生物が外界から食物としてとり入れたタンパク質,脂肪,炭水化物などの高分子を,吸収可能な低分子にまで分解する過程を消化という。
消化研究の歴史
生体外での酵素反応である発酵現象は,早くから発酵素fermentの概念を生んだ。消化も同じく酵素反応だが,生体内での反応なので,むしろ精気の転成という見方でとらえられた。17世紀にJ.B.vanヘルモントは発酵素の概念を初めて消化に持ち込み,6段階の消化を区別した(1615)。第1と第2は胃と腸での消化であり,第3の消化では粗い血液が生成される。第4と第5の消化では血液が精気へと精練される。第6は体内の多くの〈胃〉による消化で,〈それぞれの場所に固有の発酵素が自己の食物を消化する〉。最後のものは,現代でいえば組織での代謝全般であり,上記の第1と第2が今日の消化,第3~第5は吸収と同化にあたる。F.シルビウスは唾液の役割に注目し,また化学の目で消化を見ることを強調した。その門からでたR.deグラーフは,膵液の意義を論じた(1664)。ブルンナーJohann Conrad Brunner(1653-1727)はイヌの膵臓摘出実験を行い(1682),また十二指腸腺を発見した(1687)。こうして消化の化学的研究が深まった。同時にこのころから消化の物理(力学)的な側面も取り上げられるようになり,G.A.ボレリは,鳥の胃が砂を含んでいてひき臼の作用をもち,ガラス球や木片も破砕することを述べた。しかし彼は,消化液の作用も否定したわけではない。R.A.F.deレオミュールは金属製の籠に肉片や骨片をいれてトビに飲みこませ,破砕作用がなくても消化が起こることを実証した(1752)。大実験家L.スパランツァーニはこの方法を多種類の動物に広げ,自分自身も木製の小管を飲んで実験した。さらに動物にカイメンを飲ませて胃液を得て,生体外で消化を観察した。19世紀には化学的研究がさらに進み,T.シュワンは胃の可溶性酵素をペプシンと名付けた(1836)。やがて唾液のジアスターゼや膵液のトリプシンも発見,命名された。食物成分の分解と吸収に関しても,たとえば脂肪は分解されて,古くから知られていた乳糜(にゆうび)管を通って吸収されるなど,具体的な知識が蓄積されていき,20世紀の研究につながっていったのである。
執筆者:長野 敬
消化の生物学的意義
自由生活を営むすべての動物には消化機能があるが,内部寄生性の動物の多くは消化機能がなく,宿主が消化した物質を体表から吸収する。また植物でも,食虫植物は捕らえた昆虫類を消化する酵素を備えている。
大きな塊の食物をとっている動物の場合,消化の第1段階はそれを砕いて嚥下(えんげ)しやすく,消化液に触れる面積を広くすることである(物理的消化)。ついで消化酵素による加水分解が起こる(化学的消化)。この酵素的消化過程が細胞内で行われるのを細胞内消化intracellular digestion,細胞外で行われるのを細胞外消化extracellular digestionという。細胞内消化は明らかに原始的なものであり,動物の進化とともに細胞外消化に移行してきたと考えられる。
原生動物やカイメン類はもっぱら細胞内消化を行う。腔腸動物になってはじめて細胞外消化がみられ,これによってある程度処理されたものが細胞内で消化される。扁形動物には種によって,細胞外消化のみを行うものと細胞内消化のみを行うものがある。ヒモムシ類,ゴカイ類および軟体動物は発達した消化管をもっていて,本質的には細胞外消化であるが,中には消化管壁の遊走性アメーバ細胞amoebocyteが細胞内消化をしてこれを補っている場合もある。頭足類,甲殻類,昆虫類,センチュウ類,脊椎動物では細胞内消化はほとんどみられない。
細胞内消化が行われるためには,食物がはじめから細胞内にとり込むことができるほど微細であるか,または,あらかじめ細胞外消化をして,そのような大きさにしておくことが必要である。食細胞作用や飲細胞作用(エンドサイトーシス)で細胞内にとり込まれた食物の消化にはリソソームが重要な役割を果たしている。リソソームは細胞質内にある小胞状の細胞器官で,ホスファターゼ,ペプチダーゼ,グリコシダーゼなど多種類の酸性消化酵素を備えている。食物を含んだ液胞(食胞)がこれと融合し,消化が行われると考えられている。細胞外消化は一般に消化管の前部で行われる(これを消化管内消化という)が,動物によっては口から消化液を出し,あらかじめ,ある程度消化したものをとり入れるという方法をとるものもある(体外消化)。たとえば,クモやアリジゴクは餌動物にプロテアーゼをしみ込ませて半消化したものを吸い込む。ヒトデは消化管を体外に出して餌動物を包み込んで消化するので,体外ではあっても消化管内消化である。
消化酵素はすべて加水分解酵素であるが,いろいろな種類があって,個々の動物のもっている消化酵素の構成は種特異的であり,食物の性質や摂食法および動物の進化過程と大きなかかわりがある。ある物質の食物としての価値は,動物の消化能力に依存する。たとえば,一般に哺乳類の子どもは,乳汁に含まれる乳糖をブドウ糖とガラクトースに分解するラクターゼをもっているが,他の脊椎動物にはない。また,乳糖その他の糖類を含まない乳を出すアザラシ,アシカなどの海獣も,この酵素をもっていない。昆虫はブドウ糖とガラクトースを栄養として利用できるが,乳糖を利用できないことが実験的に知られている。これはラクターゼを欠いているからである。また菌類を食べている昆虫の腸内には,菌類に多く含まれている二糖類のトレハロースをブドウ糖に加水分解するトレハラーゼがある。高級脂肪酸と高級アルコールのエステルである蠟は,リパーゼでは加水分解できないから,普通動物にとって栄養的価値のないものであるが,ハチノスツヅリガの仲間の幼虫は蜜蠟だけで育つ。南アフリカに住むミツオシエというキツツキ類の鳥は,蜜蠟を好んで食べている。後者の場合は消化管内に共生する細菌の働きで消化している。
一般に植食動物は,肉食動物にくらべて,はるかに多種類の炭水化物分解酵素をもっており,そのうえ,自身では消化できないものでも,消化管内に共生している細菌や原生動物の分解作用によって生じた産物を利用している場合が多い。これを共生消化という。草食獣とくに反芻(はんすう)類やシロアリなどにおけるセルロースの利用は有名な例である。
→共生栄養
執筆者:佃 弘子
ヒトにおける消化
消化とは加水分解反応にほかならないが,消化管内にはそれぞれの高分子物質を逐次分解していく消化酵素が消化腺から分泌されたり,小腸の細胞の膜に備わっていてその反応を触媒している。三大栄養素のうち,炭水化物およびタンパク質の消化と,脂質の消化にはやや異なった様相がみられる。前者の消化は管内の中間消化(管腔内消化)と小腸細胞による終末消化の二つの段階に大別できる。管腔内消化は,食物が分泌された消化液と混和し,消化液中に含まれている消化酵素の作用で行われる消化で,管内消化ともよばれるが,この過程は高分子物質から一挙に直接吸収しうる低分子物質を産生するのではなく,高分子内部の結合を切断して逐次分子を細かくし,吸収しうる低分子の一歩手前の段階までの消化をうけもつものである。したがって中間消化とよばれる。消化の第2段階はこのようにして産生された中間消化産物が,小腸の吸収上皮細胞の管腔に面した側の細胞膜表面または細胞の中で加水分解される過程で,糖は単糖類,タンパク質はアミノ酸にまで分解される。この過程を終末消化とよぶ。吸収細胞の管腔側は特殊な構造になっており,直径約0.1μm,長さ約1μmの細胞質突起(微絨毛microvillus)が整然と密生している。この微絨毛間の間隙(かんげき)は幅0.1μmとひじょうにせまく,細菌その他の微生物のはいり込めない空間となっている。この微絨毛表面の細胞膜には構成タンパク質の一部として多糖類分解酵素(グルコアミラーゼ),各種の二糖類加水分解酵素,アミノペプチダーゼその他の酵素が存在している。糖質やペプチドの最終的な消化はここで行われ,この特殊な空間に出た最終消化産物は同じ細胞膜に備わった,濃度こう配に逆らって行われる強力な能動輸送によって,速やかに細胞内にとり込まれる。
糖質の消化
食物中の炭水化物の大部分はデンプンであるが,デンプンにはブドウ糖がα-1,4グルコシド結合のみで多数重合した直鎖構造のアミロースと,α-1,4グルコシド結合のほかに数%の割でα-1,6グルコシド結合を含む樹枝状構造のアミロペクチンの2種が混在する。唾液中のアミラーゼ(プチアリン)はα型であり,α-1,4グルコシド結合を加水分解して低分子のデキストリンを産生し,最終的にはマルトース(麦芽糖)にまで分解する。α-アミラーゼはα-1,6グルコシド結合には作用しないため,アミロペクチンの場合はグルコースがα-1,6グルコシド結合で2分子結合したイソマルトースが数%産生される。唾液腺アミラーゼの至適pHは6~7であり,胃の上部に食物と唾液の混和したものが停留している間,消化は進行する(これを胃内消化という)が,胃酸と混和するとその活性は失われる。食物が胃から十二指腸に送られると,膵臓から分泌されるα-アミラーゼ(アミロプシン)により再び消化は進行する。膵液アミラーゼは,唾液アミラーゼと類似の性質をもっている。産生されたマルトース,イソマルトースまたはそれよりやや大きいオリゴ糖は,次いで終末消化の段階にはいる。オリゴ糖は微絨毛膜のグルコアミラーゼ(γ-アミラーゼともいう)の作用をうけ,分子末端からブドウ糖,マルトースを遊離させ,二糖類はやはり膜に存在する二糖類加水分解酵素(マルターゼ,イソマルターゼ,スクラーゼ,ラクターゼ,トレハラーゼ)によってそれぞれ単糖に分解される。これらの酵素は生理的条件下では,膜から離れて管腔内液に溶出することはなく,基質が膜の酵素に接触,結合して反応が進行する。このような消化を膜消化という。
タンパク質の消化
成人の1日当りの食品タンパク摂取量は約50~80gであるが,実際に腸管で吸収されるタンパク質の量は,このほかに消化液に分泌されるタンパク質および脱落上皮細胞に由来するタンパク質が同程度ある。これらのタンパク質も,管内消化(中間消化)と膜消化および細胞内消化の2段階の消化によって,最終的にはアミノ酸の形で血中に吸収される。タンパク質の管内消化に関与する酵素として,胃底腺の主細胞から分泌されるペプシンがある。ペプシンの至適pHは約2で,変性したタンパク質にのみ作用し,芳香族アミノ酸のアミノ基が関与しているペプチド結合を選択的に切り離す。膵液は多量の炭酸水素イオンを含み,胃より送り出された内容物の中性化に働くが,そうなるとペプシンの作用は消失する。小腸管腔内では,膵液中のタンパク質分解酵素であるトリプシン,キモトリプシン,カルボキシペプチダーゼが働くが,これら酵素もそれぞれ特異的に作用するペプチド結合がきまっており,それらの総合的な作用によって,小腸管腔内では各種の段階のオリゴペプチドならびにアミノ酸が産生される。通常小腸内では,ペプチドのほうがアミノ酸よりずっと多い状態になっており,そのような状態で吸収が進行している。オリゴペプチド,とくにやや大きいペプチドは,細胞の管腔側膜に存在するアミノペプチダーゼにより,N末端から加水分解され,産生されたアミノ酸は,膜輸送担体を介して吸収される。ペプチドの場合は,この膜消化のほかに,ジペプチドおよびトリペプチドを,このままの形で細胞内にとり込む担体が発達しており,かなりの部分のペプチドは,この輸送担体によって細胞内にとり込まれ,細胞内に豊富に存在するペプチダーゼによって,速やかにアミノ酸に加水分解される。この過程を細胞内消化という。
脂質の消化
食物中の脂肪の大部分は中性脂肪(トリグリセリド)であるが,これは胆汁酸の存在下,腸管運動のかくはん混和作用により,乳化された状態で膵リパーゼの加水分解作用をうけ,モノグリセリドと脂肪酸に加水分解される。脂肪の加水分解を触媒するリパーゼは,脂肪と水の界面で作用するので脂肪が乳化され,反応表面積が増加していると加水分解は著しく促進される。リパーゼの至適pHは約7である。脂肪消化産物は胆汁酸とともに疎水基を内に,親水基を外に向けた分子会合ミセルを形成するようになる。ミセルは水溶液中で安定であるため,疎水性の脂質消化産物もこの形になると,吸収上皮細胞膜までの水層拡散が容易となる。なお,炭素数が6~10のいわゆる中鎖脂肪の場合の消化は上記と異なる。この場合は,リパーゼにより脂肪酸とグリセリンに加水分解され,加水分解産物はミセルの形にならなくとも,吸収上皮細胞膜への拡散が比較的容易である。中鎖脂肪は,肝胆道疾患など胆汁酸分泌障害の患者などに医療として用いられるが,必須脂肪酸は含まれない。
→吸収
執筆者:星 猛+日向 正義
消化管運動
口腔内にとり込まれた食物は,消化管を口側から肛門側に移動しながら,消化液と混和して消化,吸収を受け,ついには大便となって体外に排出される。消化管内容の前進と混和は,消化管壁の筋の収縮によって行われるが,このような消化管の運動を消化管運動という。前進速度は部位によって異なり,胃腸にいたる単なる通路である咽頭→食道下端部間では速く,胃→直腸間では一般に遅い。混和により胃と小腸では,消化液中の消化酵素が有効に作用し,小腸と大腸では,粘膜との接触が盛んに行われて吸収が促進される。
ヒトの消化管壁筋組織は,口側端(口腔から上部食道まで)と肛門側端(外肛門括約筋)に横紋筋があるほかは,すべて平滑筋である。平滑筋層は外側の縦走筋層,内側の輪走筋層,粘膜と粘膜下組織の間の粘膜筋板より成る。胃幽門部や回盲部などの特定の部位では,輪走筋や縦走筋がとくによく発達し,消化管の通路を開閉する関所のような役割を果たして,内容の前進速度を調節している。このような部位を括約部という。消化管平滑筋は交感神経(食道を除く)と副交感神経の二重支配を受けている。平滑筋の収縮は前者により抑制され,後者により促進されることが多い。これらの神経(外来神経)は消化管壁内神経叢(筋層間神経叢と粘膜下神経叢)に連絡する。消化筋の運動は,いくつかの反射によって調節されている。たとえば,摂食後には,急に強い大腸の運動が起こるが,これを胃-大腸反射gastrocolic reflexという。また嘔吐時には小腸運動が抑制されるが,これは胃-小腸抑制反射gastroenteric reflexのためである。また消化管ホルモンも消化管運動の調節にあずかる。たとえば,ガストリンは胃運動の促進,幽門括約部の弛緩,食道下部括約部の収縮,小腸運動と胆囊収縮の促進をひき起こし,セクレチンは食道下部括約部,胃,小腸の運動を抑制する。
消化管運動には大別して,蠕動(ぜんどう)と分節運動の二つの型式がある。(1)蠕動peristalsis 消化管の一部で内外両筋層が収縮し,この収縮が口側から肛門側に波として伝わっていく。この収縮波によって消化管はしごかれ,内容は前進する。蠕動は食道以下のあらゆる部分でみられ,消化管内容輸送の原動力である。小腸と大腸では内容を一掃するような強い蠕動の起こることもある。十二指腸起始部や盲腸では逆方向の収縮波(逆蠕動)も認められる。逆蠕動は内容の混和に役だつ。その他の部位で逆蠕動が起こるのはおおむね通過障害のある場合である。(2)分節運動segmentation contractions 小腸と大腸に見られる。まず腸管のある部位に輪走筋層の収縮によるくびれが数ヵ所に生じ,その収縮がしだいに強くなったのち,くびれの部分は弛緩し,くびれとくびれの中間部に新しいくびれを生じ,ついで弛緩する。これが繰り返し起こる。この運動は腸管内容をかくはんし,こねまわし,消化と吸収を促進する。しかし,内容の前進には関与しない。分節運動は摂食により著しく盛んになり,絶食していると少なくなる。
執筆者:東 健彦
消化器官digestive organ
食物のとり込み,貯留,消化,吸収を行う器官をいう。脊椎動物の消化器官は,原腸が口陥および総排出腔陥で胚体外に開口してできた消化管と,その付属器官とよりなる。消化管は発生的に体腔外の頭腸(口腔,鰓域(さいいき))と体腔内の胴腸(前腸,中腸,終腸)に区分される。口腔は食物のとり込み,咀嚼(そしやく),嚥下,味覚の場である。鰓域は魚類の呼吸作用に関連し,肺呼吸動物では咽頭となる。前腸(食道,胃)は食物の輸送と一時的貯留および限られた消化・吸収作用をもつ。中腸(小腸)は消化,吸収の主要な場である。終腸(大腸)はおもに食物残渣(ざんさ)の処理,排出の場であるが,草食性哺乳類の一部では腸内細菌の関与する消化,吸収の場となる。
口腔の付属器官には,舌,歯,口腔腺がある。舌は捕食に関係した器官で,種属により変化があり,両生類(無尾類),爬虫類(有鱗類)などで発達し,鳥類,哺乳類でとくに大型である。有鱗類の発達した舌は鋤鼻(じよび)器官に連動した嗅覚作用をもつ。味覚は舌の重要な機能ではないが,哺乳類の筋質の舌には多数の味蕾(みらい)が乳頭内に見られる。歯は口腔の表皮に由来する硬質構造で,採食のため発達したが,四肢動物では一般に円錐形の同型歯で歯根がなく,かつ交換を繰り返す多代性歯である。哺乳類の異型歯はその多様な食性の変化に対応したもので,1回しか交換しない二代性歯となったのは,哺乳の習性の発達に関連したものと考えられる。
口腔腺は粘液を分泌して食物をしめらせ,飲み込みやすくする。普通小型の腺が口蓋や舌で見られるが,哺乳類では大型の複合腺である大口腔腺(耳下腺,下顎腺,単孔舌下腺,多孔舌下腺,頰骨(きようこつ)腺,眼窩(がんか)下腺など)が見られる。耳下腺のみが漿液(しようえき)腺でアミラーゼを分泌するが,その消化上の寄与はヒトにおけるほど重要ではない。反芻類やハムスターなどでは,唾液は体内窒素リサイクルの重要な径路となっている。ヘビの毒腺は口腔腺の変化したものである。なお,口唇は哺乳類特有のもので,哺乳のため発達したものである。
肝臓は中腸起部腹側から出芽形成された大型消化腺で,胆汁を胆管により小腸内に分泌する。また特殊な腸静脈(門脈)により,吸収された栄養分に富んだ血液を受け入れて代謝,調節を行う。例外としてヤツメウナギの肝臓は腸と連絡がなく,血液循環に関係するのみである。肝臓は中腸の背・腹側から出る3個の原基に由来し,肺魚類や多くの硬骨魚類ではまとまった腺体をつくらない。鳥類では4葉に分かれる。哺乳類でもウサギなどのように分散するものもあるが,一般には一つの腺体にまとまる。膵臓の外分泌細胞は強力な消化酵素を合成し,膵管(1~3本)により腸内に分泌する。内分泌細胞の大半は集合してランゲルハンス島を形成する。同種類の内分泌細胞は胃腺および腸,とくに十二指腸に多く存在し,消化機能の調節を行っている。
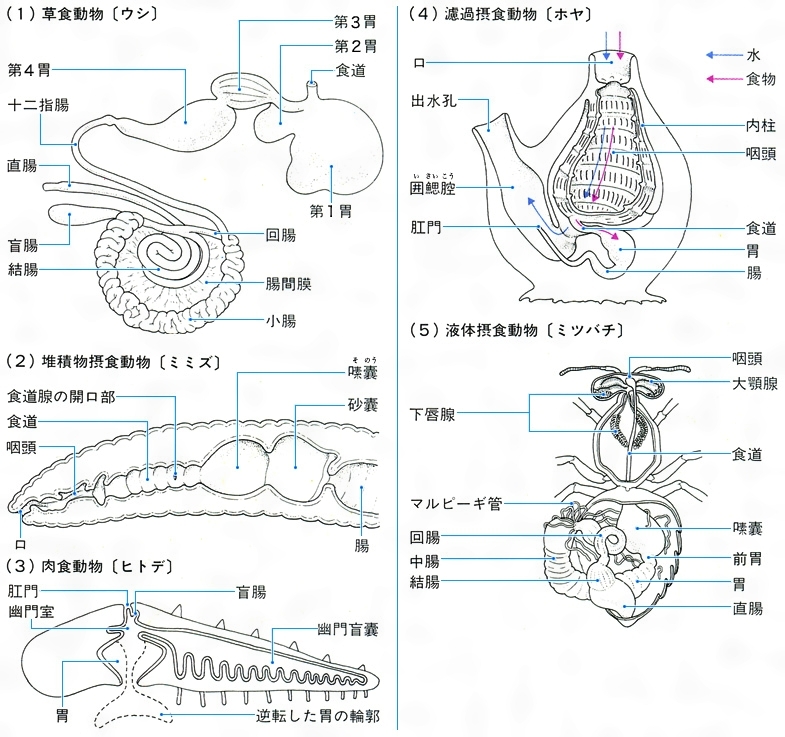
執筆者:玉手 英夫 無脊椎動物では,大多数の動物がもつ消化管の形態的,機能的に分化した各部域や,腔腸動物の胃腔が消化器官にあたる。消化管の口に近い部域である口腔,咽頭,食道は,主として食物の摂取,破砕,貯蔵の役割を果たし,歯,とげ,貯囊などを発達させていることが多い。これにつづく胃,腸では,付属する消化腺や盲囊が発達し,主として消化,吸収を行う。消化器官とそれに付属する消化腺などの付属器官をあわせて,消化器官系とするが,腔腸動物,環形動物,触手動物などの触手,ひも形動物の吻(ふん),軟体動物の腕,節足動物の口器付属肢,毛顎動物の顎棘(がつきよく)などの摂食・捕食器官は,一般に消化器官系には含められない。
執筆者:原田 英司