デジタル大辞泉 「人類」の意味・読み・例文・類語
じん‐るい【人類】
[類語]人間・人・ホモサピエンス・人倫・万物の霊長・考える
Sponsored by ![]()
Sponsored by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponsored by ![]()
現生のヒト(学名はホモ・サピエンスHomo sapiens)およびヒトに類似した特徴をもつ近縁な動物のグループをさし,その身体的特徴は直立二足歩行への適応を示す。分類学上の位置づけは,類人猿と人類に関する分類システムに諸説があるために一定しないが,最近は人類をヒト亜科としてまとめるのが一般的で,チンパンジー亜科とともに霊長目真猿亜目ヒト上科ヒト科を構成する。ホモ・サピエンス以外はすべて絶滅種で,古人類あるいは化石人類と呼ばれ,ネアンデルタール人(ホモ・ネアンデルタレンシスHomo neanderthalensis),ホモ・エレクトスHomo erectus,ホモ・ハビリスHomo habilisなどからなるホモ属Homoの他,アルディピテクス属Ardipithecus,ケニアントロプス属Kenyanthropus,アウストラロピテクス属Australopithecus,パラントロプス属Paranthropusなどを含む。また,サヘラントロプス属Sahelanthropusやオロリン属Orrorinも,直立二足歩行を示す特徴をもつことから,人類に含められる。ホモ属以外の化石人類は,すべてアフリカで発見されており,人類の進化の主な舞台がアフリカであったことは確実である。
現生人類を,形態学的特徴や遺伝的特徴に基づいて,アフリカ人,ヨーロッパ人,アジア人などの地域集団(人種)に分けるが,地域集団間の遺伝的特徴の差は肌の色など一部の特徴を除けば小さく,個人の遺伝的特徴から所属集団を確実に判定することは困難である。また現生人類を文化の差異によって分け,同じ文化を共有する人々をまとめて民族と呼ぶ。
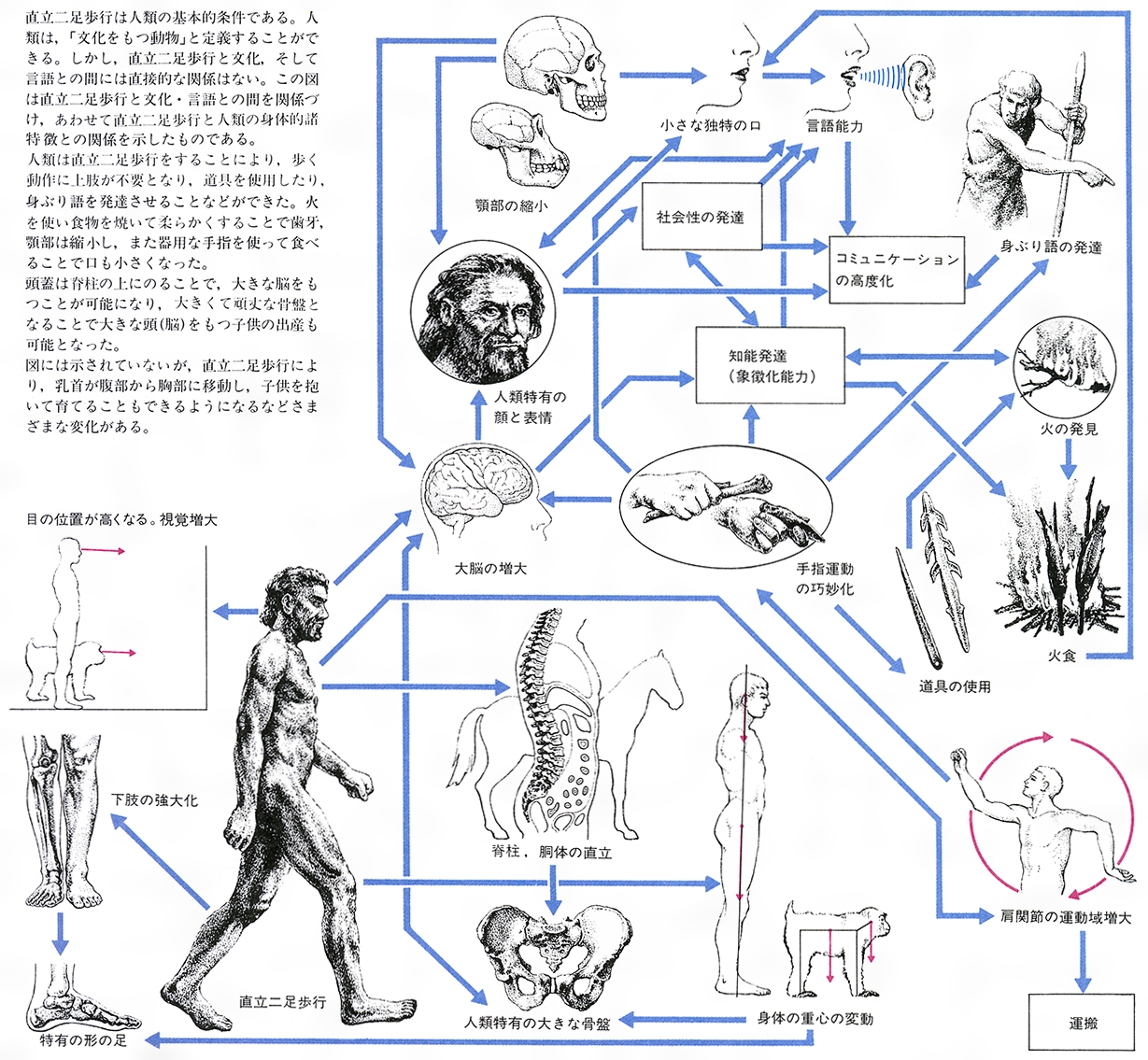
かつて人類の定義,すなわち他の霊長類と異なるヒトの特性として多くの項目があげられたが,近年の霊長類を対象とした研究によって,それらの中のいくつかは無効であることが明らかになった。狩猟もヒトだけの特性ではなかったし,近親婚の回避や,集団間での女性の交換といった項目も,人類にとって固有の特性ではないことが明らかになった。今日,人類学的に妥当と考えられる人類の定義としては,直立二足歩行,音声言語,人間家族の三つをあげることができる。このうち直立二足歩行は,脳の大型化などの他の特性に先行し,その意味で最も本質的で,化石霊長類の分類の際,人類すなわちヒト亜科に属するか否かの判定の決め手となっている。
野生チンパンジーも,木の枝や葉を加工したり石を組み合わせたりして簡単な道具を作り,シロアリ釣りや,水飲み,ナッツ割りなどを行うことが知られているが,狩猟採集生活を行う人類においては,道具製作が生存に必須の条件となった。そのような環境下で道具の使用・製作方法が改良を加えられながら代々受け継がれ,それを支える知的能力が発達を遂げたと考えられる。直接的で確実な証拠としては,道具の製作については250万年前のアウストラロピテクス・ガルヒによるオルドバイ石器が,また道具の使用については340万年前のアウストラロピテクス・アファレンシスによる石器の使用痕のある動物の骨が,現在知られている最古のものである。木や皮を使った道具も多く用いられたはずであるが,痕跡を残さない。
言語については,実験的研究によって,チンパンジーなどが手話を学習し,実験者と会話をする能力をもつことが明らかにされている。チンパンジーは,200以上の語彙を学習し,状況に応じた表現を自発的にかつ適切に生成し,そのいくつかを組み合わせて,より複雑な表現をすることもできる。また,扁(へん)と旁(つくり)からなる漢字と同様に,基本図形を組み合わせて作った人工的な文字を,諸種の概念のシンボルとして学習し,その配列によって実験者と会話をする実験によって,分節性や文法をもつ人間言語にきわめて類似したコミュニケーションの能力をもつことが明らかにされている。ただし,音声を言語的なコミュニケーションに用いさせる試みは失敗に終わっている。野生チンパンジーの相互間に,この種の身ぶりによる会話が交わされているという報告はないが,相手の物乞いに応じて食物などを分配する行動が認められている。
家族すなわち持続的な特定の男女とその間にうまれた子どもを含む小集団の存在は,人類のユニバーサル・カルチャーとされ,あらゆる民族に認められるが,霊長類社会についての研究によれば,ヒトを除くいずれの種も家族を形成するには至っていない。今西錦司は《人間家族の起原》(1961)の中で,人間家族の最小限度の条件として,インセスト・タブー(近親婚の禁忌),エクソガミー(外婚制),コミュニティ(地域社会),配偶者間のディビジョン・オブ・レーバー(分業)を挙げている。ここではヒト上科に属する現生の類人猿の社会について,人間の家族との相違点を検討しておこう。ヒト上科は,テナガザル科,オランウータン科,ヒト科に分けられ,ヒト科には人類すなわちヒト亜科とゴリラ,チンパンジー,ボノボ(ピグミーチンパンジー)からなるチンパンジー亜科が含まれる。テナガザル類はいずれも雌雄各1頭とその子どもからなるペア型の集団をもっている。オランウータンの社会は安定した集団をもたない単独生活者の社会である。ゴリラは単雄複雌の集団を基本としており,チンパンジーとボノボは複雄複雌の集団をもっている。類人猿に見られるこれらの集団は外婚の単位であり,集団内においては血縁関係が近い個体間の性的関係が回避されていることが知られている。すなわち,かつて人類に固有とされた外婚制やインセスト・タブーの基盤は,すでに高等霊長類の社会に胚胎していたといってよい。類人猿の集団間は敵対的ないし拮抗的で,集団をこえた地域社会は形成されていないが,複数の雄の協調によって支えられているチンパンジーやボノボの父系の集団は,狩猟採集民のバンドと共通する特徴をもち,その下位構造としての家族の形成を想定すれば,これを一つの地域社会と見なすことが可能である。しかし,父親の認知や性による経済的分業につながる要素は認められない。おそらく,家族の形成は言語や制度の発生とも関係しているのであろう。人類の社会は類人猿から受け継いだ父系の血縁的なきずなを基礎にしながら,家族という下位構造とともに,氏族,部族などの上位構造を構築してゆき,さらに地縁的なまとまりとしての民族,国家をつくりあげていった。このような発展が生業の発達と深い関係にあったことはいうまでもない。
約1万年前まで,人類のすべては狩猟採集を生業としていたと考えられている。したがって,ホミニゼーション(ヒト化)の舞台となったのは,狩猟採集という自然に依存し直接自然に働きかける生業形態であった。その間においても石器などの生活用具は絶えず洗練の度を加えていったが,全期間を通して本質的な生活の変化はなく,その社会は,今日の狩猟採集民のバンド社会と大きく異なるものではなかったと推定される。やがて,イネ科植物の群落が豊富な,そして有蹄(ゆうてい)類の群れが多く生息するサバンナ・ステップ地帯で,ある種の植物の栽培化や動物の牧畜化が始まる。このような生業の変革が,なぜこの時期に生じたのかは明らかではない。それを支えた精巧な石器類や火の使用,高度な知的能力などは,はるか以前から存在したからである。しかし,いずれにしても,農耕と遊牧は,いったん出現すると,人口の増加や階層化,富や知識の蓄積,集団間の競争などをもたらし,これによって人類の社会はより高度な組織と文化をもつ民族社会,古代国家へと,わずか数千年の間に驚異的な変化をとげることになったのである。
執筆者:伊谷 純一郎+多賀谷 昭
人類の進化は,系譜と特徴の両面から見ることができる。系譜すなわちヒトの系統に属する生物とその時間的配列は,ミトコンドリアDNAや核DNAなどの遺伝物質を用いた分子系統学的な方法と,化石を用いた形態学的な方法で研究される。分子系統学的方法は非常に強力であるが,遺伝物質は時とともに分解されるので,比較的新しい時代にしか適用できない。一方,ヒトの特徴の時代的変化の研究には,形態学的方法や,考古学的方法,現生霊長類の観察などの方法が用いられる。
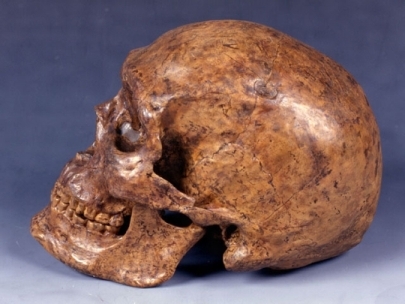
分子系統学によれば,現生霊長類のうち現生人類に最も近いのはチンパンジーやボノボのパン属Panであり,約600万年前に人類と共通の祖先をもつとされる。また,母系で伝わるミトコンドリアDNAの研究により,現生人類は約20万~15万年前にアフリカで出現し,その一部は約7万年前にアフリカを出てユーラシア大陸に広がったとされる。最初は東に向かってアジア方面に広がって,5万~4万年前にオーストラリアに達した。ヨーロッパへの進出はやや遅れ,約5万年前にメソポタミア地域を通ってヨーロッパに広がったと推定されている。ネアンデルタール人は,かつてヨーロッパの現生人類の直接の祖先と考えられたが,少なくとも35万年前に出現しており,ミトコンドリアDNAも現生人類とは別の系統として進化したことを示唆する。しかし,最近の核DNAの研究によれば,アフリカを出た現生人類はネアンデルタール人とわずかながら混血し,その結果,ヨーロッパ人やアジア人の遺伝子のうち1~4%程度はネアンデルタール人から受け継いだものであるという報告もある。
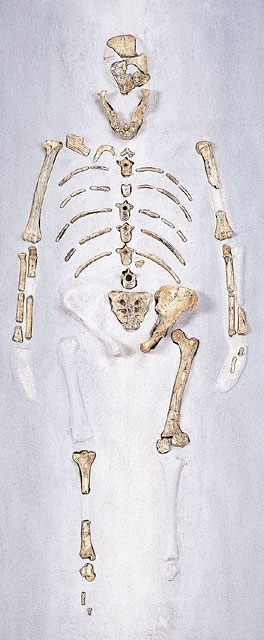
化石の骨や歯の研究によれば,700万年前のサヘラントロプス属Sahelanthropusや600万年前のオロリン属Orrorinが初期の人類の系統に属すると考えられる。ただし,年代からすると,ヒトとチンパンジーの分岐前である可能性もある。540万~440万年前に生きていたアルディピテクスは,ヒトとチンパンジーの共通祖先からわずかにヒトに向かって進化し始めた霊長類とされる。380万年前に出現したアウストラロピテクス・アファレンシスは,280万年前に出現したアウストラロピテクス・アフリカヌスや200万年前に出現したホモ・エレクトスの共通の祖先とされる。アウストラロピテクス・アフリカヌスから間もなくパラントロプスが派生し,ホモ・エレクトスからは,60万~35万年前のハイデルベルク人を経て,35万年前にネアンデルタール人が出現し,ヨーロッパから西アジア地域に4万年前まで生存した。現在,地球上にはホモ・サピエンス以外の人類は存在しないが,インドネシアのフローレス島には,わずか1万7000年前までホモ・エレクトスの仲間(ホモ・フローレシエンシス)が生存していたとされる。アルディピテクスの生息環境や身体的特徴から,直立二足歩行の進化の舞台は森林だったことが明らかになった。このため,以前に考えられていたような森林から開けた土地へ進出したことに伴う適応では,直立二足歩行の出現を説明することができず,チンパンジーとヒトの共通祖先が,ヒトほど効率よくではないにしても,直立二足歩行を移動様式のレパートリーの一つとしていた可能性も否定できなくなった。
人類をほかの霊長類から区別する第1の生物学的特徴は直立二足歩行であり,その特異な姿勢と移動様式に適応した形態は,主として腰から下にあらわれている。骨盤は短く広く,下肢は上肢より長い。大腿骨は広い骨盤の両側から斜め下方に向かうので,まっすぐ下方に向かう下腿骨と膝関節で鈍角をつくり,一直線上にない。足では親指と小指の中足骨が頑丈で,足底には縦と横のアーチが土踏まずを作り,足首の関節はしっかり固定されている。このような下肢骨の形態,構造は,それに関連する筋肉の発達とあいまって,長時間,直立姿勢を維持することを可能にし,歩行の際には,下肢を振り子のように使って足をぐらつかせることなく,滑らかに重心移動を行うエネルギー消費の少ない動きを可能にする。また脊柱がS字状に湾曲していることも,直立姿勢への適応と考えられる。このようなS字状の湾曲は,幼児が直立姿勢をとるようになると出現し,また,直立姿勢をとる猿回しのニホンザルにも出現することが知られている。顔面の前方への突出が弱まり,大後頭孔は頭蓋底の下方に向くよう前方に移動する。そして,頭蓋は脊柱の真上に位置するので,小さな力で支えることができ,頸の後の筋肉が付着する後頭骨の項平面は狭い。
人類の第2の特徴は,食性,ひいては咀嚼(そしやく)力の弱化に関連する。類人猿は基本的には植物食で,線維の多い大量の食物を咀嚼するために,歯や骨,咀嚼筋が発達しているが,人類はより効率的に栄養摂取できる肉食を併用し,また,食物を軟らかくする調理技術を身につけた結果,歯と顎骨はしだいに縮小して顔は垂直となり,下顎にはオトガイ(下顎の前端が歯より前に突出する状態)が形成された。咀嚼筋の弱化は,筋の付着部位を与える脳頭蓋,および咀嚼による応力を受けとめる力学的枠構造としての顔面頭蓋の形にも大きな影響を及ぼした。すなわち,脳頭蓋は頭蓋容積が増すにつれて横方向に拡大するとともに高くなり,顔面の骨格は頑丈性を失って縮小し,その結果,鼻が突出した。咀嚼とは直接関係しないが,犬歯の矮小化もヒトの大きな特徴で,類人猿に見られる巨大な犬歯,それを収容するための歯隙(しげき)や剪刀(せんとう)状の下顎第1小臼歯は姿を消し,歯列弓はU 字形から放物線状に変化した。
第3の特徴は道具製作の発達と関連するもので,脳の大型化,前頭葉の発達,手指とくに親指の精妙な動きなどである。これらの変化は,他の器官の変化と密接に関係している。腕渡りを行う類人猿では手の親指が短く,他の指は体重を支える強力な把握力を発揮できる構造になっており,道具の製作や使用に適した手の構造は,直立二足歩行を前提にしている。また,道具使用が犬歯や咀嚼器官の退化をもたらしたことは明らかである。さらに,道具の製作や使用は,個体の能力だけでなく,改良を加えながら伝統として受け継がれる必要があり,そのためにコミュニケーションに関係する発声器官や,豊かな表情を可能にする顔の構造,視線を明示する白目の露出,それらを制御し意味を解読する大脳などの発達を促すことになったと考えられる。
以上述べたような形態特徴は,人類の進化において徐々に発達したもので,ヒト亜科全体にあてはまるわけではなく,現生人類では明瞭であるが,古くさかのぼるほど薄れてゆく。ただし,その出現時期や発達の速度は,特徴によって異なる。たとえば,200万年前のアウストラロピテクスの歯や下肢は人類の特徴を強く示すが,その脳容積は小さく,チンパンジーとあまり違いがない。また,ネアンデルタール人の脳容積は現代人のそれよりむしろ大きいが,明瞭なおとがいの形成には至っていない。人類の系統をさかのぼれば,その特徴はヒト亜科とチンパンジー亜科の共通祖先がもっていた特徴に近づくはずである。それは,全体としてみればパン属,とくに特殊化の弱いボノボなどに近いと思われるが,共通祖先から分かれた後,ヒト亜科だけでなくチンパンジー亜科も特殊化を遂げているので,共通祖先の身体特徴には,類人猿よりもむしろヒトに近いものや,どちらにも似ていないものが存在する可能性がある。実際,アルディピテクスの骨格は,大型類人猿のようなナックル歩行ではなく,直立二足歩行と四足歩行の併用に適応した構造であったとされる。
→ホミニゼーション
執筆者:多賀谷 昭+池田 次郎
現生人類や古人骨由来のDNAの分析が進んだことで,現在ではDNA情報をもとにした人類の起源と拡散に関するシナリオが構築されている。現生人類がもつ遺伝的な多様性はアフリカ人が一番大きいことから,現世人類はアフリカで誕生したと推察されている。また塩基置換速度の推定値を元にした計算によって,その時期は20万~15万年前と考えられており,さらにアフリカ人以外の現生人類は7万~6万年前に出アフリカを成し遂げたと考えられている。現在,最も詳細に拡散の状況を描くことができるのは,ミトコンドリアDNAの系統データである。現生人類のミトコンドリアDNAには,ハプログループと呼ばれる世界拡散の過程で突然変異によって生み出された様々な系統が存在し,その分布と相互の関係は,人類の移動ルートを推定する材料となっている。
人類が世界拡散を成し遂げた時期は,最終氷期に相当し,海水面は現在よりも大きく低下していた。出アフリカのルートには定説はないが,DNA研究からは現在のエチオピア,ソマリア周辺から紅海を越えてアラビア半島に渡った経路が想定されている。海水面の低下がそれを助けたことは間違いない。
アフリカ以外では南アジアの遺伝的多様性が大きいことから,この地域が旧大陸における拡散のセンターになったと考えられている。考古学的な証拠から,オーストラリア大陸には4万7000年以上前に現生人類が到達したことが明らかとなっており,同じ時期に東南アジアから東アジアへの展開も成し遂げられたと考えられている。なお,ヨーロッパ人の遺伝的な多様性は他の地域に比べると小さく,この地域への進出は,さらに遅れたと想像されている。
およそ2万年前には,人類は当時陸続きだったベーリング海峡(ベーリンジア)を通過して新大陸に到達したことが,アメリカ大陸先住民のミトコンドリアDNA研究から判明している。その後の展開は急速で,考古学的な証拠からは数千年で南米大陸の先端にまで達したと考えられている。こうして1万年ほど前までには現生人類は,ほぼ地球上の大部分の地域に居住することとなった。
最後に残されたのは,南太平洋の島々であったが,この地域への入植は6000年ほど前に,中国大陸の南部もしくは台湾からの農耕民が移住することで成し遂げられた。3000年ほど前にメラネシアから船出した集団が,約千年間をかけてポリネシアの島々に展開することになったことが,メラネシアやポリネシアの集団の考古学や言語学,そしてDNA研究から明らかとなっている。
→人類学
執筆者:篠田 謙一
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponsored by ![]()
ヒト、人間ともいうが、用い方に若干のニュアンスの差がある。分類学的には霊長目のヒト(類人類)上科のヒト科に属する動物。われわれ自身と同類の総称であるが、歴史的にみれば、人類という概念の範囲はかなり恣意(しい)的、便宜的である。超過疎であった採集狩猟時代では、自部族中心の狭い世界の者のみを人類とみなしていた形跡がある。コロンブスによる「アメリカ大陸発見」当時、征服者たちは先住民をキリストの福音(ふくいん)を受けない者として、人類とみなさず、殺戮劫略(さつりくごうりゃく)を繰り返した。今日では、形態・行動上、多くの差異がありながら、地球上各地の人類はすべて単一種であるという一般的認識があり、それは同一世界に生きているという連帯感の基礎となっている。人類にもっとも近縁の種は類人猿各種であり、これらとは同じヒト上科をつくるが、形態・行動の点で両者の差は歴然としている。一般に人類は動物界のなかでもきわめて独自のものとされているが、共通する点も多々あり、とくに化石段階の初期人類は類人猿と区別しにくい形態をもつ。今日、人類学者たちの共通の理解では、人類である基本的条件は直立二足歩行(姿勢)であるが、他方「文化をもつ動物」として人類を規定することもできる。
[香原志勢]
直立姿勢の採用は、手足、すなわち上下肢の分化と同時進行した。胴体を直立させての前進姿勢は、他の動物にはみられず、力学的にも不安定である。このため骨盤は著しく幅広く、強大化し、上半身を担う。下肢は上肢より長く、膝(ひざ)が伸び、大腿(だいたい)も長くなる。多くの哺乳(ほにゅう)類は指先、つまさきのみを地につけて歩くが、人類はかかとを含め足底をつけて歩く。歩行速度は遅いが、二本足で歩くには安定する。足自体は前後に長く、土ふまずがみられるが、これは足部内に骨からなる縦・横のアーチがあることを示す。このアーチがばねとなり、歩行を軽やかにする。脊柱(せきちゅう)はS字状に彎曲(わんきょく)し、歩行時の衝撃が脳に直接伝わることを防ぐ。胸部横断面は前後径より横幅のほうが広い。肩は側方に張る。頭蓋(とうがい)は脊柱の上にのるため、他の動物と異なり、大後頭孔(だいこうとうこう)は頭蓋底の中央部にあって下方に向かい開口する。一般の動物の体幹は平生はほぼ水平であり、その前端が顔となるが、直立姿勢をとる人類の体幹は垂直であり、そのほぼ上端の前面に顔がくる。四足動物の内臓諸器官は前後に並ぶが、直立した人類の内臓は上下に重積するため、胃下垂のような構造的な疾患が生じる。遊走腎(じん)、めまい、痔疾(じしつ)、腰痛(ようつう)、背曲がりなどの障害も、直立姿勢に対する不適応現象といえる。
[香原志勢]
人類は霊長類の一員である。霊長類は全体として特殊化の程度が弱いが、樹上生活に対して適応し、人類の身体特徴もこれに準じる。母指(おやゆび)が他の4本の指と向かい合う母指対向性は、サルでは手足にみられ、枝を握るのに適している。人類の足ではそれは消失するが、手の母指は発達し、強大になり、またほかの指も自在に動き、道具扱いに適する。サルも人類も手足の指先に指紋をもち、爪(つめ)は鉤(かぎ)状でなく、扁(ひら)爪である。両眼は顔の前面に並ぶため、立体視が可能となり、距離の目測に都合がよい。一般哺乳類と異なり、人類とサルの嗅覚(きゅうかく)は鈍いが、色覚に優れる。森林にすむ霊長類の聴覚は格別優れてはいないが、人類の耳介(じかい)は皿状で、また動かない。高等猿類の表情は、知能が高いこともあって動きが活発であるが、人類のそれは一段と発達し、複雑かつ微妙に動く。類人猿も人類も外見上、尾はない。人類の身体は諸関節の可動範囲が広いため、肢体がよく屈伸し、さまざまな姿勢、手ぶりを示すことができるが、それは樹上生活をするサルの特質を受け継いだものである。
[香原志勢]
人類の特色として、犬歯がほかの歯とほぼ同じ程度に短小化し、その先端が丸くなる。それは牙(きば)ではなくなり、その他の歯、とくに大臼歯(きゅうし)も小さくなり、咬面(こうめん)も単純化し、歯列がU字形から放物線状に変じ、上下顎骨(がくこつ)は縮小し、結果として、突顎から直顎へ移行する。そしゃく器の退縮は人類の重要な特徴である。それに伴って外鼻は顔面中央部に突出した。人類の最主要な特色と多くの人がみなす大きな脳は、霊長類の進化を引き継ぐものであり、初期人類の脳の大きさはゴリラ程度で、現生人類の約3分の1にすぎない。人類化への過程においては、大きな脳より、直立が先行する。しかし、比較的短い人類進化の過程に脳が3倍に増えたことは大きな意味をもつ。とくに大脳半球のうち、抽象的思考と関連ある前頭葉は著しく増大したとみられる。このような脳の拡大は、手指をはじめとする運動機能や視覚を中心とする感覚機能の発達、人類独自の言語能力の獲得、精神活動の活発化などによると考えられるが、一方、そしゃく器の退縮に伴い、そしゃく時の頭骨全体への衝撃が小さくなり、そのことが脳の増大に資したとみられている。結果として、大きい脳頭蓋と小さい顔面頭蓋という組合せを生み、額が形成される。
[香原志勢]
いつの時代に始まったか検証できないが、人類の体毛は非常に少ない。人類は熱帯の日光を浴びつつ、日中活動するが、一方、発達した汗腺(かんせん)を十分に機能させるためには、体毛の少ないほうが有利である。また顔毛の少ないのは、顔の表情をよく表す。頭髪が密なうえ、脱毛しにくいのは、発達した頭部の保護に適する。人体に分布する汗腺の多くは臭(にお)いの弱い汗を分泌するエクリン腺であり、一般哺乳類に普遍的な、臭いの強い汗を分泌するアポクリン腺は腋窩(えきか)、陰部などに限られる。その他の人類の特色として、生殖行動が季節性をもたず、年中可能なこと、長い受胎期間、きわめて大きな脳をもちながら未熟な新生児、大きな脳をもつ子を分娩(ぶんべん)しうる母体の広い産道と大きい骨盤、多くの場合単胎であること、胸部上方にある1対の乳房、哺乳・育児期間が長いこと、生殖開始期が遅いこと、生殖能力喪失後も生存し続けること、そして、ほかの動物と比べても長命なことなど、多くの点で人類独特のものがある。
[香原志勢]
人類進化は霊長類の進化を引き継ぐ。かつては人類の時代をもって第四紀とよんだが、地質学上の定義が変わり、また初期人類の実態が明らかになるにつれ、人類の発祥は第三紀鮮新世末にさかのぼることになった。人類の直接的祖先としてこれまで多くの化石類人猿の名があがった。歯の退縮を根拠に1970年代前後にはラマピテクスが有力視されたが、やがて別の道を歩むシバピテクスとの類縁性が指摘され、消えた。今日人類の直系祖先の候補者の座を占める化石類人猿はいないが、より古い人類化石の探索が続けられた結果、初期人類化石はすべて東アフリカから出土しており、同地こそ人類発祥の地とする見方に異論はない。以後時代が下るにつれその分布域は広がり、人類進化も進行した。その進化過程はおおむね連続的で複雑だが、全体的理解の便のため、猿人→原人→旧人→新人という4段階に分けられる。猿人は440万年前のアルディピテクス以外は、アウストラロピテクス類である。古いものからあげると、後者にはA. アナメンシス、A. アファレンシス、きゃしゃなA. アフリカヌス、そしていずれも頑丈なA. エジプティクス、A. ロブストゥス、A. ボイセイが入る。これらは400万年前から100万年前にアフリカに存在した。猿人と原人の橋渡しをするのが、ホモ・ハビリスで、とくに大型のものは最近はホモ・ルドルフェンシスとよばれる。240万~170万年前東アフリカに存在した。原人はジャワ原人や北京(ペキン)原人を含めて、ホモ・エレクトゥスで統一される。180万~30万年前アフロ・ユーラシアの熱帯、温帯に広く分布した。なお古い東アフリカのものをホモ・エルガステル、新しいヨーロッパのものをホモ・ハイデルベルゲンシスとよぶこともある。旧人はネアンデルタール人および古代型サピエンスという。15万~3万5000年前に旧大陸に広く分布し、氷期には氷河周辺にからくも居住するものがいた。新人は3万年前から今日に至るまでの人類で、ヨーロッパのクロマニョン人、中国の山頂洞人、日本の縄文時代人、そして現代人などはこれに属する。今日では極度に過酷な気候で不毛な土地を除き、汎(はん)地球的に分布する。旧人と新人はホモ・サピエンスのなかの亜種関係とみられている。進化段階が進むにあたり、しだいに、そしゃく器である顔面が退縮すると同時に、脳は大きくなる。原人の脳の大きさは猿人のそれの約2倍、旧人と新人は約3倍となる。多地域にもわたったこれらの諸人類の系統関係には諸説があり、小異を捨てて単系進化をしたとみる説がある一方、複雑な系統関係で想定する説もあり、定説はない。しかし、人類進化の流れは確実に定まっている。そのことは、19世紀には人類進化説が否定、もしくは無視されていただけに、思想界としてはきわめて重要な意味をもつ。
[香原志勢]
人類進化の事実は、単なる身体の形や動きなど生物学的叙述だけでは説明できない。文化の発達は生物学的現象ではないが、人類進化を引き継いでいるようにみえる。また人類進化を促進したのが文化である。それとしてまずあげられるのが道具の使用・発明である。一般の動物は身体を用いて生存環境に適応するが、人類は自分の身体以外のもの、すなわち道具、あるいは石や木などを用いて事態を処理して適応し、結果として進化した。その際、人類の手の意義はきわめて大きい。道具の使用および火の利用はそしゃく器の退縮を大いに促した。また言語の使用は、自分の思いを容易に他者に伝達することにより、社会的紐帯(ちゅうたい)を強め、個人の知識を社会の知識にまで広げた。この言語の成立は優れた知能によるところが大きいが、一方、口、咽頭(いんとう)、喉頭(こうとう)が優れた発声器官として副次機能を完成させたことも銘記すべきである。そのほか、人類の身体の多くの部分が、文化を受け入れ、発展させるのに適していたことも見逃せない。人類進化に伴って、その文化は採集狩猟、農耕牧畜を経て工業・商業へと発展し、現代人と現代文化をつくるに至った。
[香原志勢]
人類は生物学的にはホモ・サピエンスという種に属している。人類の祖先を昔にさかのぼっていくと、猿の祖先との区別が困難になる。人類と猿は同じ祖先から進化してきたということは学界で一般に受け入れられている。そこで人類と他の動物との違いはどこにあるかという問題は、形質人類学の問題であるとともに、文化人類学の問題でもある。人類は社会的動物であるという説は、アリやミツバチのように、社会をつくる動物は人間以外にもあるところから、部分的な真理しか含まないことがわかる。結局のところ、人類と動物を区別する根本的な特色は言語の使用という点にあるという説に疑問を挟む者はいない。
言語能力は、レズリー・ホワイトによると、人類のみが所有するものである。これは、ホワイトによると「象徴化の機能」である。いいかえれば、あるものに意味を与える能力である。犬が「おあずけ」や「お手」を覚えるように、動物でも条件づけによって、音声の意味をある程度習得することができるが、動物は、あるものに意味を与えること、つまり「象徴化」の能力をもっていないのである。この人類に特有の「象徴化」の能力こそ言語をつくりだし、文化を生み出したというのである。
それでは、人類は形質的に象徴化の能力をまず十分に備えたうえで、言語や文化をつくりだしたのだろうか。最古の化石人類「猿人」は直立歩行しており、カモシカやクジラの骨から道具をつくり、荒削りの石器をつくっていたところからすると、素朴な原初的な文化をもっていたと思われる。文化人類学者クリフォード・ギアツが、「アウストラロピテクス(猿人)が、いまの人間のわずか3分の1の大きさの頭脳しかもたないのに、文化の原初形態をもっていたとすれば、人間の大脳皮質が拡大したのは、文化の開始に先行したのではなく、それに後続しておこった」のであろうと述べているように、人類の象徴化の能力は、文化の発生前につくられたのではなく、文化の発展に伴って育成されてきたと考えるほうが適当であろう。形質人類学者ウォッシュバーンも、「解剖学的にわれわれと等しいからだをもった人間が、ゆっくりと文化を発見していったと考えるよりも、われわれの(身体的)構造の多くは、文化が存在したことの結果であると考えるほうがおそらく正しい」といっている。
現在地球上に住むあらゆる人々は、前述のように一つの「種」に属しているが、人類のなかの人種の違いが遺伝的な優劣の違いを現すのではないかという誤った考え方がしばしばみられるし、西欧民族がもっとも優れているという偏見や、他民族を蔑視(べっし)する傾向も古くからある。とくに19世紀の西欧において、西欧文明を頂点とし、西欧以外の文化をすべてこれより劣ったものとみる見解は学問的に体系化され、文化は直線的に進化するという文化進化主義が登場した。明治以来、日本が西欧の諸科学を導入する過程において、意識的あるいは無意識的に、西欧の直線的文化進化論も受け入れてきた。
ところが、研究が進むにつれて、いわゆる未開民族の文化もそれなりに発展してきていることがわかり、しかも文化はかならずしも一直線には進化しないことも実証されるようになった。自分の国の価値判断の物差しで他の民族の文化を計ることを避け、正邪・善悪の規準はそれぞれの文化に応じて異なるという文化相対主義が主張されるようになった。確かに技術の面ではわれわれのほうがいわゆる未開人よりも優れているが、風俗習慣について優劣を決めることは困難であるだけでなく、不適切であることがわかってきた。
人類は、形質的に、白人種群、黄色人種群、黒人種群に大きく分類されるが、人種間に先天的、遺伝的な知能差が存在することを示すデータは何もないのである。誤った人種観、人種主義racismが歴史上数々の悲惨なできごとを生んできたことを忘れてはならない。
人種という概念は純粋に生物学的概念であって、風俗習慣の違いやもっと広く文化の違いは、人種の違いとは直接関係がない。同じ人種の人々も異なる文化に成長すれば、まったく異なる文化を担うようになり、異なる人種の者でも同じ文化に育てば、同じ文化をもつようになる。人類の文化は多様で、地球上の民族や国民のもつ個別的文化は、マードックによれば少なくとも3000以上ある。文化は多様であるが、その反面に類似性も存在する。そのなかには、人類のもつ共通の生物学的条件という要因を無視することはできない。また、西欧の科学技術文明に染まらない人類普遍の「人間精神」、思考様式の探究がフランスの構造人類学によって推進されている。人類の文化人類学的研究は、文化の多様性とともに文化の類似性にも注目すべきであろう。
[吉田禎吾]
『人類学講座編纂委員会編『人類学講座』全13巻・別巻3(1977~2001・雄山閣)』▽『祖父江孝男著『文化人類学入門』(中公新書)』▽『吉田禎吾著『未開民族を探る』(社会思想社・現代教養文庫)』▽『山口昌男著『文化人類学への招待』(岩波新書)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponsored by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponsored by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponsored by ![]()
字通「人」の項目を見る。
出典 平凡社「普及版 字通」普及版 字通について 情報
Sponsored by ![]()
…一方で,移動運動において上肢をまったく使用しない直立二足歩行を行うグループが出現した。これが人類である。 漸新‐中新世類人猿の果実食性は,歯の形態にも影響をあたえた。…
※「人類」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponsored by ![]()
政府首脳が外国を訪問した際の会談内容や合意事項を記した外交文書。法的拘束力は持たないが,その内容は両国を事実上拘束する。類似のものに共同発表 joint statementがあるが,これはより記録的な...
7/28 化学辞典 第2版(森北出版)を更新
6/26 日本大百科全書(ニッポニカ)を更新
4/17 デジタル大辞泉プラスを更新
4/17 デジタル大辞泉を更新
2/17 日本大百科全書(ニッポニカ)を更新