デジタル大辞泉 「染色体」の意味・読み・例文・類語
せんしょく‐たい【染色体】
[類語]細胞・細胞膜・細胞壁・細胞質・原形質・単細胞・胚・核・核酸・リボ核酸・デオキシリボ核酸・遺伝子・性染色体・ミトコンドリア・組織・胚珠・胚乳・胚芽
Sponserd by ![]()
翻訳|chromosome
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
もともとは,細胞分裂時に現れる,塩基性色素によく染まる糸状構造体のことをいったが(W.ワルダイアー,1888),T.H.モーガンらの研究により,この構造体に遺伝子が線状に配列していることがわかってから,遺伝学的には,個々の連鎖群に対応する遺伝的単位,すなわち線状に配列した一連の遺伝子をさすようになった。また,細胞学的には細胞分裂時にみられる糸状構造体に限らず,中間期の核において極度に伸びきって糸状構造として認められなくなっているものや,細菌の核域,ウイルス粒子,真核生物のオルガネラなどに存在する個々の遺伝情報分子をも染色体と呼ぶようになってきている。したがって,染色体の今日的定義としては,遺伝学的または細胞学的(電顕観察を含む)に一つのまとまりをなす遺伝情報担体とするのが適当である。その物質的実体は,真核生物の核においてはDNAとヒストンが結合した巨大糸状分子であり,ウイルス,プラスミドを含む原核生物や,真核生物のオルガネラにあっては,裸の単一DNA分子(離脱しやすいタンパク質が結合している可能性はある)である。以下には真核生物の核に存在する染色体を中心に述べる。
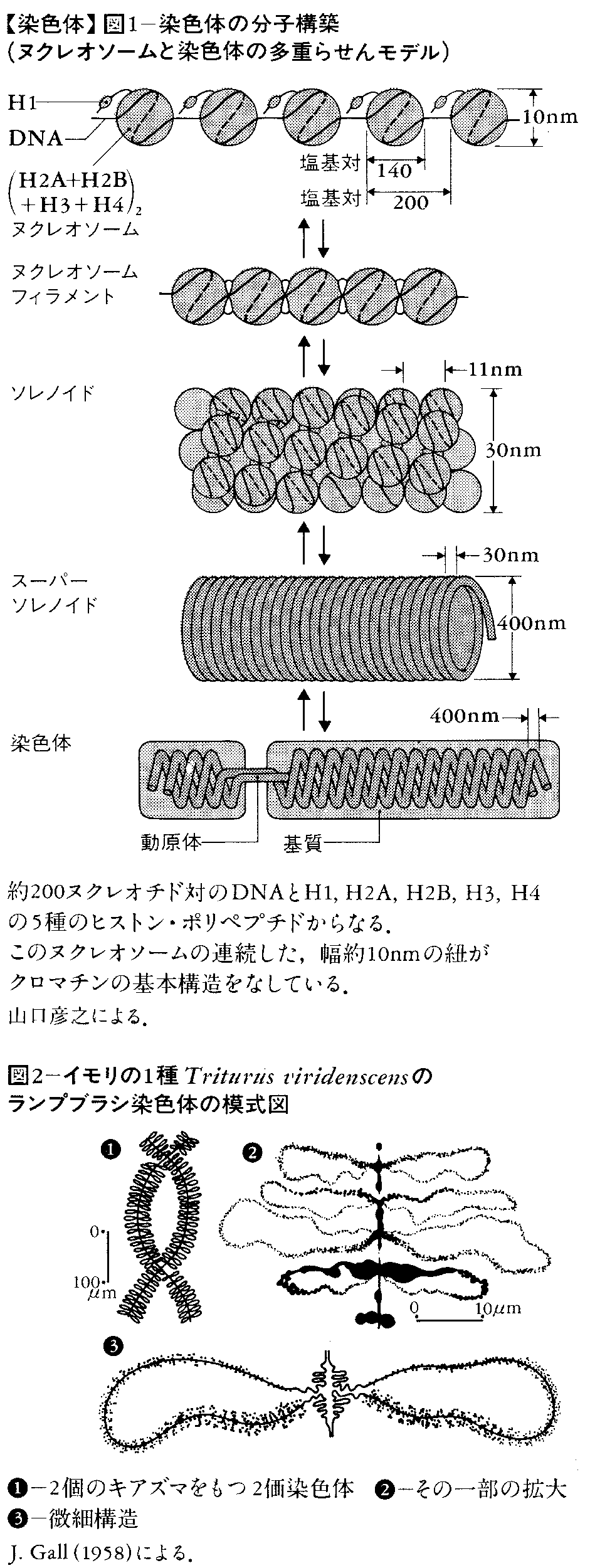
染色体ごとに量的に一定した,ほぼ等量のDNAとヒストンからなり,ほかに環境条件などによって量的に変動するRNA(転写産物)や非塩基性タンパク質(RNAポリメラーゼなど)が含まれる。基本単位は図1に示すようなヌクレオソームnucleosomeである。
染色体は細胞分裂中期にもっとも強く凝縮して太さが1μmにも達するが,中間期の核では分散して,クロマチンの形をとる。いろいろの間接的証拠から,分裂中期の太く凝縮した染色体は,連続したヌクレオソームの2本の紐からなる(複製が終わっているので)と考えられる。細胞分裂に入って染色体が凝縮し,太さを増す機構はまだ十分に解明されていないが,その機構についていろいろのモデルが提案されている。このうちの一つ,多重らせんモデルを図1に示してある。このモデルの基本繊維は幅2~3nmの二重らせん構造のDNA(一次らせん)であり,幅約10nmの二次らせんはヒストン核にDNAがらせん状に巻きついたヌクレオソームの紐にあたる。それより高次のらせんは分裂中期の染色体の大きさを説明するための仮想的な構造である。
DNA分子が半保存的な複製様式をとることと,染色体がヌクレオソームの連続した紐よりなることを考えると,染色体の複製も半保存的に行われるものと思われる。これを証明するため,J.H.テーラーはソラマメやヒアシンスの根端を,トリチウム3Hでラベルしたチミジンを含む水に短時間つけ,この間に複製した染色体(厳密にはDNA鎖)を3Hで標識した(1957,58)。そして,その後の細胞分裂でこの標識がどのように娘染色体に伝わるかを追跡したところ,第1回目の細胞分裂では2本の娘染色体がともに全長にわたって均一な標識を示したが,第2回目の細胞分裂では娘染色体の片方だけが標識されていた。このことから,DNAと同じように,染色体も半保存的に複製することがわかった。
染色体中のDNAの複製は中間期に行われる。この時期をとくにS期という。成熟分裂では第1分裂に入る前の中間期にDNAの複製が行われ,第1分裂と第2分裂の間では複製が起きない。
原核生物ではふつう一つの染色体が一つの複製単位,すなわちレプリコンrepliconを構成しており,複製開始点が染色体あたり1個しかない。これに反し,真核生物の染色体は多数の複製開始点をもち,それと同数のレプリコンから構成されている。この染色体ではDNAの複製は,各開始点から左右両方向へ進行する。複製の開始時期は開始点によって多少のずれがあり,ふつう,真正クロマチン部分ではS期の前半に,ヘテロクロマチン部分ではその後半に複製が行われる。
S期から体細胞分裂中期にかけての染色体は,DNA複製は完了しているが染色体分離はまだなので,二重構造になっている。この時期の染色体の二重構造の片方ずつを染色分体chromatidという。光学顕微鏡でこの分体構造が明らかになるのは,分裂前期の終りである。成熟分裂では複製を終えた相同染色体が成熟分裂第1分裂前期の早い時期(接合期)に対合するので,この時期から第1分裂中期までの二価染色体は四重構造をなしている。この構造が光学顕微鏡でわかるのは第1分裂前期の複糸期以降である。
(1)染色体の長さ 染色体の相対的な長さは,その遺伝情報量と高い相関を示す。パンコムギの21本の染色体の長さとDNA量の間には+0.82の相関係数が得られているし,キイロショウジョウバエの4本の染色体の長さは連鎖群の全長や唾腺(だせん)染色体のバンド数と高い正の相関を示す(表)。
(2)動原体 この部位は塩基性色素に染まらず,染色体のくびれのように見えるので,第1次狭窄(きようさく)とも呼ばれる。ここは細胞分裂時に形成される微小管の付着する部位になっており,染色体の分裂前期における核板への移動(前期移動)と分裂後期における両極への分離(後期移動)に決定的な役割を果たす。動原体を欠損した染色体はいずれの移動も行いえず,細胞質にとり残されて消失するし,二つ以上動原体をもつ染色体は後期移動において,それぞれの動原体が機会的に移行する極を選択するため,高い頻度で染色体橋を形成し,そのため,切断したり細胞質にとり残されたりする。
(3)仁形成体 仁nucleolusと呼ぶ球形の小構造が中間期の核に見られる。この仁は特定染色体の仁形成体と呼ぶ部位に付着していることが,細胞分裂前期または後・終期の核の観察によって確かめられる。動原体と同じように,仁形成体部位も塩基性色素に染まらず,染色体のくびれのように見え,そのため第2次狭窄とも呼ばれる。この部位に,転写によってリボソームRNA(rRNA)をつくる遺伝子,すなわちrRNA遺伝子(または単にrDNA)が高度に重複して存在する。例えば,キイロショウジョウバエの第Ⅰ染色体やトウモロコシの第Ⅵ染色体の仁形成体部位には,それぞれ約100および数千コピーのrRNA遺伝子が存在する。
(4)ヘテロ(異質)クロマチン ふつうの遺伝子を含む真正クロマチン部位に比べ,色素に対する染色性が異なるのでこの名がある。組成的には両クロマチンともDNAとヒストンからなるが,ヘテロクロマチンには長さのいろいろな特定塩基配列(短いものでは数ヌクレオチド対から長いものでは数百ヌクレオチド対までいろいろの種類がある)が数千コピーも重複して存在する。そのため,ふつうの意味での遺伝子は存在せず,遺伝的に不活性である。
ヘテロクロマチンはふつう動原体の近くや染色体の末端部に見られるが,真正クロマチンに挟まれて存在する場合も少なくない。これを介在ヘテロクロマチンという。成熟分裂第1分裂前期の太糸期において介在ヘテロクロマチンがこぶ状に見えることがあるが,これをとくに染色体瘤(りゆう)chromosome knobと呼んでいる。ヘテロクロマチンは,細胞がどのような生理的条件におかれたときにも存在する構成ヘテロクロマチンと,条件により消失したり出現したりする機能ヘテロクロマチンに分けられる。染色体をキナクリンやキナクリンマスタードで染色し蛍光顕微鏡でみると,構成ヘテロクロマチン部位は蛍光で光ってみえる。この部位をQバンドという。また,染色体を加熱やアルカリ(例えば水酸化バリウム)処理したあと,ギムザ液(アズール色素,エオシン,メチレンブルーの混合液)で染色すると,構成ヘテロクロマチンは濃紫色に染まる。この部位をGまたはCバンドとよぶ。これらのバンドは染色体を識別,同定するための構造的特徴として利用される。
(1)形態的分類 動原体が染色体の中央に位置すると,後期移動のときV型にみえる。その位置が多少端に寄ったり,末端に移ったりすると,J型やI型になる。そこで,分裂後期の形態から染色体をV,J,Iの3型に分けることがある。また,染色体の長・短両腕の比から次の4型に分けることもある。短腕/長腕の比がほぼ1のとき中部動原体染色体,1/2より大きいとき次中部動原体染色体,1/2より小さいとき次端部動原体染色体,短腕が欠けているとき端部動原体染色体という。
(2)性染色体と常染色体 性と結びついて染色体に2型がある場合,それを性染色体という。これに対し,雌雄両性の間に違いのみられない相同染色体を常染色体autosomeという。
(3)B染色体とA染色体 ヘテロクロマチンだけからなる染色体をB染色体,または過剰染色体という。これに対し,真正クロマチンを含むふつうの染色体をA染色体という。B染色体は,その数や形が同一の種または系統に属する個体の間で違うことが多い。この型の染色体は遺伝的に不活性であり,その数がとくに多くならないかぎり個体の形質に大きな影響を与えることはない。
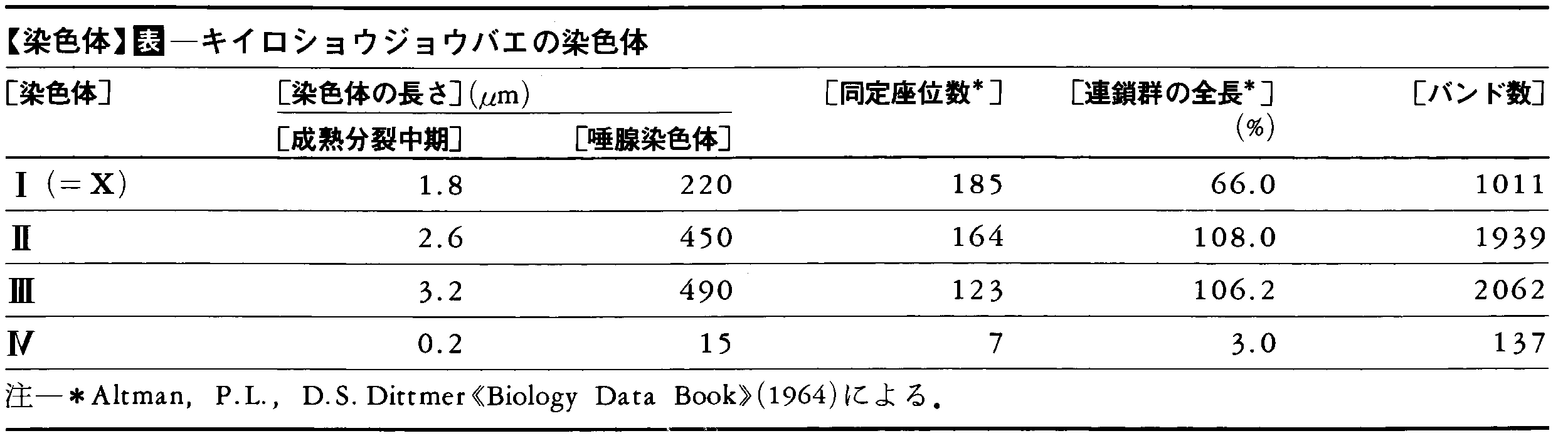
(4)巨大染色体giant chromosome ふつうの体細胞分裂や成熟分裂には見られない大きい染色体が特殊な細胞で見られることがある。その代表的なものは双翅(そうし)目の昆虫の唾腺細胞に見られる唾腺染色体である。このような巨大染色体は他の分泌細胞でも見られる。昆虫の腸上皮細胞,マルピーギ管細胞,哺育細胞,植物の胚囊の反足細胞,胚盤細胞,などがその例である。もう一種の巨大染色体は,脊椎動物の卵母細胞の成熟分裂第1分裂前期に見られるランプブラシ染色体である。この名は図2に示すようにその形からきており,サメ,鳥類,両生類でよく研究されている。イモリの1種Triturus viridenscensでは,個々の相同染色体対の長さは350~800μmあり,総長は6mmにも達する。これは複糸期ないし移動期にある染色体が高次らせん構造を部分的に解き,受精後の胚発生に備えて染色体の各所で転写を行っている姿とみなされている。染色体の全長にわたって見られるループ状の突起の一つ一つは転写の単位をなすDNAであり,これに転写でつくられつつあるたくさんのRNA分子が付着しているのが電顕で見られている。
染色体の構造異常には重複,欠失,逆位,転座の4種がある。
(1)重複duplication 染色体の一部の遺伝子配列とまったく同一の遺伝子配列が,同一または別の染色体に存在していること,またはそのような状態になること。重複が同一染色体の同一部分に起こっているとき,とくに反復repeatという。重複はゲノム全体の遺伝的調和を大きく乱すことなしに,単一または少数の遺伝子の量を増やすことを可能にする。仁形成体部位に見られるrRNA遺伝子の反復はその好例である。キイロショウジョウバエの唾腺染色体には,同形同大のバンドが2本隣りあった二重バンドが全バンド数の約70%を占めている。これは,単一またはごく少数の遺伝子の重複がいかに頻繁にこのハエの進化の過程で起こったかを示している。
(2)欠失deficiency(deletion) 染色体の一部が欠けること,または欠けた状態。2倍性の生物では欠失のホモ接合体はふつう生存できない。したがって,欠失は自然界にはヘテロの形でしか存在しない。欠失はしばしば優性突然変異としてとらえられる。これは遺伝子の量がヘテロ個体で2から1に減ずることに伴い遺伝子の量的効果が現れるからである。ただし,遺伝子の一部分が欠失した場合には,劣性遺伝子としてふるまうことが多い。欠失の位置や大きさは,唾腺染色体や成熟分裂太糸期染色体の観察から決められる。
(3)逆位inversion 標準染色体に比べ,遺伝子の配列順序が染色体の一部において逆転すること,または逆転している状態。逆位は一つの核に含まれる遺伝子の種類や数になんの変化もきたさない。逆位をホモにもつ個体は,その形質のみならず,成熟分裂や配偶子の生存力など生殖に関する諸性質についても,標準型の個体と変わるところがない。ただし,逆位を含む連鎖群では,逆位の中にある座位と外にある座位の間で,連鎖地図上の距離が違ってくる。逆位ヘテロの個体では成熟分裂に異常が生じる。標準染色体と逆位染色体が対合する際,逆位部分においてループを形成する。このループ内で交叉(こうさ)が起こると,交叉に関与した染色体に重複と欠失が起こり,この染色体を受けとった配偶子は受精力を失う。そのため,逆位は実質的に交叉抑制因子として働く。
(4)転座translocation 染色体の一部分がその染色体の別の位置や他の染色体に移ること,または移った状態。転座の場合も一つの核に含まれる遺伝子の種類と数には変化がなく,転座ホモ個体の諸形質は標準型個体と同じになる。しかし,転座に関係した染色体では,遺伝子の新しい連鎖関係を生じる。2本の相同染色体の間で,それぞれの一部を交換する場合を相互転座という。例えば第Ⅰ染色体と第Ⅱ染色体が互いにその片腕を交換するようなケースである。こういう相互転座をホモにもつ系統と標準系統の間で交雑が起こると,相互転座ヘテロ個体を生ずる。このような個体の成熟分裂では四価染色体が形成され,高い頻度で不稔(不妊)の配偶子が形成される。そのため,相互転座ホモ系統は標準系統からある程度生殖的に隔離される。相互転座が種分化の一つの原因となる理由がここにある。
執筆者:常脇 恒一郎
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
生物の細胞内に含まれるデオキシリボ核酸(DNA)を主成分とした自己増殖性のある小体。染色体の基本構造は、ウイルスやバクテリアなどのような下等生物から高等動植物に至るあらゆる生物を通じ同じである。下等な原核生物では二重螺旋(らせん)構造をしたDNAが裸の形で細胞内に含まれているが、高等な真核生物の染色体はDNAのほかにリボ核酸(RNA)、塩基性タンパク質(ヒストン)、酸性タンパク質および少量の脂質や多糖類が加わり、複雑な長い紐(ひも)状構造(染色糸)となって細胞核内に含まれている。細胞分裂が始まると、核をもたない原核生物ではDNAがそのまま二分して2細胞に移行するが、核をもつ細胞(真核生物)では染色糸は短縮して塩基性色素によく染まり、光学顕微鏡で観察可能な小体に発達する。この小体の数と形は生物の種類により一定で、従来これを染色体とよんだ(狭義の染色体)。しかし、最近では原核生物の遺伝物質(DNA)も染色体とよんでいる。真核細胞における分裂期の染色体は、ヘマトキシリンやカーミンなど塩基性色素によく染まるのでその名がある。休止核の染色糸は光学顕微鏡では判別できないが、電子顕微鏡でみると太さ約0.3マイクロメートルの細長い糸状構造として観察される。この時期にDNAの複製がおこり、分裂中期にみられる染色体はつねに2本の娘(じょう)染色体からなっている。分裂後期では染色体は1本ずつの娘染色体に分かれ、それぞれ娘細胞に入る。
分裂中期の染色体の大きさは生物の種類によって多少違うが、ヒトの細胞を例にとると、染色体の太さ(直径)は約1~2マイクロメートル、長さは数マイクロメートルから十数マイクロメートルである。染色体の数と形は、生物の種類により一定で、生物種の重要な特徴となっている。
染色体の形は生物の種類により一定しており、それを核型という。体細胞は形の等しい2組の染色体(相同染色体)から成り立っている。1組は母方(雌親)から、他方は父方(雄親)から由来したものである。一般に体細胞の染色体数は2nで、生殖細胞のそれはnで表される。ヒトを例にとって染色体の構成を調べてみると、体細胞の染色体(2n)は46個で、そのうち23個(n)は卵子から、ほかの23個(n)は精子由来である。23対のうち不等の1対は性決定に関与する性染色体で、大きいほうをX、小さいほうをYで表す。性染色体以外の22対の染色体を常染色体とよび、これらは大きさと形によりAからGの7群に分類されている。従来の染色法では各群内の個々の染色体対(つい)の識別は困難であったが、最近、発展したバンド染色法ですべての染色体対の識別が可能となった。
染色体上の遺伝子を相対的距離によって配列し、図式によって表したものを染色体地図という。互いに連関して遺伝する任意の二つの遺伝子を組換え価から直線上に配列してみると、遺伝子間の相対的な距離を示す地図が描かれる。これを連鎖染色体地図とよぶ。染色体地図には染色体の転座、重複、欠失などの染色体異常と遺伝子の位置を対応させたり、異種間の雑種体細胞をつくって雑種細胞における染色体の脱落分離から染色体と遺伝子を対応させる細胞学的染色体地図がある。最近、ヒトとマウスの雑種細胞からヒトの詳細な細胞学的染色体地図がつくられている。双翅(そうし)類の唾液腺(だえきせん)染色体(唾腺染色体)の横縞(よこじま)模様と遺伝子の位置を結び付けたものを唾腺染色体地図とよぶ。
染色体の逆位や転座などの異常が、ある生物種の自然集団中につねにある割合で含まれていることがある。これを染色体多型という。また、同一種でありながら生息する地域で異なった核型を示すこともあり、これを地理的変異という。これらはいずれも生物種の分化や進化の要因となっている。
染色体を最初に観察したのは1842年ネーゲリであり、その後1875年にストラスブルガーStrasburger、1880年にW・フレミングなどによって確かめられた。染色体chromosomeという名称は、1888年ワルダイヤーWaldeyereによるもので、ギリシア語のchroma(色)とsoma(体)から名づけられた。それを染色体と訳したのは1892年(明治25)石川千代松である。
[吉田俊秀]
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
細胞分裂のときにみられる遺伝子の集合体.“よく染まるもの”という意味からこう名づけられた.その化学組成は原核細胞ではほとんどDNAであるが,真核細胞ではDNAのほかに塩基性タンパク質(ヒストン,プロタミン),それに少量の非塩基性タンパク質,リボ核酸(RNA)などを含んでいる.細胞中のDNAのほとんどは染色体に局在している.染色体の形と数は各生物により定まり,そのなかには雌雄を決定する性染色体も含まれている.また,いくつかの遺伝的疾患が染色体異常と関連していることが明らかにされている.
出典 森北出版「化学辞典(第2版)」化学辞典 第2版について 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
Sponserd by ![]()
…いろいろの遺伝様式のおもな特徴と相互関係を表1にまとめてある。
[相同染色体と対立遺伝子]
核には決まった数の染色体があり,一つ一つの核内遺伝子(以下,遺伝子という)はいずれか1本の染色体の一定の部分を占めている。いいかえれば,遺伝子が線状に配列したものが染色体である。…
…
[細胞遺伝学cytogenetics]
メンデルが遺伝の法則を発見したころからそれが再発見されるまでの19世紀最後の四半世紀は,細胞学がメンデルの法則,したがって遺伝学そのものを生物学へ受け入れるための準備をした期間である。この間,受精において卵と精子の核が合体すること,細胞分裂において染色体が縦裂し,その半分ずつが娘細胞に分かれて入ること,卵と精子から同数の染色体が接合体にもち込まれることなどが知られてきた。そして,メンデルの法則の再発見後間もなくサットンW.S.Sutton(1902)などによって成熟分裂(減数分裂)における染色体数の半減が発見されるに及び,メンデル因子と染色体の行動に完全な並行関係がなり立った。…
…このような遺伝因子はメンデル因子,または単に因子とよばれていたが,W.L.ヨハンセン(1909)の提案した遺伝子という語がしだいにこれにとって代わるようになった。 サットンW.S.Sutton(1902)らはいち早く成熟分裂における染色体の行動がメンデル因子の行動と一致することを明らかにしたが,さらに,1910年に始まるT.H.モーガンらのキイロショウジョウバエの研究により,遺伝子は染色体に線状に配列して連鎖群を形成しており,一つの遺伝子は特定の染色体の特定の部位を占めていることがわかった。このような研究により,それまで仮想的な存在であった遺伝子が物質的基礎をもつことになり,その構造と機能を物質的に研究する道が開けた。…
…この病気は,メンデルの遺伝の法則にしたがうので,メンデル遺伝病Menderian diseaseとか古典的遺伝病とか呼ばれる。ヒトの染色体は46本あり,うち44本は男女同じで常染色体と呼ばれ,残り2本は,男でXY,女でXXの構成をもち,性染色体と呼ばれる。メンデル遺伝病には次のような遺伝形式をとるものがある。…
…明らかに原始的とされる原核細胞は,細胞の大きさが1μm程度で小さく,通常細胞壁の形成によって決まる単純な形態をとっており,一部を除いて原形質にはとくに発達した構造が見あたらない。原核細胞と名付けられるとおり,核膜によって囲まれる原形質の区分(核質とよぶ)がなく,遺伝子を配列する染色体は,ほとんどDNAの1分子と考えられ,核様体とよばれる構造をとって原形質中に広がっている。原始的な細胞生物である細菌類は,すべて原核細胞に属しているが,なかでも最小の細胞といわれる球形のマイコプラズマは直径0.1~0.25μmの範囲にあり,細胞壁はなく,細胞膜のみによって囲まれる原形質には微細構造がきわめて乏しく,最小の染色体(長さ500μmにも達しない環状DNA)と最少数のリボソームが含まれるにすぎない。…
※「染色体」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
日本の年中行事。年末に天井や壁にたまった煤を取除き大掃除をすること。近年は正月休みに入る 12月 29日とか 30日が多いが,伝統的には 12月 13日に行なった。この日は正月を迎えるための準備を開始...