デジタル大辞泉 「代謝」の意味・読み・例文・類語
たい‐しゃ【代謝】
1 古いものと新しいものとが次々と入れ替わること。「新陳
2 生体内で、物質が次々と化学的に変化して入れ替わること、また、それに伴ってエネルギーが出入りすること。
[類語]交代・入れ替わり・入れ替え・
Sponserd by ![]()
翻訳|metabolism
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
代謝ということばは正確には新陳代謝の略語である。地球上の各種生物が外界との密接なかかわりをもちつつ,しかも自己の生命を維持するために,必要なさまざまな活動を推進するための最も基本になる活動が代謝にほかならない。代謝には,エネルギー代謝,物質代謝(物質交代)という二つの用語に示されるように,エネルギーの獲得,利用と物質の変換が不可欠な活動である。言い換えると,代謝とは酵素の触媒作用に助けられて,生物の体内で絶えまなく営まれている各種の化学反応の総称ともいえる。外界の環境条件,例えば温度,圧力,水素イオン濃度(pH),また種々の物質の濃度などは時々刻々変化している。一方ではまた,生物が日常摂取する食物の内容,種類,組成も変化する。それにもかかわらず,生物の体内の物質の組成や濃度,あるいはpHや体温などはほぼ一定に保たれている。このように,外界への適応性と自己の恒常性という,一見相反する特性を兼ね備えていることからもわかるように,生物は決して閉ざされた系ではなく,外界との疎通を積極的にはかる,むしろ開かれた系としての特徴を備えている。外界のある物質の濃度が,生体内における濃度よりも低い場合であっても,その生物がもっと要求する場合には濃度こう配に逆らってでも摂取する,いわゆる能動輸送の現象などがその典型的な例といえよう。動的平衡ということばで生物の特徴を表す人もある。
代謝の営みは内容の上で次の四つに大別される。(1)太陽や食物から化学エネルギーを獲得する営み,(2)外界から摂取した栄養素を,生体構造構築の素材や生体高分子合成のための素材(前駆体)に転換させる営み,(3)上記の前駆体から,核酸,タンパク質,脂質,多糖などの各種生体成分を合成する営み,(4)ホルモン,ビタミン,成長促進因子など,生物が必要とする各種の生理活性物質を合成し分解する営み。
生物の種の違い,生育環境の違い,生活様式の違いなどで,代謝の様式もかなり大幅に変化する。以下にその一部について考察してみよう。
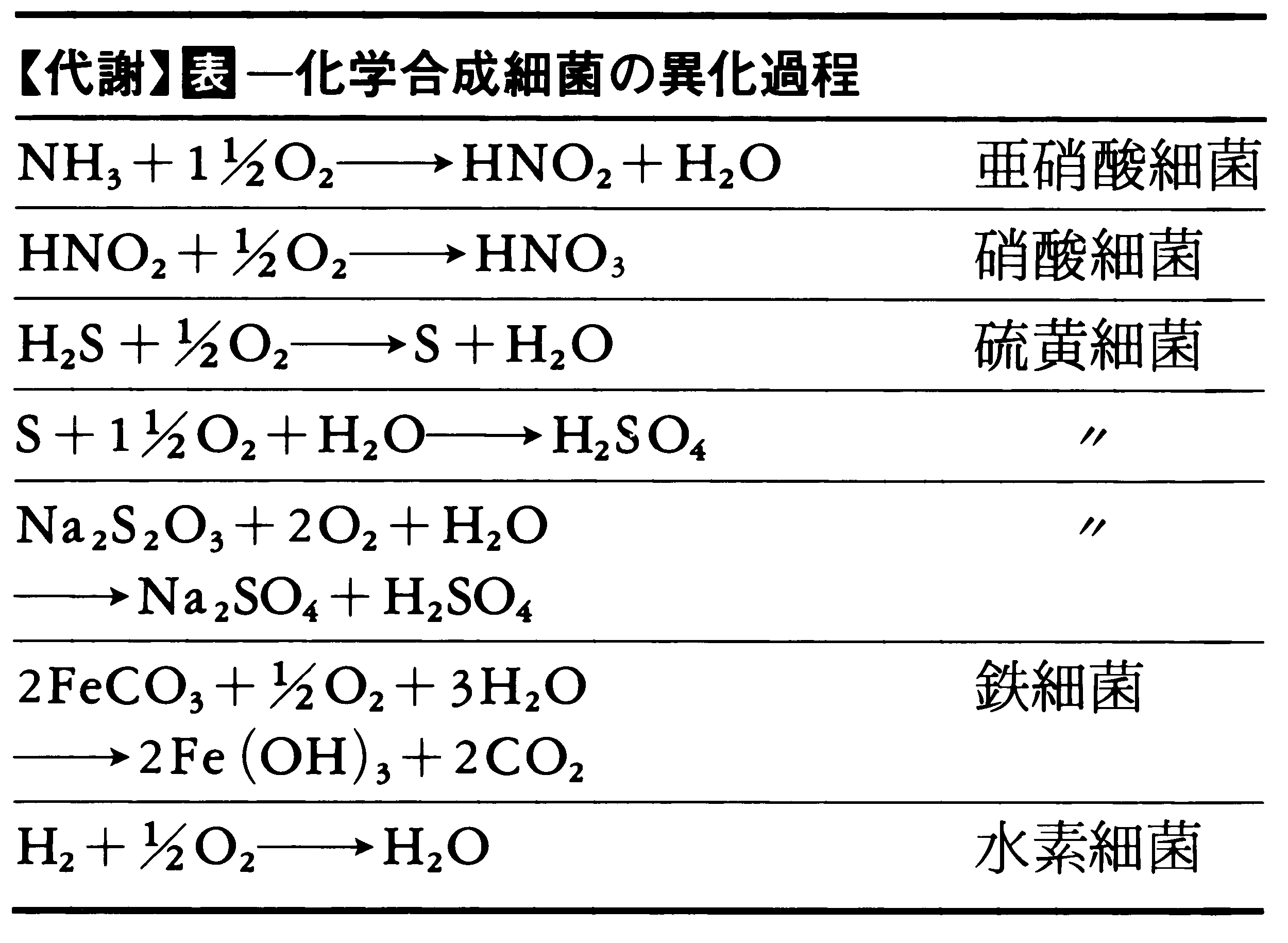
(1)エネルギー源の違い すべての生物のエネルギー供給源はもとをたどれば太陽にいきつくが,太陽エネルギーの利用変換の方式は,生物によって異なる。光を直接エネルギー源として利用できる生物は光合成生物phototrophと呼ばれ,これに対して酸化還元の化学反応を介してエネルギーを獲得する生物を化学合成生物chemotrophと呼ぶ。化学合成生物の中で,グルコースなどの有機物質を電子供与体とする生物を化学合成有機酸化生物chemoorganotroph,また硫黄,アンモニア,水素などの無機物質を利用するものを化学合成無機酸化生物chemolithotrophと呼ぶ。土壌中と海洋に生息する微生物の数は,地球上の全生物の過半数におよぶが,それらの多くは分子状窒素N2の固定やアンモニアの酸化による硝酸の生成などの化学反応によってエネルギーを獲得している。その一部を表に示した。
(2)炭素源の違い 生物が大気中の二酸化炭素CO2を唯一の炭素源として利用し,生体内のすべての有機化合物の炭素骨格を二酸化炭素から合成することを独立栄養,あるいは自養autotrophと呼ぶ。これに対してグルコースなどの,より複雑な還元型の有機化合物を利用しなければならない場合を,従属栄養あるいは他養heterotrophと呼ぶ。光合成生物やその他の限られたある種の細菌が独立栄養性であるのに対し,すべての高等動物と大部分の微生物は従属栄養性である。
(3)酸素を必要とする生物と必要としない生物の違い 生物の多くは,含水炭素(炭水化物)や脂肪を酸化することによって,その分解の過程で遊離するエネルギーを利用するが,このようないわゆる生体酸化は,酸素の存在下に進行する場合もあれば,深海や土壌中のような酸素分圧のきわめて低い環境下では,脱水素反応として営まれる場合も少なくない。われわれ人間の体内でも,心臓や肺のように酸素の供給が活発な器官と,筋肉のようにむしろ嫌気条件に近い部位とでは,代謝の方式も大きく変わってくる。従属栄養生物の中には,酸素を積極的に利用する好気性生物と,酸素があっては生育できない嫌気性生物がある。これらに加えて,好気・嫌気両方の環境で生育できる通性嫌気性生物facultative organismも存在している。嫌気性生物を好気条件下に移すとたちまち死滅するが,その原因の一つとして,カタラーゼが備わっていないために,発生した過酸化水素を処理できず,中毒死してしまうという解釈もある。酸素を利用できる生物にとっては,好気性代謝のほうが嫌気性代謝を営む場合よりもはるかに効率よくエネルギーを獲得することができる。例えば,グルコース1molの酸化によって獲得できるATPは,好気条件下では36molであるのに対し,嫌気条件下ではわずか2molにとどまる。ATPに転換可能のGTPの生産量を加味しても,前者は38mol,後者は4molと大きく開いている。それでは嫌気性生物は息も絶え絶えに生育するのかというと決してそうではない。細胞の重量あたりの酵素活性を比較してみると,嫌気性細菌における各種酵素の中には,とびぬけて活性の高いものが少なくない。このように,エネルギーの転換,獲得の効率は悪くても,これを補うための別のしくみを巧みに利用して,嫌気性生物は活発に生育増殖を行っているのである。
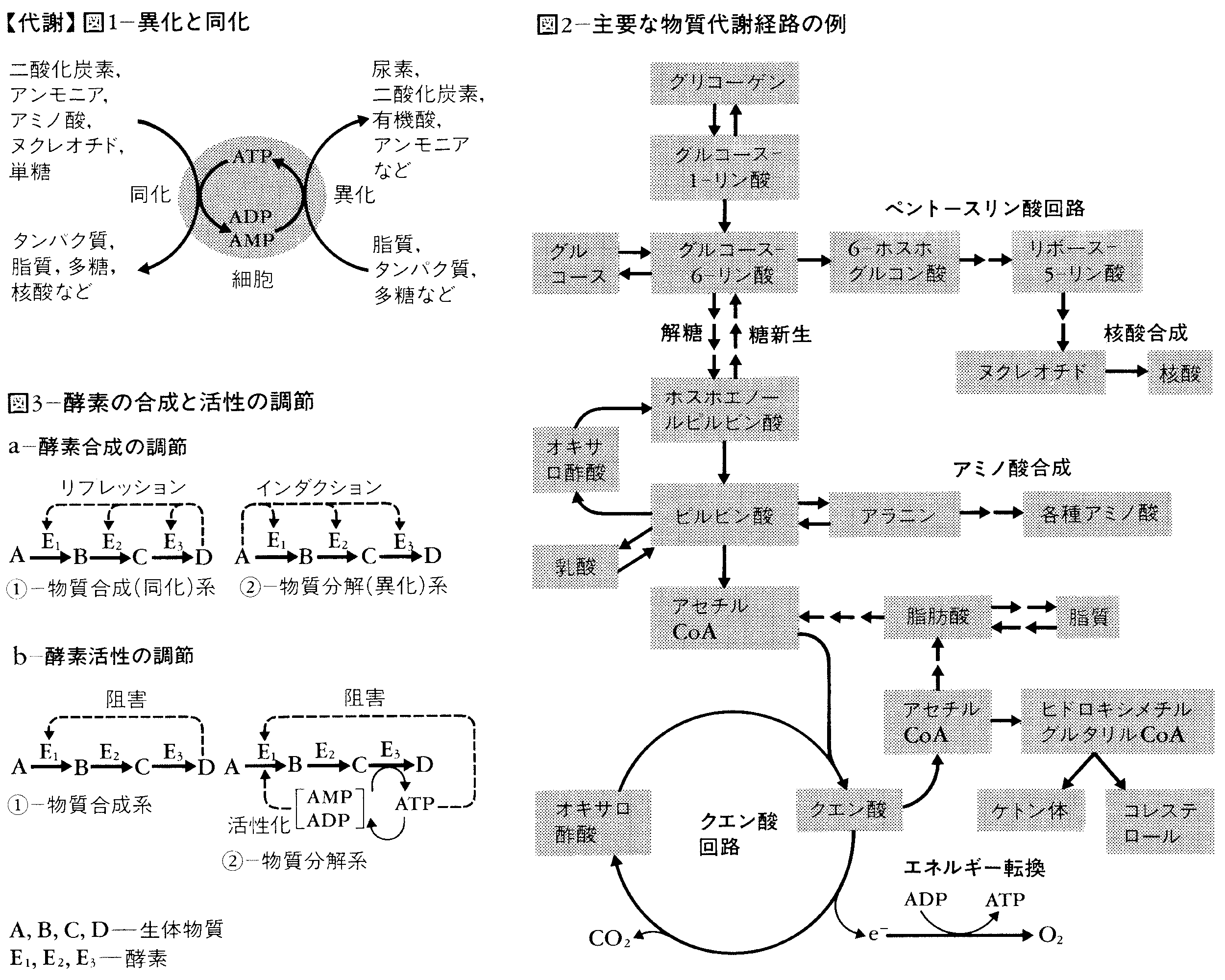
エネルギー代謝と物質代謝はつねに密接に関連した形で進行するが,エネルギーの獲得につながる物質代謝を異化catabolism,またエネルギーを消費する代謝を同化anabolismと呼ぶ(図1)。異化代謝においては,食物や生体内の複雑な化合物,すなわち,核酸,タンパク質,多糖,脂質などを分解して,乳酸,酢酸,二酸化炭素,尿素,アンモニアなどの簡単な化合物に変え,それらの過程で遊離するエネルギーを,ATPなどの高エネルギーリン酸化合物の形で貯蔵したり,直接運動エネルギーとして利用したりする。これに対して同化代謝は,アミノ酸,有機酸,単糖,脂肪酸,あるいはアンモニアや二酸化炭素などの比較的簡単な化合物を素材とし,ATPなどのエネルギーを利用して,より複雑な生体物質を合成する営みである。同化と異化は,その流れの途中にまったく同種の酵素反応を含むこともあるが,全体の流れとしては相互の妨げにならないように,種々のしくみが働いている。例えば,同化方向と異化方向にそれぞれ関与する酵素が別個のものであるとか,長鎖脂肪酸合成多酵素複合体のように,複数の酵素が順序よく並んだ集合体をつくり,そのまわりで代謝中間体(それらは個々の酵素にとっては順次基質,あるいは生成物となる)が,酵素分子とチオエステル性の共通結合を形成した形で最後のステップまで反応を進めるというようになっていたり,また後で述べるように,代謝の局在化による制御も行われている。同化と異化の一つの大きな違いは,後者はピルビン酸,アセチルコエンザイムA(アセチルCoA)といった,ごく限られた中間体をすべての代謝の流れが共通に経由するという特徴であろう。
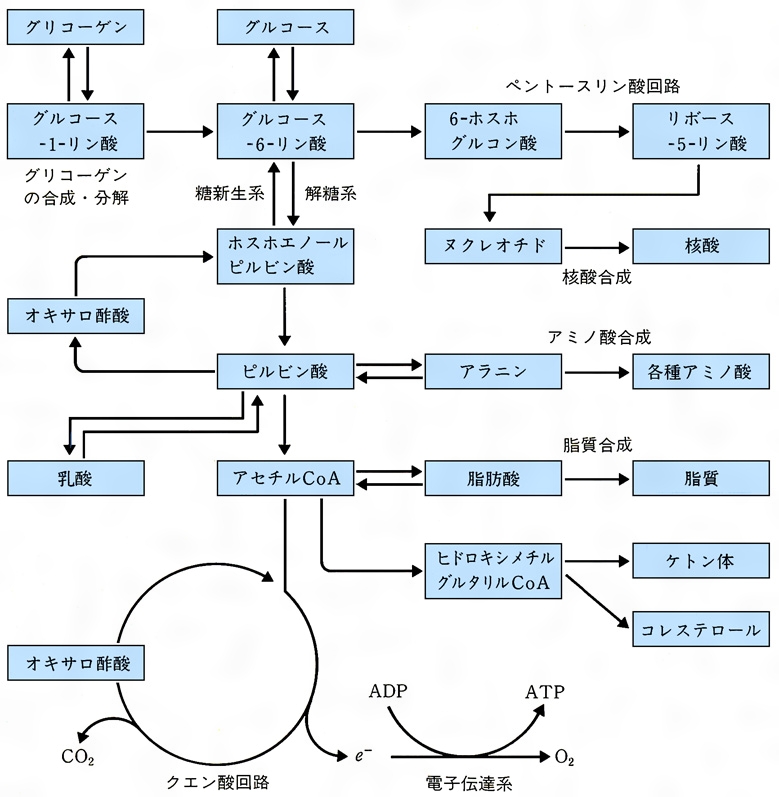
同化代謝と異化代謝の両方の目的にかなう両用代謝amphibolic metabolismの例もいくつか知られている。生体内で営まれている化学反応の種類はきわめて多いが,それらを生体物質の合成と分解という立場で整理してみると,案外限られた数にまとめられる。われわれの栄養素として最も重要な糖,脂質,タンパク質,そしてさらに種の保存と遺伝情報の発現,伝達に不可欠な核酸を中心に整理すると次のようになる(図2)。
(1)解糖 グルコースが嫌気的に分解して,ピルビン酸,ATP,NADH各2分子を生成する経路で,主として細胞質で進行し,各種生体物質の生合成に必要な炭素骨格の供給と,エネルギー源としてのATPの生産を行う。(2)クエン酸回路(TCA回路) 解糖系につづく好気性のエネルギー代謝回路で,主としてミトコンドリアで営まれる。炭水化物,アミノ酸,脂肪酸などがアセチルCoAを経て代謝され,ATP12mol,NADH3mol,FADH21mol,GTP1molが生成する。この回路の一員スクシニルCoAはポルフィリン生合成にも使われ,この回路は物質代謝の面でも無視できない。(3)五炭糖回路 核酸の前駆体であるヌクレオチド生合成の素材であるリボース-5-リン酸の供給やNADPHの生成のために重要な代謝で,主として細胞質で営まれる。(4)糖新生 乳酸,グリセロール,アミノ酸などの炭水化物以外の物質からピルビン酸の生成→炭酸固定によるオキサロ酢酸の合成→リン酸化と脱炭酸反応によるホスホエノールピルビン酸の生成を経てグルコースを合成する代謝で,おもに肝臓や腎臓で営まれる。(5)グリコーゲンの合成と分解 合成はUTPとグルコース-1-リン酸からUDPグルコースを経てグリコーゲン合成酵素が,また分解はホスホリラーゼが触媒する。(6)脂肪酸の合成と分解 合成は先に述べた多酵素複合体の関与のもとに細胞質で進行するが,分解反応の主流であるβ-酸化は細胞質で進行した後,アセチルCoAとなってミトコンドリア内のクエン酸回路に入るか,またはケトン体の合成に利用される。
→解糖 →クエン酸回路 →脂肪酸酸化回路
微生物,植物,動物に代表される各種生物の細胞あるいは体の中には,膜,壁,器官,組織,亜粒子(オルガネラ,細胞小器官)などの名称で呼ばれる,いろいろな部分が区別される不均一性構造が存在している。そしてそれぞれの部位では,特徴的な酵素と代謝が局在しており,しかも全体としてはみごとな調和を保ちつつ1個の生命を支えている。例えば先ほども述べたように,解糖や脂肪酸生合成は細胞質,クエン酸回路と電子伝達系はミトコンドリアで営まれる。細胞の外では血液中で働くプロテアーゼによる血液凝固や血圧の調節,また唾液(だえき)中のアミラーゼによる含水炭素性食物の消化,胃液中の酸性プロテアーゼによるタンパク質の加水分解など,体の要所要所で重要かつ特異的な代謝が進行している。
同位元素15Nを含むアミノ酸を食餌として動物に与え,肝臓のタンパク質を標識した後,その分解の速度を測定するという優れた手法で,シェンハイマーR.Schoenheimerは,ラット肝では5~6日という短期間で代謝が回転する事実を明らかにした。肝細胞自身は数ヵ月,また筋肉タンパク質や脳の脂質などはもっとゆっくり交替することが知られている。大腸菌などの単細胞性微生物は常温で約20分ごとに分裂するが,細胞1個の中に2000~3000種のタンパク質分子がそれぞれ400分子程度存在し,約1000種類の有機化合物が200万~300万分子つねに合成と分解をくり返しているといわれている。さらに,食物その他の外的要因や刺激の変動によって,物質とエネルギーの代謝回転はかなり大きく変化することも珍しくない。
エネルギー代謝についてはすでに触れたが,グルコース1molが完全燃焼するときに遊離する600kcalの自由エネルギーの約40%がATPに蓄えられている。無生物の世界ではとうてい考えられない高い熱効率である。解糖が,あるいはクエン酸回路が,リン酸エステルの生成と分解を含めたきわめて多くのステップを経て進行することの利点がそこに関連する。
生体が生命を維持するために必要な最小の効率を基礎代謝basic metabolism(BMと略記)と呼ぶ。筋肉作業などによる機能性消費を除外するため,絶対安静,絶食下,臨界温度において消費するエネルギーを指すが,成人1日の基礎代謝量は日本人で1200~1400kcal,欧米人で1500~2000kcalとされる。実測値と標準値の比を基礎代謝率basal metabolic ratio(BMRと略記)と呼び,臨床診断の資料として利用される。
すべての生物は外界から摂取した食物とエネルギーを利用して,上記の各種代謝を営んでいるのであるが,食物の内容が変わっても,また外界の気温その他の環境条件が変化しても,自己の体内の各種物質の濃度,例えば,血糖値,尿の組成,汗の中の塩分濃度などはつねにほぼ一定に保たれている。生物のこのような特徴を恒常性(ホメオスタシス)と呼ぶが,生物が外界との疎通をはかりながら自己の生命活動を独自に維持することができるのは,種々の複雑な代謝の流れを巧みに調節する機能を備えているためである。
代謝の調節のしくみは,生物の種類,器官や組織の違い,さらに代謝の内容などによって異なるものもあるが,ここでは比較的広い範囲で営まれているものを中心に述べる。最初に分子のレベル,次に生体のレベルで働くしくみに大別して考察してみよう。
(1)分子レベルにおけるしくみ 代謝の流れに沿って働く個々の酵素の量と活性の増減をひき起こす要因が種々知られている。酵素の量とは,活性型分子の濃度を指すが,その増減はまず酵素の主成分であるタンパク質の生合成と分解の調節の結果を意味する。タンパク質の分解を調節する分子機構がまだ統一的理解に達していないのに対し,生合成のほうはJ.モノーらのオペロン説に基づくインダクション(誘導)とリプレッション(抑制)のしくみによって巧妙に説明されている(図3)。活性型酵素の量はこのほか,ペプシノーゲン→ペプシン,キモトリプシノーゲン→キモトリプシンなどの例で知られる不活性前駆体(チモーゲン,プロ酵素)のプロテアーゼ限定分解による活性型への転換,不活性サブユニットの分子集合による活性発現,補酵素の供給などによっても調節される。一方,酵素の活性は,外部環境のpH,イオン強度,圧力,温度などの物理化学的要因,基質その他の非タンパク質性分子との相互作用,タンパク質-タンパク質相互作用などの影響で変動するが,それらの中でとくに代謝の自動制御に寄与する合目的的分子機構としてのフィードバック制御機構が重要である。その要点を図3に示すが,物質合成系において,最終産物が過剰に蓄積すると,その代謝系の初発段階(律速段階)の酵素E1の活性が特異的に阻害される。しかし,E2,E3の活性はまったく阻害されない。この点がリプレッションとの大きな違いである。これに対して,物質分解系においては,遊離するエネルギーを利用して合成されるATPが,その前駆体であるAMPまたはADPの濃度よりも低いレベルで存在する場合,これらの前駆体がE1の活性を賦活し,ATPの補給をはかるというしくみが知られている。高等動物の筋肉のグリコーゲンホスホリラーゼのAMPによる活性化,偏性嫌気性細菌ClostridiumtetanomorphumのトレオニンデアミナーゼのADPによる活性化などが典型的例である。上述のリプレッション→インダクション,フィードバック阻害→ヌクレオチド活性化のしくみはいずれも非共有結合性の調節であるのに対し,酵素タンパク質の特定のアミノ酸残基が,特定の酵素によって化学修飾されることによって,酵素分子の活性,pH依存性,フィードバック感受性などが大きく変化する場合がいくつか知られている。上記のグリコーゲンホスホリラーゼのb型は,その活性にAMPを必須とするが,ホスホリラーゼキナーゼとATPの関与によって,酵素タンパク質の特定のセリン残基がリン酸化されるとAMPが無くても約60%の活性が発現するa型分子に変換する。プロテインキナーゼのリン酸化,グルタミン合成酵素のアデニリル化なども同様な例である。
(2)生体レベルにおけるしくみ 膜透過を含む物質輸送,特定の酵素の局在化,さらに各種生理活性物質が関与する液性,神経性の調節など,生物種の違いや代謝の内容によって,多種多様な分子機構が相互に巧みな調和を保ちながら作動している。プロテアーゼ限定分解によるプロインシュリンのインシュリンへの変転と糖代謝の調節,ACTH(脳下垂体ホルモンの一つ)に端を発した限定分解産物としてのエンドルフィン,エンケファリンのような,いわゆるオピエート様ペプチドによる痛みの感覚の調節,血液凝固,血圧の調節など,広い範囲で分子レベルのしくみが高次の調節にも寄与する。
執筆者:徳重 正信
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
生体内のあらゆる物質変化の総称で、物質代謝、新陳代謝ともいう。また、生体がその生命活動を維持するための最小必要量のエネルギー量を基礎代謝という。生体の細胞内に存在する物質を同定してみると、その生物の摂取する食物が細胞内に存在するすべての分子を含んでいるわけではなく、生体内に取り入れられてから大きな変化を受けてつくられることがわかる。極端な場合には、グルコース以外のすべての有機物が、その体内で合成されるという生物もある。これらの変化は一段階でおこるのではなく、一連の化学反応を経て、いろいろな中間体を通る。この過程でみられる変化を中間代謝という。
代謝は、比較的簡単な分子から複雑な化合物を合成する反応(同化)と、高分子や巨大分子など複雑な化合物から簡単な分子に分解する反応(異化)に大別できる。同化の過程はエネルギーを要求し、異化の過程ではエネルギーが遊離される。このように物質代謝とエネルギー代謝は密接に共役している。生体物質の合成と分解の反応は一連の化学反応によって達成されており、その道筋を代謝経路とよぶ。生体内でみられる化学反応は多数あるが、それらを整理すると、ごく限られた代謝経路を中心としていることがわかる。
[飯島康輝]
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
 山に登る〕詩 人事、代謝
山に登る〕詩 人事、代謝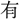 り
り 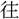 來、古今を
來、古今を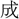 す 江山、
す 江山、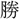 跡を留め 我が輩復(ま)た登臨す
跡を留め 我が輩復(ま)た登臨す字通「代」の項目を見る。
出典 平凡社「普及版 字通」普及版 字通について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
Sponserd by ![]()
[同義異語]物質代謝
出典 森北出版「化学辞典(第2版)」化学辞典 第2版について 情報
Sponserd by ![]()
福岡県福岡市博多区の櫛田神社の夏祭り。壮麗な山笠で知られる。今日,山笠には飾り山笠と舁き山笠(かきやまがさ)の 2種類がある。明治時代に電線が架設されて以降,物語場面の人形などを飾りつけた高さ 15m...