膚呼吸をするものもあり、気門のないものもある。なお、腹面に微毛を密生し、ここに空気の薄層を蓄え、気門に導いて呼吸するガムシ(甲虫)やナベブタムシ(半翅目)などもあり、この呼吸をプラストロン呼吸という。
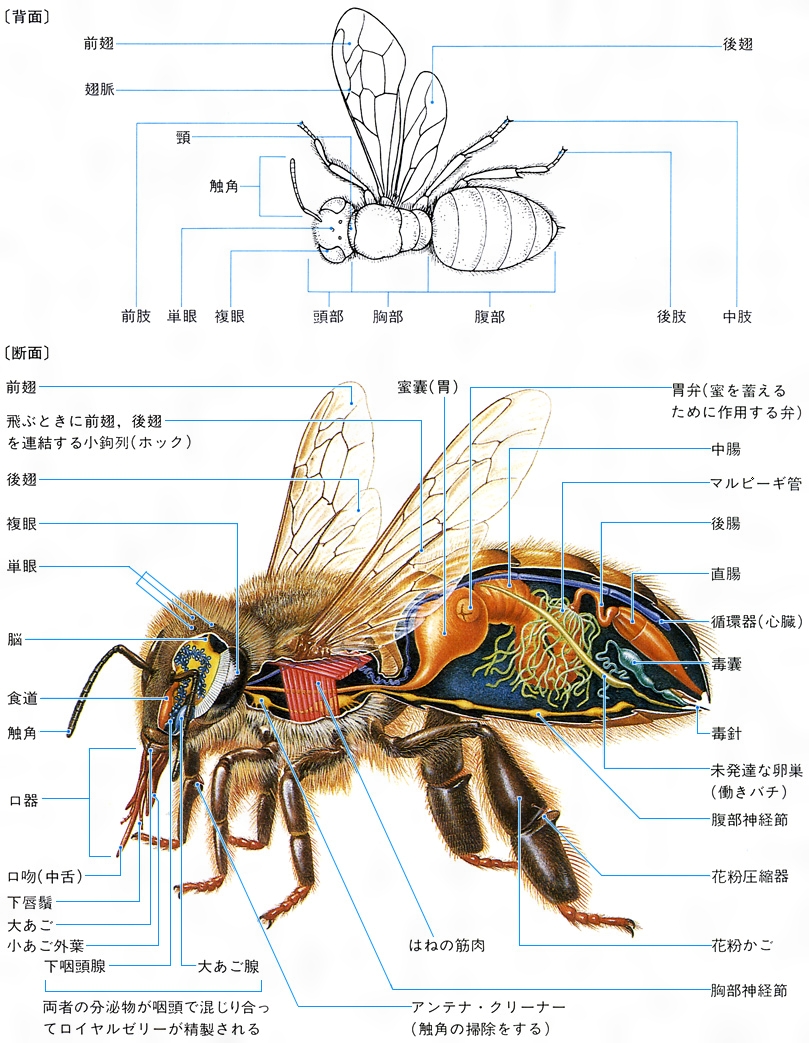
(3)消化系 昆虫の体内中央には消化管が縦に走り、前端は口に、後端は肛門(こうもん)に開き、前腸、中腸、後腸の3部に区別される。前腸には口に続く咽頭(いんとう)と食道の次にそ嚢があって食物を蓄えるところになっており、ハエなど吸食性の類では食道と細い管で連なる袋(吸胃)になる。次は前胃(砂嚢)で壁が厚くて突起やひだが内面にあり、中腸との境に普通、噴門弁がある。中腸は中胃ともよばれ、食物の消化分解はおもにここで行うが、前部にいくつも袋状の盲嚢をもつものや、固形物を胃腔膜(いこうまく)で包んで消化するものもある。中腸と後腸の間は幽門弁でくぎられ、ここからマルピーギ管が体腔内に出ていて、体液から老廃物を集め尿酸をつくり腸内に排出する。後腸には小腸(回腸)、大腸(結腸)、直腸の3部があるが、前の2部分は分化していないこともあり、また大腸に盲嚢をもつものもある。口部には、大あご、小あご、下唇の3腺の存在が知られる。多くは下唇腺が唾腺(だせん)の作用をしているが、鱗翅類の幼虫などでは絹糸腺(けんしせん)として働く。ほかにミツバチには側咽頭腺があり、働きバチの哺育腺(ほいくせん)としてロイヤルゼリーを出す。昆虫の消化酵素はおもに中腸の円筒上皮から分泌され、炭水化物、脂肪、タンパク質をそれぞれ分解するものがあるが、ほかにセルロース分解酵素を出す食材性の種類があり、腸内にそれを生産する細菌(コガネムシの幼虫など)や原生動物(シロアリ類)を繁殖させるものもある。
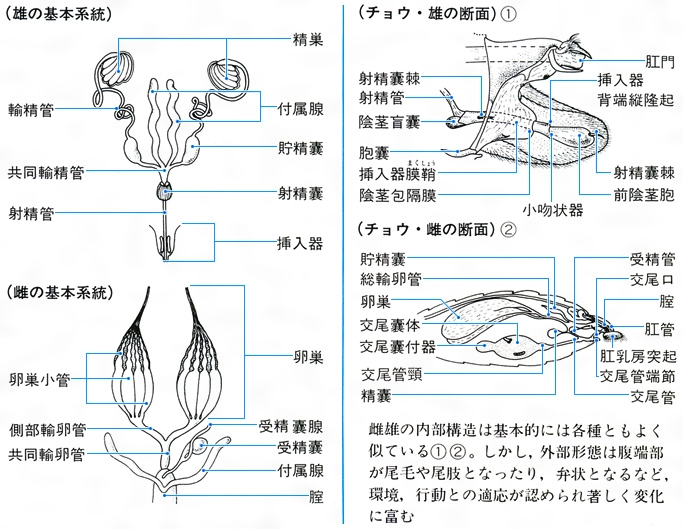
(4)生殖器系 雄では精巣、輸精管、貯精嚢、付属腺、中央の射精管、交尾器からなり、雌では1対の卵巣、輸卵管、貯卵嚢、受精嚢、交尾嚢、産卵管、それに付属腺がある。卵巣は卵小管の集合からなり、卵小管には栄養細胞の有無と位置により、無栄養、端(たん)栄養、交互栄養の3型がある。なお、産卵管はハチでは毒針になることが多い。
(5)神経系 昆虫の中枢神経は基本的には体の腹面中央に沿い、前後に走る1対の平行した神経索と、それによって連結される一連の対(つい)になった神経節からなり、神経節は原則的には各体節にあり、梯子(はしご)状につながれるが、一部ないし大部分の神経節は集合して、高等な類、たとえばハエなどになると集中的になり、頭、胸部に限られてくる。脳はこの中枢神経の前端にあり、3対の神経節の集合したもので、食道の直上方に位置し、食道下神経節とともに頭部の神経中心を形成しており、神経分泌の機能もあって体全体の調節的中心ともなっている。体肢の各部には神経節から神経繊維が延びている。
(6)昆虫の感覚 昆虫が周囲の刺激や状況に反応するのは感覚器が存在するからである。感覚には、触覚、聴覚、嗅覚(きゅうかく)、味覚、視覚などの別があるが、触覚以外は体の特定の部分ないしは局部的に感覚器がある。体表や付属肢に生えている棘(とげ)や毛はほとんどが触覚器で、根元の可動関節部に神経が達している。これらは触覚のほか空気の流れや地面の振動を感じることもある。鐘状感覚器は体表に埋もれていて上端が露出し、クチクラのたわみや緊張、あるいは風圧や水圧を感ずる。弦音感覚器は体や付属肢の内部にあり、外部からの圧力や運動による内圧の変化あるいは振動などを知覚するが、聴覚器として鼓膜があるところには内部にかならず弦音感覚器が付随している。鼓膜はキリギリスやコオロギでは前肢脛節(けいせつ)に、バッタやシャクガなどでは腹部第1節の両側に、またシャチホコガやドクガ、ヤガなどでは後胸部両側にあり、高度に発達したセミでは腹部第2節、発音器腹弁の裏側に位置し、多数の弦音感覚器が鼓膜の内側にある。音ないし空気の振動はほかに感覚毛によっても感知されることが種々の昆虫で知られており、カの雄は触角第2節にあるジョンストン器官で音を感ずるという。なお、この器官は、ミツバチでは空気の流れに反応して飛行速度の調節に役だち、ミズスマシでは水面のゆがみや振動を感じるという。
味覚は口部で感知されることが多く、舌の基部(ハナバチ)、咽頭(鱗翅類の幼虫など)、吻(ふん)の口葉(クロバエ)、口枝(こうし)(口ひげ)の先端部(ゴキブリ、ゲンゴロウ、ガムシ、センチコガネ、コオロギなど)などに感覚器がある。しかし、アリやミツバチ、スズメバチなどでは触角(先の8節)に感覚器があり、肢の跗節と脛節先端に味覚器をもつものにタテハチョウ、マダラチョウ、シロチョウ、ヤガ、イエバエ、クロバエ、ミツバチなどがあり、跗節の下面で砂糖水に触れると吻を伸ばすことが知られている。
嗅覚は、おもに触角に感覚器が密集し、細い毛状あるいは突起状のもの、小さい穴の底に突起があるものがあり、ハチ類やコガネムシなどではほかに板状のものがあり、ミツバチではこれがおもににおいを感ずるらしい。一般には触角にはるかに及ばないが、口枝にも多くの昆虫で嗅覚の存在が知られており、触角を呼吸に使うガムシではもっぱら口枝が嗅覚を受け持っている。なおクロキンバエの雌のように産卵管につく小片に嗅覚突起があって産卵場所を選ぶものもある。嗅覚がよく調べられているミツバチではだいたいにおいて人間と同じ程度に、においを区別できることが知られているが、ガの幼虫などは食草のにおいを感じ、カイコガの雄では雌のにおいしか感じない。この最後の例のように昆虫の出すにおい物質が同じ種のほかの個体を刺激し、行動をおこさせたり規制したりすることは、近年とくに社会性ないしは集合性の昆虫でよく研究されており、そのような物質はフェロモンと名づけられている。
視覚は目によるが、昆虫ではトンボ、ハチ、ハエなどでよく発達している。複眼は多くの個眼が集合して構成されており、個眼は望遠鏡のように細長い円錐形であるので、昆虫の見る像は各個眼に映る像の集合で、電光ニュース板のように対象物の形は正確に知覚できないかわりに、動きはよくわかるようである。各個眼の像は重なる場合もある。ミツバチが、円、三角、四角の図形、X、Y、|||の図形をそれぞれ区別できないことはよく知られている。昆虫の知覚できる色は、人間と違いミツバチでは赤色が見えないかわりに紫外線が見え、黄、青緑、青、紫外の4色を区別できることがわかっているが、一般に紫外線をよく感じることは、モンシロチョウの雌のはねが強く紫外線を反射することや、紫外線を多く出すブラックライトに夜間、ガなどの昆虫がよく集まることからも推察される。チョウなどは赤色も知覚できるという。ミツバチの働きバチは餌(えさ)をみつけると巣に戻って半円形を交互に描くダンスをするが、中央の直進方向は餌の方向(水平面)、または太陽と餌の方向のなす角度だけ垂直線から外れた方向をさすのは、個眼の細胞が偏光板の働きをして視野に明暗を生じ、直進部で餌に向かうときと同じ明暗像を示すことによるという。チョウが好天時に一定の蝶道(ちょうどう)を巡回するのも樹木の葉の反射光の偏光を感ずるためといわれる。
(7)肢とはねの運動 昆虫の運動は主として肢とはねによるが、これは、感覚器が受けた刺激が感覚神経を通じて中枢神経に伝えられ、ここで調整されて運動神経を通じて筋肉に必要な運動をおこさせるのである。はねや肢につく骨格筋は横紋があり、硬い外骨格ないしはそれが内部に入り込んだ部分の2点に関節部または節間部を横切ってついているので、筋肉の伸縮によって関節では屈伸がおこる。軟らかい体をもつ幼虫では、表皮は、体液による内圧と、内面を裏打ちする交差した帯状筋によって補強され、骨格筋の運動を支えている。昆虫の歩行は普通胸部の肢により、基本的には片側の前後肢と反対側の中肢で体を支え、残りの3本の肢を前へ動かすことを交互に繰り返すが、実際には複雑に調整されている。また、水生の昆虫が泳ぐ場合には、ガムシなどを除いて、平泳ぎのように後肢を同時に動かすことが多い。鱗翅類の幼虫は主として腹部の肉質の肢で歩くが、この場合は1体節の筋肉が収縮して、その後部にある肢を持ち上げ前進させる運動が波状に前方へ進み、前進後の肢は垂直方向の筋肉が緩み、体液の圧力で膨らむ。体液の圧力を局部的に働かせることは、カメムシが吻を伸ばすときや、ハエなどが産卵管を伸ばすときにも使われている。
はねの運動は、トンボ類でははねの根元に直接ついた上昇・下降の筋肉によっているが、大部分のほかの昆虫でははねを後方へ畳む筋肉が発達し、はねを曲げたり傾けたりする筋肉は直接はね自身についているが、上下させる筋肉は大部分が胸部の壁についていて、それらが収縮すると胸部の形がゆがみ、間接的にはねを動かす作用をする。よく飛行する昆虫は双翅類(ハエ、アブなど)や膜翅類(ハチ)に多いが、これらでは胸部が一体になっており、前者では後ろばねが縮小して前ばねだけで飛び、後者では前後のはねが連結して動き、前後が分かれて動くものよりも機能的であることを示している。はねは下へ動いてから上へ戻る間に角度を変え、はね先は横から見ると8字形に動く。筋肉の伸縮は飛行と体の大きさによって左右されるが一般にきわめて速く、はねの羽ばたきはチョウで毎秒8~12回、トンボで20~30回、大形のガで50~70回であるが、体が小さくなるにしたがって多くなり、ハナバチ、ハナバエで200回前後、イエカなどで600回ぐらい、微小なヌカカになると1000回以上に達する。このような高速の羽ばたきには、胸部の弾力的な構造とはねの傾きが、羽ばたきの際、自動的に変化するような関節の存在が関係している。昆虫の運動は一般に気温によって左右され、低温になると停止するが、その程度は種や類によって異なっている。夏山の涼しい夜明けに灯火にきたスズメガなどがはねを震わせているのは、胸部の温度を飛行できるまでに上げるためである。
[中根猛彦]
昆虫の卵は、球形から細長い形までいろいろな形をとるが、外側を包む卵殻はしばしば種々の彫刻があり、シジミチョウなどの卵ではきわめて美しい。構造的には昆虫卵は中黄卵(ちゅうおうらん)とよばれる型で、表層に薄い原形質の層があって内方の卵黄を囲み、核はほぼ中央にある。卵殻の頂点には卵孔があり、ここから精子が侵入し受精すると核は分裂を始め、核が一定数に達すると一部を残して表層へ移動し、表層原形質に分裂をおこす。やがて表層の一側に細胞が小さく密な胚条(はいじょう)が生じ、ついで分節ができ将来の体をつくる部分になる。胚体は腹面を外側に向けた形で原頭域と原胴域に分かれ、前者と後者3節が合して頭を、次の3節が胸を、残り11節が腹部を形成する。前端の1節を除き各節に1対の付属肢(し)が生ずるが、第1対は触角を、第3~5対は口器を、続く3対は胸部の肢(あし)をつくる。胚体は表層に沿い両側から背部へ延びて広がり、左右から背中央で合着して虫体を形成し、やがて幼虫として孵化(ふか)する。卵はその発生に必要な栄養は卵の中にもっているが、水分と酸素は外部から取り入れなければならない。これらを表面から得るものもあるが、水分は普通一端にある水孔から吸収され、酸素の取り入れも特別の機構をもつことが多い。乾燥すると発育を停止し長期間耐えるある種のバッタの卵などもあり、また冬季などに備え休眠する卵も多い。卵の孵化のときは幼虫が卵殻や膜をかみ破るものもあるが、ナナフシやカメムシのように卵殻上部が蓋(ふた)状に開くもの、イエバエなどのようにあらかじめ裂け目が用意されているもの、幼虫の頭の頂部に殻を破る歯をもつものなどがある。また、バッタなどのように幼体の腹部第1節両側にある腺から孵化酵素を出して殻の内層を溶かすものもある。卵殻を押し破るときは、幼虫はあらかじめ空気や羊水を飲み込み、筋肉を伸縮して圧力を加える。
[中根猛彦]
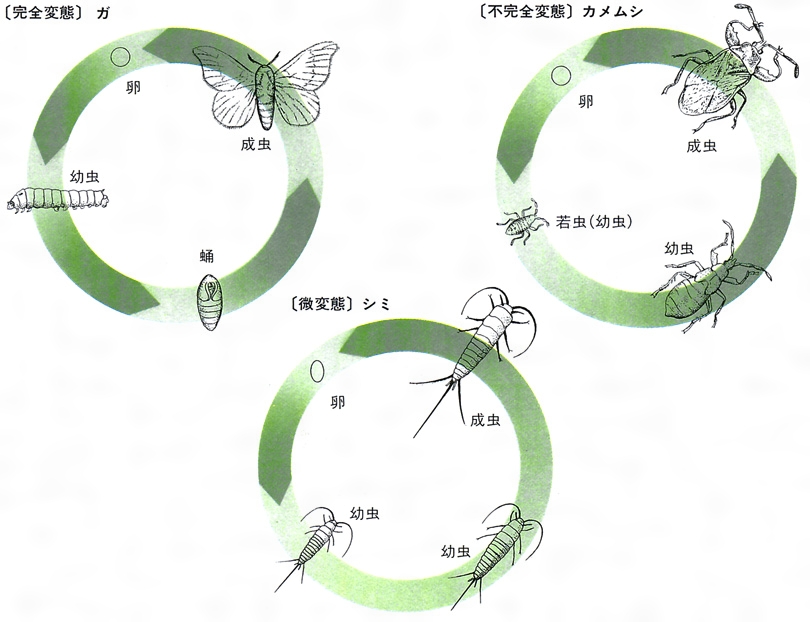
孵化以後の成長は後胚子発生(こうはいしはっせい)とよばれるが、幼虫期の成長はじょうぶな皮膚をもっているので、一般に何回か脱皮を繰り返して大きくなる。幼虫は老熟(十分成長)すると、次の脱皮によって多少とも形態に変化を生ずる。このような成長過程におこる明らかな変化は変態とよばれるが、一般に昆虫では高等な類ほど著しい変態を行う。昆虫の変態は普通、その程度に従って不完全変態と完全変態に分けられるが、シミやトビムシなど無翅(むし)類の昆虫では、成長による形態変化があまりないので不完全変態と区別して微変態とよばれることもある。一般的な不完全変態では、若虫(幼虫)が成虫とかなり似たところがあり、若虫の中期から胸背部の後方両側にはねの芽が現れ、外部生殖器も後端にみえてきて、脱皮するごとに大きくなり、最後の脱皮ではねは完全に延びて成虫となる。完全変態では、これに反して幼虫と成虫がまったく異なっており、はねなどの成虫芽は外面には認められず、幼虫期の終わりの脱皮で蛹(さなぎ)になる。この時期は大幅な体内の改造を伴う静止期であり、次の最後の脱皮によって成虫が現れる。このように幼虫または蛹から成虫になるのが羽化である。完全変態のなかでも甲虫のツチハンミョウやオオハナノミ、寄生バチなどのように幼虫期にも脱皮ごとに形が変わるものは過変態とよばれる。
脱皮と変態の機構は、カイコやサシガメなどの研究によって明らかにされたところによると、脳の分泌細胞から出されたホルモンが前胸腺またはそれと相同の器官を刺激してエクジソン(変態ホルモン)の分泌を促して虫体の脱皮をおこさせるが、このとき体内にアラタ体から分泌されたネオテニン(幼若ホルモン)が十分にあることが必要である。幼虫の終わり近くなるとアラタ体からの分泌が止まり、抑えられていた成虫形質が現れてきて蛹化(ようか)がおこり、続いて羽化がおこる。この間に幼虫の組織は分解され、成虫体が形成される。なお、幼虫とは狭い意味では完全変態をする類の幼期をさし、幼虫から直接成虫に変わる不完全変態類の幼期を若虫とよんで区別することがある。また、カゲロウ類には、若虫から成虫になる前に成虫とまったく同形ですこしはねが曇った亜成虫という時期がある。
[中根猛彦]
昆虫は莫大(ばくだい)な数の種類があり、大きさや形がきわめて多様であるだけでなく、その生態も著しく変化に富んでいる。はねをもたないトビムシやシミなど無翅(むし)類の昆虫では、すむ場所や食性も比較的限られているが、高等な類、とくに膜翅類などに属する昆虫には、きわめて興味深い生態を示すものが少なくない。また、食物も甚だ広範囲にわたっている。よく知られている例をあげてみると、アオムシなど鱗翅(りんし)類の幼虫、バッタなど直翅類そのほかの昆虫、クモ類などを襲って神経節を刺して麻酔し、巣に運んでから産卵し、幼虫の餌にするドロバチ、ジガバチ、アナバチ、ベッコウバチなどのカリバチ(狩蜂、狩人(かりゅうど)蜂)、葉を切り取って巣をつくり、花粉と蜜(みつ)で団子をつくって幼虫に食べさせるハキリバチ、社会生活を営み幼虫を哺育するミツバチ、スズメバチ、アリなど膜翅類をはじめ、卵や若い幼虫を母虫が保護するハサミムシ、モンシデムシ、ツノカメムシ、糞塊(ふんかい)を地中の室に運び入れ、糞球をつくって産卵し、幼虫に食べさせるタマオシコガネ、ダイコクコガネなどの糞虫(ふんちゅう)、葉を円筒形に巻いて中に産卵するオトシブミ、幼虫のため孔内に菌を培養するキクイムシ、アリの巣で生活するクロシジミなどシジミチョウの幼虫やハネカクシなどの甲虫などがある。また、シロアリはゴキブリに近縁であるが、ミツバチ以上に複雑な社会生活を形成しており、地上に巨大なすみか(シロアリの塔)を構築するような種類もある。
昆虫の複雑な生態や行動は、一見それらが意志をもっているようにみえる。ファーブルが彼の『昆虫記』(1879~1907)のなかで擬人的にそれらの生活を描いたのは、むしろ自然であったと思われる。しかし、昆虫の脳の発達からも推定されるように、彼らの行動や生態が実は意志によるのでなく、反射運動の精巧な連鎖によって行われていることは注意されてよいであろう。フェロモンはこの意味で、昆虫の行動上に重要な役割を果たしているものといえる。
[中根猛彦]
フェロモンについては近年の研究によって多くのことが知られるようになった。雄のガが雌に誘引されることは古くからわかっていたが、これは雌の出す性フェロモンによることがカイコガをはじめ多くのガで明らかにされ、類によってはそれが2種の物質の混合で、種の間でその割合が違う場合や、近縁種が同じフェロモンを出すが、それぞれが他種の誘引を防ぐ物質を出す場合などが明らかにされた。ゴキブリが自身の排出物に集まるのは直腸から出る集合フェロモンによるものであり、キクイムシの雌はほかの個体を誘引する集合フェロモンを出すが、またその効果を止める抗集合フェロモンを出して余分な個体がこないようにしたりする。社会性の昆虫では集団生活を維持するため、その調節や規制がとくに重要になるが、仲間に餌のある場所や巣への道を教えるアリの足跡物質(道しるべフェロモン)、外敵に襲われたときに出して仲間を集めたり分散させたりする警報物質(警報フェロモン)、働きバチの性巣発育や王台(新女王幼虫の巣室)の形成を抑え新女王をつくらせないミツバチの女王物質などはよく知られたフェロモンである。シロアリでは階級の分化や集団内の構成員の比率を一定にすることなども一連のフェロモンによって調節されている。昆虫の出すこの種の物質にはほかの昆虫に対しても誘引あるいは防衛の作用などをするものもあり、さらにほかの生物、たとえば菌などが出す物質を生活のなかに巧みに取り入れているものもある。
なお、昆虫の社会は、種族保存の役目や労働の分化が形態的にも現れており、一つの巣の構成員が多くは1匹の女王の子供であって、あたかも1個体の部分であるように行動する点で、高等動物の社会とはまったく異なっている。1巣の個体数は、大きな巣ではシロアリの場合など300万を超えるといわれている。
[中根猛彦]
昆虫の発音は、夏のセミ、秋の鳴く虫でよく知られていて、季節の風物として話題になるが、このほかにも多くの昆虫が発音することがわかっている。鳴く虫は普通、左右の前ばねをこすり合わせ、はねの脈にある刻み目(やすり)や顆粒(かりゅう)列の摩擦によって発音し、膜部に共鳴させるが、ナキイナゴなどのバッタ類では、はねと後肢にある突起列の摩擦で発音する。セミは腹の基部内にあるV字形の発音筋を伸縮させて発音膜を振動させ、腹腔や腹弁を共鳴させる。このほか半翅類や甲虫類などにも摩擦によって発音するものが広くみいだされており、幼虫期に発音するものもある。はねの振動によって発音する昆虫は双翅類のハエ、アブ、カなどや、膜翅類のハチ、バッタ類が身近なものであるが、コガネムシなど甲虫類やカメムシ類もかなりの音を出す。ほかにシバンムシの一部のように頭で壁面をたたくようなものもある。チャタテムシも、その名が、虫の出すかすかな打撃音に基づくという。
[中根猛彦]
昆虫の発光では、ホタルがもっとも有名であって、腹部の後部に発光器があり、ルシフェリンがルシフェラーゼとマグネシウムの存在下で発光をおこすことが明らかにされている。明滅する機構やリズム、色調その他の生態的な面の研究もなされている。また、ホタル類は幼虫期も発光器をもっており、成虫が発光しないマドボタルなどの幼虫も発光する。このほか中央および南アメリカに分布するヒカリコメツキ類Pyrophorusは、前胸の両側と腹部に光の色の違う発光器をもち、よく光ることで知られる。また、半翅類のハゴロモの1種は頭に発光器があり、洞穴性のキノコバエのある種の幼虫や一部のトビムシは体全体が光るという。
[中根猛彦]
昆虫類は節足動物門の1綱として、頭、胸、腹の3部が区別され、触角が1対、肢(あし)が3対胸部にある点で他綱と分けられ、普通はねを2対もつことで特異であるが、原始的で、もともとはねがない5目は無翅(むし)亜綱として、普通はねのある有翅亜綱から区別されている。このうち、イシノミ目はシミ目に含められて4目とすることがある。しかし、無翅の5目(あるいは4目)は互いに明らかに体制が異なり、シミ目が有翅類のカゲロウとやや近いほかは、他の4目(あるいは3目)はかなり類縁が遠いものと考えられ、近年は六脚類Hexapodaという上綱を設け、無翅の4目をそれぞれ綱として昆虫綱と同格に扱う学者もある。有翅亜綱は、はねの芽が幼期に現れ、不完全変態をする外翅類(不完全変態類)と、幼虫が成虫とまったく異なり、完全変態をする内翅類(完全変態類)とに分けられるが、またはねの構造などから、はねを畳むときは垂直に真上に立て、腹部上に畳めない旧翅群と、休止のときはねを腹部の上に畳むことができる新翅群に分け、さらに新翅群を、翅脈、とくに後ろばね内方の翅垂片とその脈の多いか単一かなどの特徴により、多新翅群、準新翅群、貧新翅群(内翅類に相当する)に区分する方式も提唱されていて、一部これを採用することも多い。
なお、近年、昆虫の分類ではネジレバネ目を甲虫目に包含することがある。また、脈翅目は広翅目、ラクダムシ目、扁翅(へんし)目に分割することがある。
[中根猛彦]
現代の都市では昆虫の姿を見ることが少なくなったが、人類が地上に現れたときはすでに昆虫は世界中の陸地に広がって繁栄していた。昆虫はつねに人間の身近にいて、その生活に深くかかわってきた。それは人間の食料にもなり、生活に利用されもしたが、一方では直接間接に人間に被害を与える生物であり、人類の歴史は昆虫との戦いの歴史とさえいわれるほどの敵でもあった。
[中根猛彦]
(1)食用・薬用の昆虫 現代人にはあまり知られていないが、昔は人間にとって昆虫も重要な食料であった。日本の農村でも戦前はイナゴが重要なタンパク源の一つであったし、長野県のような山地帯では名産として知られているハチの子(クロスズメバチ類の幼虫と蛹(さなぎ))をはじめ、テッポウムシ(カミキリムシ類の幼虫)、ゲンゴロウ、ガムシ、カイコの蛹、コウモリガやメイガなどの幼虫と蛹、タガメの卵、セミ、カマキリ、ケラ、コオロギ、カワゲラやカゲロウなど水生の幼虫、トンボなどが食用として利用されてきた。海外でもアメリカ・インディアンがテンマクケムシなどのケムシやイモムシ、バッタ類、水生昆虫やその卵、ハエの蛹などいろいろな昆虫を食用にしていたことが知られている。現在でも東南アジア地域ではタガメの干物や食糞(しょくふん)コガネムシ、その他いろいろな昆虫やクモが市場で売られているという。アフリカやオーストラリアなど熱帯域の原住民の間ではアリ、シロアリ、ミツバチなども重要な食料となっている。ミツバチの蜜(みつ)は砂糖が食用になる前の時代まではもっとも甘い食物として珍重され、養蜂(ようほう)もエジプトや中国などで古くから行われていた。蜂蜜(はちみつ)は食用のほか薬としても用いられてきたが、近年は働きバチの出すロイヤルゼリーも精力剤として売り出されている。現在はほとんど使われないが薬用として利用された昆虫も少なくなく、たとえば、孫太郎(まごたろう)虫(ヘビトンボの幼虫)は子供の疳(かん)の薬として名高く、イナゴ、ゴキブリ、トンボ、カイコの糞なども薬用に使われた。戦後も九龍虫(きゅうりゅうちゅう)(キュウリュウゴミムシダマシ)が強精剤になるとして生(なま)で飲むことが流行したことがある。
(2)昆虫生産物の利用 ミツバチと並び、古くから人間によって飼養された昆虫にカイコがある。養蚕は中国で始められたもので、ヨーロッパや日本にも伝えられ、とくに日本では人工繊維が開発されるまで重要な産業の一つであった。生産量は現在では減少したが、繭からつくられる絹糸は人工にないよさが賞用されている。以前はテグスサン、クスサンなどのガの糸は釣り糸として不可欠のものであり、ヤママユ、サクサン、エリサンなどのガの糸も種々の用途があったが、現在ではナイロン糸の出現によってほとんど使用されなくなった。産業的に利用されてきた昆虫には、このほか、ラッカーの原料となるラックカイガラムシ、カーミン(赤い色素)や臙脂(えんじ)色の染料の原料であるコチニールカイガラムシ、タンニンの原料になるヌルデミミフシ(虫こぶ)をつくるフシバチなどがあり、ゲンセイやマメハンミョウからは発泡剤のカンタリジンが抽出される。しかし、科学の進歩はこれらの薬品を人工の合成物質に置き換えていく状況にある。
(3)天敵の利用 人間の生活に直接間接に被害を与える昆虫は害虫とよばれ、多くの種類があり、これらの防除は農林業をはじめ種々の方面で重要な課題である。害虫を捕食したり、それに寄生して、減少ないしは絶滅させる動植物が天敵であるが、天敵にも昆虫がきわめて多い。そこでこれらの昆虫を害虫駆除に利用することが以前から行われている。ベダリヤテントウが、ミカン類につくイセリヤカイガラムシの駆除のために輸入されて成功したことは有名である。また、戦後にもミカン類の大害虫ルビーロウムシの駆除にルビーアカヤドリコバチが有力な天敵であることがみいだされ、きわめて効果的であった。DDTから始まった殺虫剤の開発と普及で、戦後の一時期は害虫駆除の重点が主として農薬の散布に置かれていたが、薬害により鳥類や果樹などの花粉の媒介に必要なハナバチやハナアブが激減し、害虫の薬剤に対する抵抗性増大や、農薬による土地や水の汚染などの問題がおこった結果、天敵の重要性がふたたび注目され、その増殖や放飼(ほうし)が行われている。
[中根猛彦]
(1)衛生害虫と農業害虫 昆虫のなかには直接人間や家畜を襲って吸血するカ、ブユ、サシバエ、アブ、ノミ、シラミなどの害虫があり、血を吸うだけでなく、病原を媒介することで有名なものも少なくない。たとえば、マラリアのハマダラカ、日本脳炎のコガタアカイエカ、睡眠病のツェツェバエ、ペストのネズミノミなどがそれである。血を吸うのではないが、イエバエ、キンバエ、ゴキブリなどは食物や汚物に集まるので病原の伝播(でんぱ)や媒介の可能性が高い。これらはまとめて衛生害虫とよばれるものである。
家畜や家禽(かきん)に寄生するものは一面で農業害虫ともいえるが、衛生害虫でもある。一方で人類が耕作を始めてから、数多くの昆虫が周囲の野草から作物へ移って繁殖し農業害虫となった。とくにこれらによる害は主食植物の場合は重要で、イネにつくニカメイガ、トビイロウンカ、ツマグロヨコバイなどはよく知られており、最近日本に入ったイネミズゾウムシなども注目されている。ジャガイモの害虫オオニジュウヤホシテントウ、海外で大害を与えているコロラドハムシなども有名な例である。穀類や食料が貯蔵されるようになって、それらを加害する昆虫も現れた。コクゾウムシ、コクガ、コクヌストモドキ、マメゾウムシなどがその例である。このような害虫は本来野外で穀類やマメ類を食していたものもあるが、他のものから転移したものもあるであろう。このほか作物の根を害するもの、蔬菜(そさい)を害するもの、果樹を害するものなどきわめて広範囲にわたり、林業上の害虫にもマイマイガなどのように樹葉を食べるもの、キクイムシなどのように幹や枝に食い入るものなど多数に上る。家材や家具を害するシバンムシ、シロアリなど、干魚や皮革に加害するカツオブシムシ、生薬などの害虫であるジンサンシバンムシなどいろいろのものが知られている。
(2)害虫の防除 人間にとって長年にわたる課題であるが、戦後多くの殺虫剤がつくりだされた。初期に画期的と思われる効果があったDDTやBHCなどは害虫駆除に大きな貢献をしたが、一方では人間に対する毒性や環境の汚染、害虫の抵抗性獲得など種々の問題を生じ、次々と使用が中止され、現在用いられているのは有機リン系では、低毒性のスミチオン、マラソンなどのほか、カーバメート系のカルバリル、プロポキサー、ピレトリン系のアレスリンなどと、植物から得られるピレトリン(除虫菊)、ロテノン(デリス根)、ニコチン(タバコ)である。このほか、害虫の性フェロモンを用いて誘引したり、発生を予察したりして、駆除する方法、フェロモンや変態ホルモンを散布して生殖活動や発生を乱す方法なども試みられている。天敵の利用による防除は前にも触れたが、昆虫に感染または寄生する微生物を用いる害虫駆除も行われている。たとえば、アメリカでは日本から侵入したマメコガネをバチルス・ポピライアBacillus popilliaeという細菌を用いて駆除しており、バチルス・トーリンギエンシスB. thuringiensisは製剤(BT剤)として鱗翅(りんし)類の害虫を駆除するのに用いられている。
最近では、ミカンコミバエなどで成功しているように、このハエを大量飼育し、雄に放射線を照射して不妊化し、これを放し飼いして野外の雌が子孫を残さぬようにする方法も実施されている。このように害虫の防除はあらゆる可能な方法が研究されているが、害虫はもともと人間がつくりだしたものである。人間と昆虫との争いは今後も続くことであろう。
[中根猛彦]
研究のため昆虫を調べるには、まず採集して観察する必要がある。もっとも一般的な採集道具は、捕虫網、殺虫管または毒壺(つぼ)、三角紙とその容器で、目的とする昆虫によっては吸虫管、ピンセット、幼虫採集瓶が必要になる。
チョウは捕虫網でとらえ、網の口を下にし、網の外から胸部を指で押して弱らせ、はねに触れないように注意して三角紙に移すが、ほかの虫は殺虫管に入れる。殺虫管の底には酢酸エチルを脱脂綿かティシュペーパーにしませて入れるのが安全であるが、ガやハエなどは底に青酸カリを入れ、綿や石膏(せっこう)で押さえたものがよい。管にはまた、細く切った紙片か細い葉を入れておくと、虫が中でかみ合ったりしない。軟弱な虫は70~80%のアルコールに浸(つ)けるほうがつぶれないでよいだろう。昆虫はあらゆる場所にいるので、草木の花、実、葉、幹、枝、樹皮下、枯れ木、薪(まき)、キノコ、コケ、動物の死体、糞(ふん)、石や倒木の下、落ち葉の中などいろいろな所を注意してみる必要がある。樹葉上や枝にいる虫の採集には、たたき網を下に広げ、棒で枝をたたいて網上に虫を落として吸虫管で吸い込むのが効果的である。水生昆虫の採集は水網を使用し、草原などの虫の採集はじょうぶな網ですくって歩くのがよい。トンボなど速く飛ぶ虫をとるには網目の粗い捕虫網が採集しやすく、高い梢(こずえ)を飛ぶチョウや高いところの花にくる虫には継ぎ竿(ざお)式の捕虫網が便利である。
ガをはじめ夜行性の虫は夜間採集を行うのがもっとも効果的である。この方法は、展望のきく場所に白布を垂直に吊(つ)るし、その前にアセチレン灯や蛍光灯を下げるのであるが、近年は紫外線を多く出すブラックライトがよく用いられる。街灯や照明の明かりにも注意するとよい。地表性の虫の採集には、紙コップや空き缶を口のところまで地面に埋め、底に糖蜜(とうみつ)(黒砂糖と焼酎(しょうちゅう)を混ぜ、酢を少量加えて煮たもの)を入れて虫を誘引するトラップ法がよく用いられ、糖蜜のかわりに腐肉や糞などを用いる方法もある。このほか土壌や落ち葉の中にすむ小昆虫やダニなどを集めるのに漏斗(ろうと)型のトゥルグレン装置、ハエやハチなどよく飛ぶ虫をとらえるのにかすみ網式のマレーズトラップが用いられる。
なお、採集は単なる愛玩(あいがん)や収集趣味など、研究目的以外は慎まなければならない。安易な採集による乱獲は、自然界の生態系を破壊させる要因となることが指摘されている。したがって、近年の教育学習では自然観察やカメラによる記録が奨励されている。
[中根猛彦]
採集した昆虫は標本にして保存するのであるが、それにはデータが必要である。採集地、採集年月日、採集者、わかれば食草、宿主などをラベルに書き込んで、昆虫を刺した針につけておけばよい。
チョウ、ガ、ハエ、アブ、ハチなどの標本ははねを水平に広げるほうがよいが、これには展翅(てんし)が必要である。胸の中央に垂直に昆虫針を刺し通し、展翅板の溝に刺してはねの根元を板の高さにそろえ、はねを広げて紙テープで板上にはねを押さえるのであるが、原則として前ばねの後縁を溝と直角に、左右一直線にし、チョウ、ガでは後ろばねの前縁をすこし前ばねと重ね下側に入れるが、ハチでは前後のはねを連結する鉤(かぎ)があるので、つなげて展翅する。トンボははねを畳んだまま横向きにしてもよいが、展翅するときは後ろばね前縁を体と直角にしたほうが形がよい。一般に古くて硬くなったものを展翅するには軟化が必要で、缶の底に防腐剤を加えた水で湿らせた砂か脱脂綿を敷き、その上に2~3日置いて軟らかになってから展翅する。チョウの場合は湯を胸部に注射する速成軟化法もある。採集後すぐ冷凍したものは解凍すると展翅できるが、腐食が早いので長期間保存することはできない。展翅を必要としない甲虫やバッタなどは、必要があればコルク板や多泡性樹脂板の上に留め、留め針を用いて肢の形を整えておいて乾燥させるとよい。乾燥してから虫を適当な高さまであげるが、だいたい昆虫針の高さの上から3分の1ぐらいがよい。虫の高さはそろえたほうが多く並べたとききれいにみえる。小さい虫は長方形や長三角形に切った台紙の先に貼(は)り付けるか、待ち針形の虫ピンとよばれる微針で虫を刺し、さらに多泡性ポリエチレンやコルクの小片の一端に針を通し、他の一端に別の針を刺して固定させる。針を刺すと破損しやすいものは台紙に貼るほうが安全である。虫の高さやラベルの高さを決めるために段になった平均台というものがある。普通、昆虫針を刺すのは胸の中央であるが、甲虫では右上ばねの前のほうに刺すのが普通である。
内臓が腐りやすい大形のトンボやバッタなどは、首のところから先の細長いピンセットを刺し入れ、内臓を抜き取り、トンボは竹串(たけぐし)、松葉、ウマの毛などを刺し込んでおき、バッタなどは脱脂綿を詰めておくとよい。内臓を除くのに、腹部の大きいものはその両側の溝を切り開く方法もある。
幼虫はアルコール浸(づ)けにするのが普通であるが、大形のものは内臓を体の後端からもみ出し、そこへガラス管の先を当てて空気を暖めながら吹き込んで乾燥させる方法もある。データのラベルは虫体の下方に刺し、種名を書いたラベルはその下に刺す。アルコール浸け標本の場合は瓶にラベルを貼るか、中に入れるが、鉛筆で書くか、または墨や製図インクを使わないと、ぬれたとき消えてしまう。
標本は標本箱に並べて刺して保存するが、箱は密閉できる木箱がよく、底に厚めの軟コルク板か多泡性ポリエチレン板を接着する。虫やカビの害を防ぐにはナフタレンかパラジクロルベンゼンを入れるが、前者は長もちするが殺虫力はなく、後者は殺虫力はあるが早く消耗する。標本は日光によって色がさめるので、なるべく光を避けたほうがよい。
[中根猛彦]
飼育は生態の観察や生活史を調べるのに必要な方法であるが、鳴く虫なども観賞用に飼育される。飼育にはその状態をできるだけ野外の環境に近くすることがたいせつである。昆虫の生活は種類によって違うので、飼育もそれにあわせ、たとえば葉を食べる虫は、金網を張った飼育箱内に水差しに食草を刺して入れ、水差しの口には虫が落ちないように綿などを詰める。蜜や樹液を吸う虫には薄めた蜂蜜(はちみつ)や砂糖水、スイカなどを与え、適当な足場をつくる。水分の補給には脱脂綿に水を含ませたり、リンゴなど果実を切って入れると、食肉性の虫の場合にもよい。霧を吹いてやるのもよいが、水分が多くなりすぎるとカビが発生して失敗する。物陰に隠れる性質の虫には木片や石で隠れ場所をつくったり、草を植えてやるのもよい。地中にすむアリなどは土を余分に入れ、容器の下方を黒い布などで包む。小さい容器は直射日光の下では温度が高くなるので、日光にさらすのは避ける必要がある。飼育は根気のいるもので、つねに注意を怠らないことが肝要である。
[中根猛彦]
昆虫標本の展示はおもに博物館にあるが、生きた昆虫はインセクタリウム(昆虫園、昆虫館)で展覧している。後者では東京にある多摩動物公園の昆虫園や豊島(としま)園の飼育施設がある。研究機関は博物館のほか、大学の昆虫学研究室、国や県の農業試験場、林業試験場などがあり、また、衛生昆虫は国立感染症研究所、国や県の衛生研究所で研究されている。
[中根猛彦]
『八木誠政著『昆虫学本論』(1957・養賢堂)』▽『井上寛・中根猛彦他著『原色昆虫大図鑑』全3巻(1959・北隆館)』▽『伊藤修四郎・中根猛彦編『原色日本昆虫図鑑』上下(改訂版・1977・保育社)』▽『松本義明他著『昆虫の科学』(1978・朝倉書店)』
ミツバチの体制模式図
昆虫の体の内部(呼吸系、消化系、神経系…

昆虫のクチクラ
昆虫の器官(触角)
昆虫の器官(口器)
昆虫の器官(肢)
昆虫の器官(生殖器)
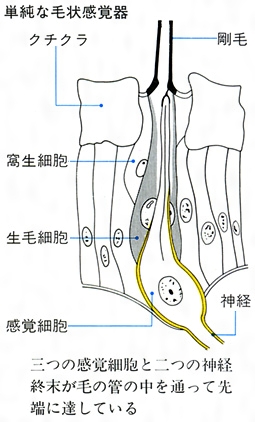
昆虫の感覚器(触覚)
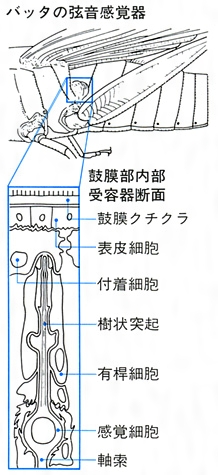
昆虫の感覚器(聴覚)
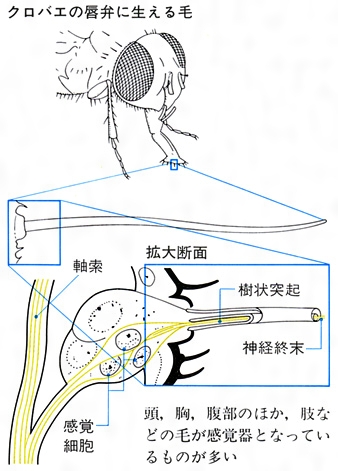
昆虫の感覚器(味覚)

昆虫の感覚器(視覚)
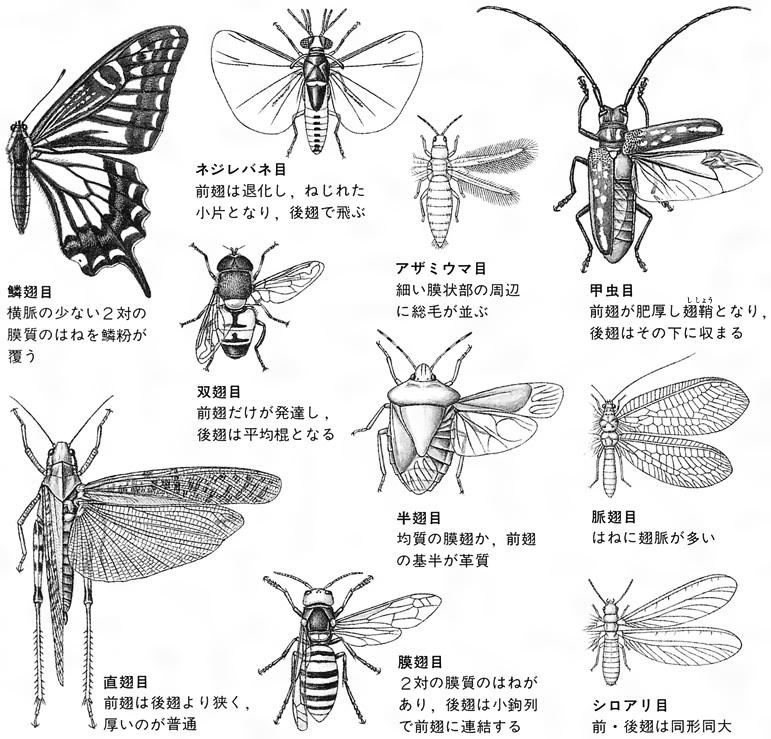
昆虫のはね
昆虫の変態

アゲハチョウ

ウスバカゲロウ

エダナナフシ

オオクワガタ

オオムラサキ

カブトムシ

キイロスズメバチ

キリギリス

クマバチ

ゲンゴロウ

シオカラトンボ

タマムシ

ナナホシテントウ

ニイニイゼミ

ハラビロカマキリ
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
昆虫類は蛛形(ちゆけい)類(クモ,ダニ,サソリなど),多足類(ムカデ,ヤスデなど),甲殻類(エビ,カニ,ミジンコ,ワラジムシなど)などとともに節足動物に属し,体や付属肢は多くの環節からなる。英語のinsectは,分割されたものを表すラテン語insectumに由来する。昆虫類は全動物の種類の3/4を占める最大の生物群であり,総種類数75万とよくいわれるが,毎年発表される莫大な新種の数からみて,実際ははるかに多くの種類を含むであろう。昆虫類はさらに個体数の多さでも大半の陸上動物を圧倒する。このことは日常出会う動物たちを昆虫とそれ以外に分けて数えてみれば容易にわかる。
莫大な種類数に応じて,昆虫綱Insectaは多くの分類群に分けられる。細部については専門家によって見解が異なるが,最上位の分類群である亜綱と目の,現在一般的に認められている区分を例とともに表に示した。昆虫はその大きな特徴である翅の存否によって,まず二大別される。無翅亜綱Apterygotaは今まで翅を出現させたことがなかった類で,このうちシミ目が有翅亜綱の祖先型に近縁とされる。ただし研究者によっては,無翅亜綱の各目は,有翅亜綱と対等の分類群,もしくは昆虫とは独立な群とみなされ,ことに種数,個体数ともに多いトビムシ目は,かなり特殊な別の類だとの見解がある。
有翅亜綱Pterygotaの諸目は表のように分かれており,下方の類ほどより進化した類である。この亜綱にも無翅の種類は多く,ガロアムシ,ハジラミ,シラミ,ノミの諸目は全体が無翅である。しかし,これらの無翅昆虫は,無翅亜綱と異なりすべて有翅の祖先型から退化して翅を失った。上記の最後にあげた3目の場合は,明らかに寄生生活に対する適応とみられる。
表の目の名と例にあげた昆虫の名は,その目の昆虫の中での成功度と関係がある。例があまり聞きなれない名の場合,その目は(トビムシを除いて)多くは種類数の少ない,小型の,または目だたない虫たち,つまり現在あまり成功していないグループである。一方,目の名に昆虫名がついていない,直翅目,半翅目その他は,右側の例を見るとわかるように,形態や生態が変化に富み,一つの虫名で代表させにくい。これらの目は,さまざまな生活様式を開発して多様性を増大させた成功者といえる。目の名に翅が多く用いられているのは,動植物の分類の開祖C.リンネが翅を重視して命名したことに基づく。翅は重要な形質だが,現在の分類にはそれ以外のさまざまな形質が用いられている。
表の最後にあげた脈翅目以下の9目が,卵→幼虫→さなぎ→成虫の生活環を完成させた,もっとも高等な完全変態類である。そのうちの鱗翅目,双翅目,甲虫目,膜翅目と,不完全変態類でもっとも進化した半翅目とが圧倒的に種類数が多い。他のすべての目の種数を合計しても,これら5目のどれか一つの種数にも及ばない。つまりこの5目が,全動物の種数の過半を占める真の成功者たちだといえる。
多様な昆虫の形態と機能を,限られた紙数で解説することは不可能である。ここでは彼らの特性をもう一つの成功者,われわれ自身が属する高等脊椎動物と比較しながら概観してみる。
脊椎動物と節足動物は,感覚・運動能力の優秀さで,動物における二つの頂点をなしている。運動能力の発達は,両者がともに強力な筋肉の付着・支持点となりうる骨格を備えていることに基づく。ただしその方法は正反対で脊椎動物の内骨格に対して,節足動物は外骨格を発達させた。外骨格は運動力の増大のほかに,外敵と乾燥に対する防御機能も果たす。乾燥への対策の重要性は,20門を超す動物群のうちで陸上生活に成功したのはわずか4門(体内寄生者を除く)にすぎないことでわかる。ただし外骨格は成長を阻止するという欠点がある。そのため周期的に脱皮せねばならないのが節足動物の宿命であり,脱皮の失敗および脱皮前後の無防備状態は,彼らの死因のかなりの比率を占める。
昆虫はこの外骨格を異節化によってさらに進化させた。節足動物の体節構造は,その先行者である環形動物(ゴカイ,ミミズなど)でより明りょうに見られる同節性を出発点とする。1対の付属肢と1対の神経球をもつ同型の体節が前後に連なる方式で,多足のムカデなどにまだこの傾向が残存する。昆虫は各体節の変型や消失,体節間の癒着などによって,異節性を強化した。これは体軸に沿って異質な区域をつくり出し,その間に分業を起こさせることであり,それによる効率化の増加は大きい。体が頭,胸,腹の3部分に分かれているという,よく知られた昆虫の特徴は,この異節化の成果である。図示したトノサマバッタの形態を眺めつつその特徴をおってみる。頭部では異節化は極端な癒着を起こし,元来の構成要素である6体節はまったく認められない。わずかに口器としての大あご,小あご,下唇などの付属肢が,かつての体節性の証拠となっている。癒着は頭部内の神経球にも起こり,消化管を前上方と後下方からかこむ二つの大きな神経球にまとめられた。前者は脳として,他の無脊椎動物(イカ,タコを除く)よりはるかに優秀な中枢機能をもち,さらに接近して存在する複眼,単眼,触眼(アンテナ)を支配して情報センターの役も果たしている。胸部はこれに対して脚と翅を備えて運動能力を独占する。異節性は強いが,体節構造は認めうる。腹部は重要な内部器官の収容場所として,外界との直接交渉はあまりない。体節構造はもっともよく残存し,各節が互いに可動性をもち,これが呼吸・循環機能の効率を高めている。
異節性は広義に解して,付属肢に関しても考えられる。口器についてみると,バッタの場合大あごがそしゃく機能を果たし,小あご,下唇は餌の触知や取込みに役だっている。このようなそしゃく口器は,昆虫類における食物摂取の基本的な型だが,一方で口器が変形して管状となり,露出した液体,または動物の体液,植物の柔液など障壁で隔てられた液体の摂取に適した吸収口器も,いくつかの類で出現した。ことに二つの成功群,半翅目と鱗翅目のほとんどすべて,およびもう一つの成功群,双翅目のかなりの類がこの方式に転じたことは注目すべきであろう。運動器官としての胸部の脚も,前脚が捕獲器(カマキリ),掘削器(ケラ),後脚が跳躍器(バッタ),遊泳器(ゲンゴロウ)などに転換し,対応した形態的特化を起こしている例がある。
このような頭や胸の付属肢の機能転換は,昆虫類にさまざまな生活様式を可能にし,莫大な種類数を生み出す一因となったのであろう。しかし昆虫類のもっともユニークな特性は飛翔(ひしよう)能力の獲得にみられる。人間の航空技術を別として,空中に進出できた動物は,鳥とコウモリと昆虫だけである。しかも前2者の翼は前肢の機能転換であり,新機能はもとの機能の喪失という犠牲のもとに獲得された。昆虫の翅は,初めから飛翔器官として発現した点で全動物でも類がない。翅は発育後期まで胸部の囊状突起としてとどまり,不完全変態類では外から認められるが,完全変態類では皮膚下に潜んでいる。成虫になるとともに伸長し,それまで翅芽に酸素を供給していた内部の気管(後述)は,翅脈となって飛翔時に翅を支持する骨格の役を果たす。翅脈の配列は目から種までのすべての分類群の識別に重要な特徴として用いられ,さらに系統発生をさぐるうえでの重要な手がかりともなる。一般的にいって,翅脈相は原始的なグループほど複雑で,高等な類ほど主脈以外は単純化する傾向がある。2対の翅はときに前後形を異にし,バッタやカマキリでは前翅が硬化し,使用していないときは,その下にたたまれた膜質の後翅を覆っている。この傾向をおし進めたのが甲虫類で,その前翅は硬い革質の鞘翅(しようし)となって体を保護している。別の特化として双翅目では後翅を微小な平均棍(こん)に変えてしまった。この舵つき単葉機を開発した双翅類の中に,ホバリングなどの最高の航空技術を行いうるグループが含まれている。翅の出現は,明らかに昆虫類に画期的な成功をもたらした。しかし二つの点は指摘しておくべきであろう。第1にすべての昆虫が優秀な飛行技術者ではないということ。滞空性が生活時間のかなりを占めるのは,もっとも進化した完全変態類中の双翅目,鱗翅目,膜翅目と,もっとも原始的な有翅昆虫であるトンボ目,それに数百kmの飛行が可能な移住性のバッタくらいであろう。第2にすべての昆虫で翅は成虫,つまり最後の生活段階でのみ機能的となる。長い発育段階は翅の効用にあずかれないのであり,外骨格のための脱皮という宿命がここにも現れている。
次に体内構造について特徴的な点をあげる。節足動物の神経系は体節制を基本とする。各体節ごとに1対の神経球があり,これが左右前後に神経索で連結されたはしご状構造をとって,脊椎動物とは逆に腹面を縦走する。昆虫類もこの様式に従い,脳を別とすると神経球と主神経索は腹部にある。しかし左右の神経球はすべて癒着し,前後方向への癒着も著しい。頭部にある食道上・食道下神経球(前者が脳)は,それぞれ3対の神経球が癒着したものである。この集中化が,昆虫類の示す高度な諸行動の発現を可能にしたのであろう。消化管は前・中・後腸の3部からなり,前・後腸は外胚葉性で内壁はクチクラ層で覆われる。中腸は内胚葉性で,消化・吸収機能はおもにここで行われる。なお中・後腸の境界に,排出器官である何本かのマルピーギ管が開口している。
昆虫の体内器官系で脊椎動物と著しく異なるのは,循環・呼吸系の役割と配置が逆転している点であろう。昆虫の循環系は,背面を縦走する管状の背管(背脈管)のみであり,血管系はない。背管の後半,腹部にある部分は,いくつかの心室からなり心臓とも呼ばれる。前半(動脈)は単純な管にすぎず,胸部を貫き頭部内に開口する。心臓が接続する筋肉で動かされると,体液が各心室の心門から入り,前方に流れて頭部で体腔内に流出し,胸,腹を経てまた心室に入る。血液とリンパ液の区別はなく,体液(=血リンパ。一般には血液と称する)は直接体組織を洗っている。
一方,呼吸系は,基本的には胸部2対,腹部8対の気門から体内に入りこむ気管とその分肢からなる。気管は各気門からのものが相互に連絡して体内を縦走し,さらに分岐してその末端は毛細気管となり,あたかも脊椎動物の毛細血管のようにあらゆる組織内に入りこみ,そこでガス交換を行う。
以上述べた昆虫の特徴,効率的な体制をもち,ことに優秀な中枢と感覚・運動力を備えていることが,彼らを陸上で脊椎動物に匹敵する成功群に仕立てあげた。体が小さいことは,脊椎動物が利用できない微細環境を有効に用いるのに役だつであろう。しかし小型であることが彼らに有利に作用したとしても,なぜ大型になれなかったのかという疑問は残る。外骨格,脱皮の必然性,開放血管系で体液が体腔内を直接洗うことなどの彼らの体制そのものに,大型化を阻止する原因があるのかもしれない。
生き方(生活様式)は,どんな動物でも,どこにすむか(生息場所),何をどのように食べるか(食性),どうやって敵から,もしくは不適当な無機環境条件から身を守るか(防御習性および環境への抵抗性)などの個体維持習性と,どうやって子孫を残すか(生殖習性)という種族維持習性から成り立っている。最大の動物群である昆虫類では,これらの各習性も多岐にわたり,その組合せとしての生活様式も極度に変化しており概括しにくい。彼らの特性は彼らが採用していない生活様式で否定的に示すしかない。まず第1に,きわめて効率的な体制をもちながら脊椎動物のように大型化しなかったことが,個々の種類の微細環境への束縛をもたらし,これがおそらく種類数増加に一役買った。第2に獣や鳥のもつ恒温性をもてなかった(例外,ミツバチの集団)ため環境温度の変化から十分に独立することができず,温・寒帯では,生活を活動期と不活動期に分けざるを得ない。第3の特性として,生きものの故郷である海と,これほどはっきり絶縁した動物群も少ない。莫大な種類数の中で海産昆虫の占める比率はごく小さく,この点過半の種類が海産で,一部が淡水産,ごく少数のグループが陸産である甲殻類と対照的である。この陸上生活志向が,上述のような温度変化への適応を起こさせ,また海産動物に多い群体生活,定着生活,浮遊生活などの出現を不可能もしくは困難にしている。
以上の限定のもとに,昆虫の生活様式は多様をきわめるが,ここでは二つの特殊な生き方,集団生活と寄生生活にふれておく。いずれも昆虫類の専売特許ではないが,この類でとりわけ多様化している。集団性は三つの型に分けられる。(1)必ずしも血縁性のない個体の集合による一時性開放集団。これは交尾・休息・越冬・移住集団などに見られ,集団の統合度は弱い。集団の大きさはまちまちで,移住性バッタのような巨大集団ともなりうる。(2)幼虫集団。これは多くは同一卵塊から孵化(ふか)した血縁者集団だが,ときにいわゆる行列毛虫として知られる各種の鱗翅目幼虫やクロバネキノコバエ科の幼虫のように,融合によって巨大な二次集団に成長する場合もある。統合度は,前者と同等,またはより高い場合があり,しばしばメンバー間に同調行動が見られる。カレハガの1種では独立して食餌源に移動する幼虫と,他幼虫の吐いた糸のあとをたどってのみ移動する幼虫がまざっている。前者は食物発見にリーダーシップを果たすが,これのみで後者を欠くと,分散度が激しく,安定した吐糸巣がつくれない。(3)家族集団。上記の幼虫集団も一種の家族集団といえるが,ここでは成虫とその血縁の子の集団を指す。子が成虫となり親と共存した場合,子が不妊性となって集団の維持につくすと,カースト制が生じ,膜翅目とシロアリで著しく分化を遂げた社会性昆虫の出現を生み出した。
昆虫類は寄生生活の多様性においても,他の動物群をしのぐ。まず脊椎動物専門の寄生昆虫があり,これが寄生時期によって,成虫期のみ(ノミ,ナンキンムシなど),幼虫期のみ(ウマバエ,ウシバエ),全ステージ(シラミ,ハジラミ)に分けられる。次に無脊椎動物寄生の類がある。ほとんどが昆虫かクモを寄主とするが,双翅目には多足類,ミミズ,軟体動物に寄生するグループもいる。これらの類の寄生はすべて幼虫期に行われる(例外はネジレバネの雌,ミツバチシラミバエ)。昆虫寄生者も成虫の演ずる役割に応じて二つの型に分けられる。(1)成虫は寄主と関連ある場所に産卵する場合が多いが,寄主そのものには産まない。孵化直後の1齢幼虫は活発な探索行動を行う場合が多く,また寄主発見のチャンスの低さに応じて,産卵数が多いことが多い(ツチハンミョウ,カマキリモドキなど)。(2)成虫が寄主を探索し,その体表または体内,もしくは巣穴内に産卵する。一部の双翅目(寄生バエ)と多くの膜翅目(寄生バチ)の採用する方法で,捕食寄生性とも呼ばれる。この型の特性は寄生動物に多く出現する退化現象が,幼虫には現れても成虫には生じない点にある。膜翅目において寄生バチがしだいにカリウドバチに変わり,さらに社会性まで出現させた原因は,この捕食寄生性にある。最後に労働寄生があげられる。これは他種のなした労働(集めた食物やつくった巣など)を横取りする現象で,膜翅目で全動物中最高の発達を遂げた。寄生が昆虫の種類増加に大きな原因となっていることは,成功群である膜翅目の過半がこの生活を採用している点からも明らかであろう。
今まではどちらかといえば成虫を対象として述べてきたが,確かに分類はおもに成虫を材料として行われるし,体制も成虫でもっとも完成する。しかし昆虫の成虫期は個体の生活史の最後にくる比較的短い生殖期間であり,それ以前に長い摂食期間がある。多くの不完全変態昆虫では,この摂食期(仔虫(しちゆう)期)と成虫期は,後者で翅と生殖機能が完成する以外とくに差異がなく,個体の一生はしだいに成長はするがほぼ同じ体制と生活が保たれている場合が多い。一方,完全変態類では幼虫期と成虫期とは体制的にも生活様式でも別個の動物といってよいぐらい異なり,その間に位置する不活動期(さなぎ)に幼虫から成虫への劇的な体制の再編成が起こる。この現象(変態)が比較的遅く出現し,かつ現在成功しているグループに見られることは,この方法の採用が適応的意義をもつことを示唆する。ただし生活史の分化は必ずしも変態のみで起こるとは限らない。不完全変態をするトンボと,その水中生活を過ごす仔虫,同じくセミとその地中生活をする仔虫の生態的差異は,オサムシ類などの完全変態昆虫の幼虫と成虫の場合よりはるかに大きいといえる。昆虫の生活史は変態のほかにもいろいろな変化を示す。温・寒帯では生活史のある時期に不活動期(休眠,休止など)が挿入される。移住性バッタのように,過密状態になると通常の少食,非群居,定住的な孤独相から,移住性をもつ多食性の群集相に生活様式をスイッチさせる場合,アブラムシのように単為生殖世代と有性生殖世代を交互に繰り返す場合など,昆虫類の生活史の展開は多様をきわめる。
→変態
人類の起源をどの時点に置くかにかかわらず,人間の出現時すでにすべての現存昆虫群は現在とほぼ同じ体制をもって生活していた。初期の人類はそのあるものを食用とし,あるいは逆に彼らに吸血され,病気を伝搬させられたかもしれない。人類よりはるか以前からアリ,シロアリ,キクイムシは農耕(菌類)を,またアリは牧畜(アブラムシ,カイガラムシ)を開発した。人類がおくればせながら農業,牧畜を始めて以来,多くの昆虫類が害虫として登場したし,逆にミツバチ,カイコなどの利用も行われるようになった。彼らについては〈害虫〉〈益虫〉の項目にゆずるが,一言しておきたいのは,これらの用語がきわめて人間中心主義であること,および害虫の多くは人間がつくり出すことである。昆虫はそれ以外にも文学や芸術の中でさまざまな形でとり上げられたし,ホタルや鳴く虫は日本人の季節感と強く結びついてきた。今後も人間は害虫を駆除し益虫を増殖し続けるだろうし,それは必要なことではあろう。しかし地球規模で進行している環境破壊の結果,目につく虫といえばハエとゴキブリだけという状態は起こしたくないものである。
執筆者:坂上 昭一
最古の昆虫化石は,スコットランドのライニーRhynieのデボン紀中期の地層から発見されたリニエラRhyniellaという微小なトビムシ化石で,現生トビムシと比べても基本的な違いはないとされている。しかし現在の知見では,トビムシ類は一般の昆虫とかなり異なった体制をもち,有翅昆虫の先祖とは考えがたく,むしろイシノミやシミに近い無翅昆虫より有翅昆虫が生じたと考えられているが,その化石はまだ発見されていない。
有翅昆虫化石の出現は,古生代石炭紀中期(約3億2000万年前)で,原翅類Protoptera,古網翅類Palaeodictyoptera,疎翅類Megasecoptera,原トンボ類Protodonata,原直翅類Protorthopteraなどの絶滅した目のものばかりでなく,カゲロウ,トンボ,ゴキブリ,直翅類,半翅類中の同翅類,シリアゲムシ,ラクダムシなどの先祖型も出現している。二畳紀になると,さらにカワゲラ,チャタテムシ,甲虫などがこれにかわる。空中という新しい生活圏に進出した昆虫は,このように爆発的に発展分化したが,なかにはメガネウラのような巨大なものまで現れた。二畳紀末までに,石炭紀中に出現した原始的なグループのいくつかは絶滅している。
中生代に入って出現したのは異翅類,ハチ,ハエ,ナナフシなどの先祖型で,中生代の主要なハエ類はケバエ,キノコバエなどに近い長角類,ハチではキバチ,ハバチ類など広腰類であった。ジュラ紀にはトンボ(ムカシトンボはこのころ多い),直翅類,同翅類,甲虫,脈翅類(現在のヒメカゲロウに近いガのような広い翅や眼状紋をもったものもある)などが繁栄し,このころには現在の昆虫のグループの大半が出そろっている。中生代後期には顕花植物の出現発達に伴い鱗翅類も現れ,新生代の初期(始新世ころ)には,現在の昆虫の主要な科がひととおり見られる。バルト海沿岸の虫入りコハクは漸新世初期のものと考えられている。
古生代後期には今の日本の大半は海中にあり,陸上植物さえごくわずかしか発見されず,古生代の昆虫は見つかっていない。日本最古の昆虫化石は三畳紀後期のもので,山口県下で甲虫,ゴキブリ,同翅類,原直翅類などの化石が発見されている。それ以後の中生代や古第三紀の地層からも昆虫化石が少数ながら見つかっているが,その数が増えるのは新第三紀の中新世以降のことである。このころの化石昆虫は現生のものに近いので,昆虫の進化系統のうえからの興味は薄いが,現在の日本の昆虫相が形成された由来を知るうえで貴重な資料である。
執筆者:藤山 家徳
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
字通「昆」の項目を見る。
出典 平凡社「普及版 字通」普及版 字通について 情報
Sponserd by ![]()
…しかし,最近天然ピレスロイドをモデルとした合成開発研究が進み,ペルメトリン,シペルメトリンなどのNRDC系,フェンバレラートやエトフェンプロックスなどのきわめて殺虫力の高い薬剤が開発され,従来衛生害虫の防除にのみ用いられていたピレスロイドの農業面への利用の可能性が開かれた。 ジフルベンズロンやブプロフェジンなどの昆虫成長制御剤と呼ばれる一連の殺虫剤は,昆虫におけるキチン合成を阻害したり,幼若ホルモン活性を示し,その作用により昆虫の脱皮・変態が撹乱されて殺虫作用が発現する。この系列の薬剤には尿素構造を含むものが多い。…
…例えば動物は,迫ってくる危険を避けたり,種内の交信,雌雄の求愛,交尾などに聴覚を利用する。 聴覚が発達した動物の一つに昆虫が挙げられる。昆虫のうち,直翅(ちよくし)類のキリギリス,コオロギ,バッタやセミ類のように発音する昆虫では,同種個体間の交信に聴覚が用いられるが,ヤガやカゲロウなどでは,コウモリから逃れるためにコウモリの出す音を聞き分けている。…
…土中にトンネルを掘るモグラの足はパワーショベルのようであり,水中に餌を追い求めるアザラシやイルカの足は魚のひれに似ている。夜空を飛びまわって昆虫を捕らえるコウモリの羽が前足の変形であることはいうまでもない。このような形態変化の系列を眺めると,それぞれの動物の生きざまによく適した構造が,進化の過程で形成されたことを容易に想像できるであろう。…
※「昆虫」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
[名](スル)二つ以上のものが並び立つこと。「立候補者が―する」「―政権」[類語]両立・併存・同居・共存・並立・鼎立ていりつ...