動物の変態
生物の個体発生において,幼生での分化が緩やかになるか,または停止した後にくる成体への分化の過程。卵から孵化(ふか)した個体の基本構造が,性成熟の有無を除き,成体と同一の場合を直接発生というが,変態はこの直接発生に対置される概念ともいえる。変態は原生動物を除くほぼすべての動物門で知られているが,哺乳類をはじめとする胎生および卵胎生種,直接発生をする軟骨魚類や鳥類,爬虫類,クラゲや群体ボヤなどでみられるような無性生殖をする動物では変態は起こらないといえる。
変態過程を生活環に組み入れるか否かは,その種のとる繁殖戦略と密接なかかわりをもつ。この戦略は二つに大別される。一つは大卵方式で,胚は栄養蓄積の豊富な卵中で発生の全過程を終える直接発生をするか,あるいは親による保護下で胚発生あるいは後胚発生を終了する。いま一つは小卵方式で,多数の小卵を産するが,卵中の栄養物質は限られるため,産卵後すみやかに孵化し,自由生活者である幼生に移行する。このため,幼生は生命維持に必要な最小限の付属器官のみを備え,成体構造の構築はあとにまわす。このような種の孵化後の個体を幼生といい,多くの場合,幼生は成体と異なる生態的地位(生活空間,餌などが異なる)を占める。
水生無脊椎動物
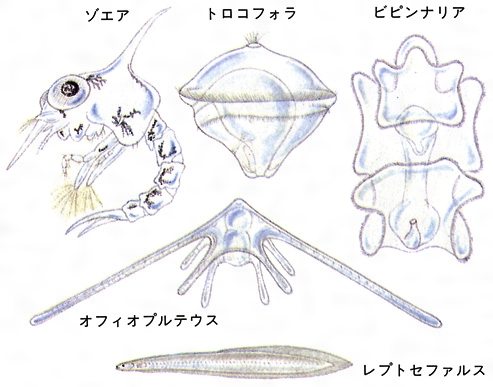
変態はほぼすべての海産無脊椎動物でみられる。これらの成体はカイ,ゴカイなどのように底生か,フジツボ,カイメン,ホヤなどのように固着生であることが多い。このため,胚と成体の間に幼生世代をはさみ,この世代に種の分散と成体の生活空間の選択という役割を担わせている。繁殖には小卵方式をとることが多く,幼生は自由浮遊生活者である。上記の目的のために幼生は特有な運動器官と摂食器官,成体の生活空間を選択するための感覚器官などを備えるが,これらの一部は成体では消失するか新たなものに置き換えられる。このような変態はふつう漸進的に生じ,成体の器官が成長するにつれて,幼生器官は退化する。例えば,軟体動物腹足類では体軸がねじれ,肛門が口の付近まで移動するが,幼生の構造の多くが成体でも保持される。環形動物のムカシゴカイでは,トロコフォラ幼生の下部に体幹部の原基を生じ,これが後方に増節しながら伸びて,成体の匍匐(ほふく)生活へと転向する。しかし,体制をまったく変えてしまうものも少なくない。棘皮(きよくひ)動物の幼生は左右対称であるが,成体は放射相称となる。ウニではプルテウス幼生の左脇の真皮上にウニ原基が生じ,ここから成体の構造が構築され,幼生のもつほとんどの構造は捨てられる。また,ホヤのオタマジャクシ幼生の変態は劇的な例として知られる。幼生の尾部は幼生が岩などの基盤に付着後,速いものでは数分以内に吸収され,続いて脊索,神経,筋肉などほとんどの幼生器官は退化してしまう。変態はしばしば脱皮を伴い,とくに節足動物では顕著で,甲殻類では脱皮ごとに体制が変化することもある。短尾目十脚類(カニ)では,ノープリウス→プロトゾエア(孵化)→ゾエア→メタゾエア→メガロパと異なる5段階の幼生をへて幼ガニとなるが,各段階のうちでも数回の脱皮を繰り返す。このことから,各幼生段階への移行を一つの変態としてとらえることもできる。
昆虫
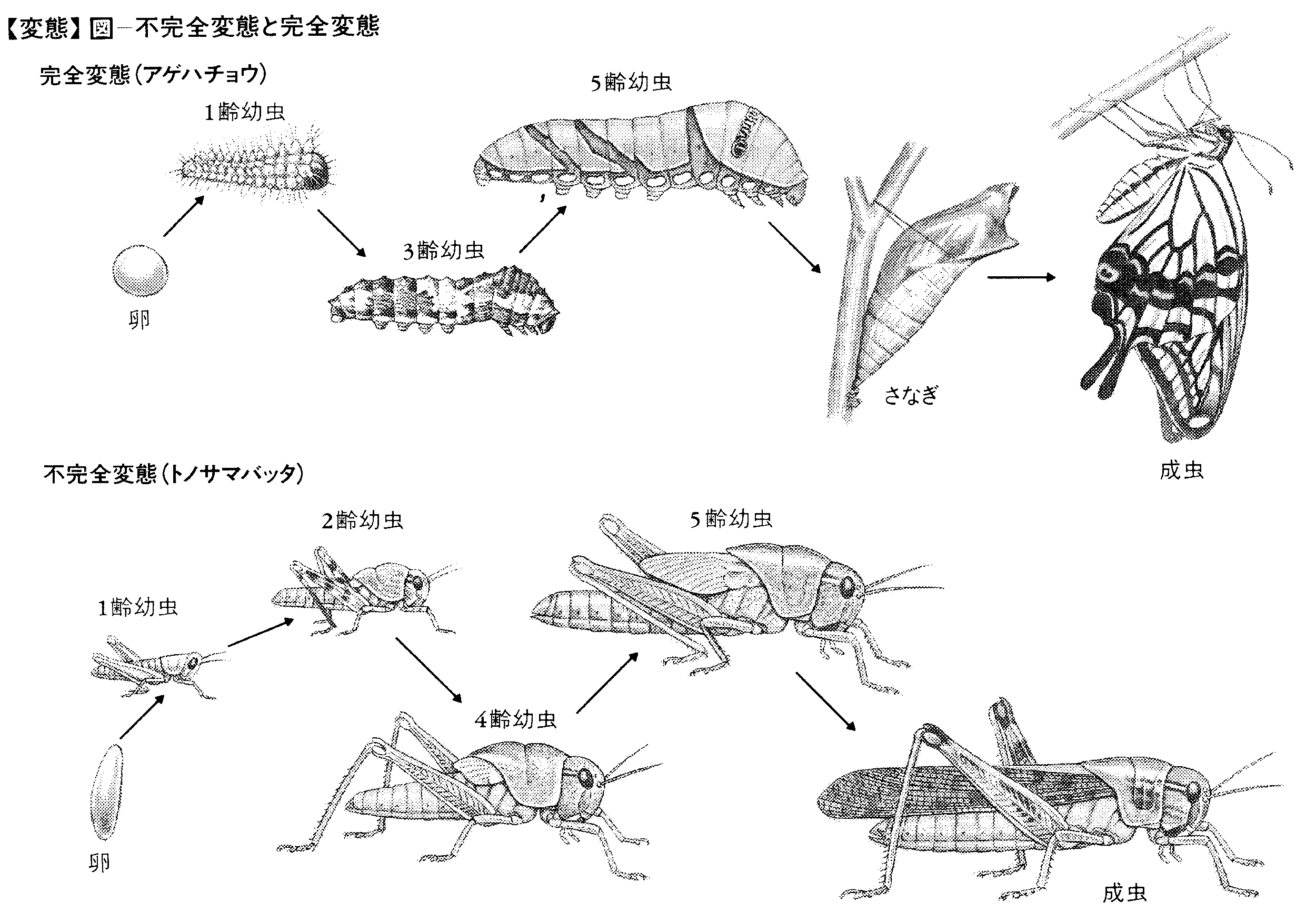
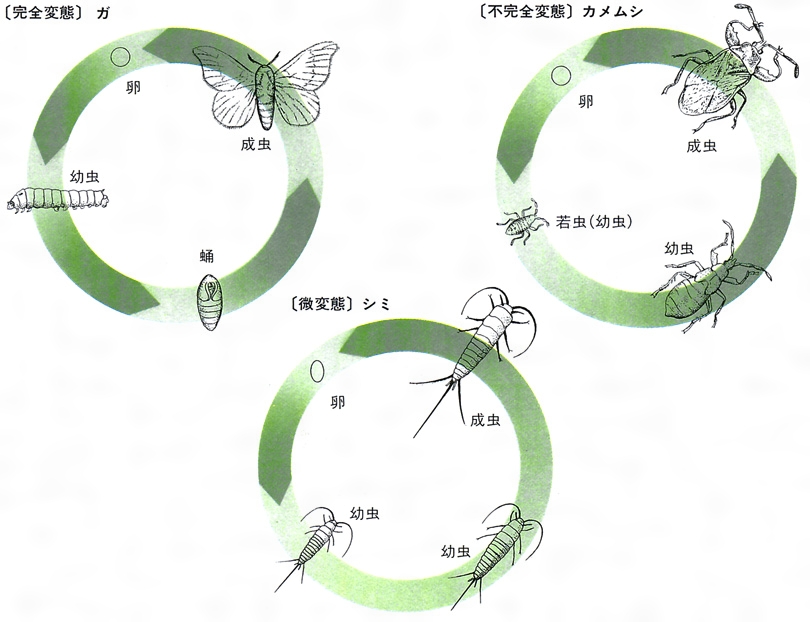
陸生無脊椎動物の変態は昆虫で詳しく調べられてきた。無翅(むし)類を昆虫に含めるか否かはさておき,昆虫の変態は便宜的に,無変態(不変態,上変態)ametaboly,不完全変態hemimetaboly,完全変態holometabolyに大別される。
(1)無変態 無変態昆虫には無翅亜綱のトビムシ目(トビムシ),原尾目(カマアシムシ),コムシ目(コムシ),シミ目(シミ)があり,外部生殖器以外は体形に変化なく成虫となる。無翅類原尾目は各脱皮ごとに腹部環節が1節ずつ増加するため,とくに増節変態ともいう。有翅昆虫は旧翅類と新翅類に大別され,後者のうちの少新翅類だけが完全変態をし,残りはすべて不完全変態である。
(2)不完全変態 不完全変態は蛹(よう)期のないことにより特徴づけられ,孵化後,翅や外部生殖器の原基が外部に現れている形式をさすが,変態の形式は多様である。幼虫と成虫の生態学的地位は同一であることが多いが,トンボやセミなどのようにまったく異なる場合もある。不完全変態は前変態,漸変態,新変態に大別される。前変態はカゲロウ目のみで,これは亜成虫から成虫へと成体で脱皮する唯一の例である。漸変態はさらに原変態と少変態に分けられ,そのうち原変態をするものにトンボ目,カワゲラ目がある。これらの幼虫は水生で,形態は成虫と著しく異なり,陸生の成体へと移行するため変態は激しい形態変化を伴う。少変態は典型的な形式の不完全変態で,ゴキブリ目,カマキリ目など10目以上にわたってみられる。新変態は半翅目同翅亜目とアザミウマ目でみられ擬蛹を形成する。特殊な変態型が多く,タマカイガラムシ類のように翅原基が最終齢で初めて外部に現れるものを同変態,アザミウマのように不動の半若虫から翅原基が外に現れる若虫をへて成虫になるものを再変態,摂食をしない若虫の齢期が成虫の直前にあるものを副変態(カイガラムシ科の雄),無翅の終齢幼虫から成虫が羽出するものを異変態(半翅目コナジラミ類)と称する。シロアリ目シロアリ科では,変態の様相は混乱をきわめ,Kalotermesでは,孵化後,脱皮を繰り返したのち擬働きアリ(幼虫)が生じ,これが脱皮して前兵アリから兵アリになる場合と,1齢若虫から2齢若虫をへて有翅の成虫になる場合があり,さらに2齢若虫が1齢若虫,1齢若虫が擬働きアリへと脱皮後,形態的に戻ることもあり,統一する変態の名称はない。
(3)完全変態 蛹期があることで特徴づけられる完全変態は,翅肢などの成虫付属器官が成虫原基(成虫芽)の形で幼虫期の間体内にある内翅類(鱗翅目,双翅目,甲虫目など)にみられる。この型の変態をする種のほとんどで,幼虫と成虫との生態的地位は異なる。また,幼虫から成虫への形態変化は劇的で,幼虫内部組織・器官の多くが消化され,新たに成虫のそれが形成される。幼虫にはシミ型(三爪(さんそう)幼虫を含む),芋虫型(多肢型),うじ虫型(無肢型)があるが,各幼虫期ごとに異なった型をとる種もかなりある。多変態をするシデムシでは1齢はシミ型,2~3齢は芋虫型である。ツチハンミョウでは1齢はシミ型,2~3齢は芋虫型,4齢は不動の囲蛹(3齢幼虫の皮膚が囲蛹殻となる),5齢でまた芋虫型となりさなぎをへて成虫となる。寄生性の膜翅目とネジレバネ目は過変態昆虫で,ネジレバネ類は1齢の三爪幼虫が寄主のスズメバチにとりつき,体内へ侵入したのち,うじ虫型のうじとなるし,ヤセバチ類では1齢はシミ型,2齢は芋虫型,3齢はうじ虫型幼虫となる。
脊椎動物
脊椎動物の尾索類(ホヤ,オタマボヤ,ウミタル)の有性生殖個体や頭索類(ナメクジウオ)で変態がみられる。脊椎動物では円口類(ヤツメウナギ),硬骨魚類(ウナギ,サバ,カレイなど),両生類に顕著な変態がみられるが,いずれも漸進的変化である。ウナギでは白色半透明のゼリー状の葉形幼生(レプトセファラス)から小型のシラスと呼ばれる稚魚期へ,ヤツメウナギではアンモシーテス幼生が孵化後数年たったのち数ヵ月で成体へと変態する。海産脊椎動物で顕著なのは,ヒラメやカレイで,両側にあった目は前頭部をまわって一側となり,無眼側の体色は白色となってしまう。ただし,有眼側が体の左右いずれにくるかは,種によって決まっているわけではない。
両生類無尾類(カエル)の幼生はオタマジャクシと呼ばれ,変態時には尾の吸収,四肢の出現,エラの退化と肺の形成,口器の変更,目の位置の移動などと体制は大幅に変化する。これらの変化に伴い,窒素代謝物の排出はアンモニアから尿素へと,ヘモグロビンは酸素親和性の高いものから低いものへと変化するように,生理的にも水中から陸上へ適応する移行がみられる。このような移行も変態を間にはさむことにより解決されている。
両生類の変態には二次的変態がある。イモリの1種の成体は,繁殖期になると一時的に水中生活に適応するようになり(water-drive),これはプロラクチン分泌によるとされている。ブチイモリ属のイモリは,水生の幼生は外部のえらと扁平な尾を備え,体色は緑であるが,第1次変態でオレンジ色の粗い皮膚の陸生イモリとなり,森林の林床にすむようになる。数年後,第2次変態を起こし,滑らかな皮膚をもつ水生の生活へと戻るが,尾には骨があり,外部のえらはない。同時に生理的変化も生じる。視物質は陸生特有のロドプシンrhodopsinから水生特有のポルフィロプシンporphyropsinへと変化し,窒素排出も尿素からアンモニアへと変化する。これらすべての変化はプロラクチン様物質でひき起こされる。
変態とホルモン
変態過程の引金は少数の例を除いてよく解明されていない。昆虫では,幼若ホルモンが減少あるいは消失したとき脱皮ホルモン(エクジソン)が作用すると変態が始まる。また両生類では腺下垂体(下垂体前葉)から分泌されるプロラクチンが減少し,同じく前葉から分泌される甲状腺刺激ホルモンが増加し,次いで甲状腺ホルモンが増加すると,変態が起きるとされている。
執筆者:桜井 勝
植物の変態
植物に変態という語を初めて用いたのはJ.W.vonゲーテで,彼は植物体のすべての部分は茎や葉から変形してきたものであると考えた。維管束植物の体制は,茎を基本とし,根と葉が茎から分化してきたものであることが明らかにされているが,そのように分化した基本的な器官である根,茎,葉が,普通の様態からはなはだしく変形したものになることを,現在では,変態という。根は通常植物体の支持と吸水の機能をもっているが,サツマイモの貯蔵根,ヒルギやアコウなどの気根や呼吸根,クモランの同化根などのほか,カワゴケソウの仲間のように根が植物体の基本となる場合もあり,根の変態の例とされる。葉からの変形としては,サルトリイバラの巻きひげ,サンショウモの沈水葉,タヌキモの捕虫葉,メギの葉針などが,また,茎からの変形には,カラタチのとげ,ジャガイモの貯蔵茎,サボテンの扁平になった葉のような茎,アリ植物にみられるアリの住居となる茎,ヒルガオのつる茎などがよく知られた例である。
変態は植物の形態が種によって定まっている変形であるが,種の属性をふみ外した変形もしばしばみられ,このような異常形は奇形malformationと呼ばれる。これは個体発生の異常などによって出現するもので,その性質が子孫に伝わることはない。
植物の種の進化はいろいろの表現形質が変形することによって認められるものであるが,その変形のうち,特定の形質が異常に変形してしまったものが変態であるといえる。ゲーテは植物の形態を一つの原型からの変態で説明しようとしたが,陸上植物の進化についてみると,根や葉は茎の変形として分化してきたものであり,花は胞子葉を中心とした苗条の変形したものであることが確かめられてきているので,ゲーテの思弁的な形態の解釈は,現代的な実証によってあらためて確認されているといえる。
執筆者:岩槻 邦男

 (せんくわい)して、憂ひを
(せんくわい)して、憂ひを (たの)しましめんとす 南人(苗族など)の變態を
(たの)しましめんとす 南人(苗族など)の變態を る
る