デジタル大辞泉 「ウイルス」の意味・読み・例文・類語
ウイルス(〈ラテン〉virus)
2 「コンピューターウイルス」の略。
[類語](1)菌・細菌・バクテリア・球菌・乳酸菌・黴菌・雑菌・病原菌・病原体・大腸菌・サルモネラ菌・ピロリ菌・ヘリコバクターピロリ・スピロヘータ・リケッチア・酵母・イースト・黴・青黴・麹黴
Sponsored by ![]()
翻訳|virus
Sponsored by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponsored by ![]()
細胞性生物とは明確に異なる分子的構成と、独特な増殖相をもつ非細胞性構造体の一群をいう。
ウイルスは、それ自体を複製させうる宿主(しゅくしゅ)(ウイルスの寄生対象となる生物)細胞内に入って増生(増殖)し、それに伴って宿主にいろいろの影響を及ぼす。タバコモザイクウイルス(TMV)がタバコの葉にモザイク症状をおこし、口蹄疫(こうていえき)ウイルスがウシの口の粘膜や蹄部皮膚に水疱(すいほう)を生ずるなどがその一例である。これらの諸症状をもたらすものは、細菌よりも小さいため光学顕微鏡では観察できないし、また、細菌濾過器(ろかき)を水とともに通過する濾過性の粒子であるが、病原性のあることは、すでに19世紀末に発見されており、ラテン語で「毒」を意味するウイルスの名が用いられるようになった。その後、いろいろのウイルスが知られ、さらに電子顕微鏡によって形態や性状に対する研究が飛躍的に進んだ。
学術用語としては「ウイルス」が採用されているが、ほかにビールス、ウィルス、バイラス、濾過性病原体などの語が使用されてきた。
[曽根田正己]
ウイルスは、増生のための遺伝情報を担う核酸(デオキシリボ核酸=DNA、またはリボ核酸=RNA)をもつ核タンパク質の構造体で、大きさは一般に、微小細菌であるリケッチアの半分以下(20~300ナノメートル。1ナノメートルは10億分の1メートル)である。ウイルスは、構成する核酸の種類によってDNAウイルス、RNAウイルスに2大別されたり、あるいは宿主によって植物ウイルス、動物ウイルス、細菌ウイルスに3大別、または昆虫ウイルスを区別して4大別されるが、植物と昆虫、恒温動物と昆虫のように2宿主の両方で増殖するものもある。また、1種類の宿主にのみ寄生するものもあれば、キウリモザイクウイルスのように宿主が100種以上のものもある。植物ウイルスはRNAウイルスであり、タンパク質以外にポリアミンや金属を含むウイルスもある。また、脂質、炭水化物を含むウイルスもみられるが、これらを含むものは、むしろ動物ウイルスに多い。動物ウイルスとバクテリオファージ(細菌ウイルスまたはファージともいう)には、DNAウイルスとRNAウイルスとがある。バクテリオファージのなかには一般細菌や放線菌につくものがある。また、菌類である酵母につくウイルスや、アオカビ、コウジカビなどのようなカビ、さらに、キノコにつくウイルスもあり、これらはマイコファージといわれている。
[曽根田正己]
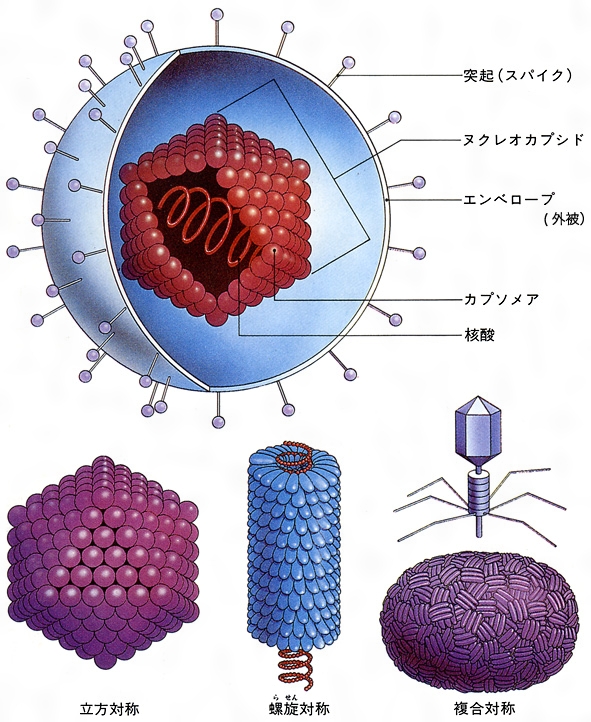
感染性をもつウイルス粒子の完全形態をビリオンとよぶ。ウイルス粒子の構造や形態はグループ(基本的には科)によって違いがある。
ウイルス核酸とそれに結合しているタンパク質を取り囲むタンパク質殻をカプシドとよぶ。ウイルス遺伝子とカプシドをあわせたものがヌクレオカプシドである。ウイルスによってはカプシドの外側にエンベロープ(外被)がある。パラミクソウイルスなどはカプシドとエンベロープの間にMタンパク質、ヘルペスウイルスにはテグメントとよばれるものがある。カプシドは小さいタンパク質粒子である多数のカプソマーからできている。
ウイルスの核酸はDNAかRNAのいずれかである。それぞれ二重鎖(2本鎖)DNA、単鎖(1本鎖)DNA、二重鎖(2本鎖)RNA、単鎖(1本鎖)RNAの4種類がある。多くのものが二重鎖DNA、単鎖RNAである。DNAは線状のものが多いが、環状のものもある。RNAの多くは、一つの構造体であるが、いくつかの分節(断片)になっているものがある。A型、B型インフルエンザウイルスは8個、レオウイルスは10個、ロタウイルスは11個、ブニヤウイルスは8個の分節からなる。また、レトロウイルスのように同じものを2組もつものもある。
ウイルスの核酸は宿主の力を借りて自分と同じものを複製する。ウイルス遺伝子の完全な1組をウイルスゲノムとよぶ。ウイルス遺伝子の数は3個から200個であり、細菌類などより少ない。
ウイルスの核酸はウイルスの感染性を担っていると考えられる。ウイルスによっては、ウイルス粒子から抽出した核酸だけを宿主細胞に注入すると、これだけで感染がおこり、同じ型の子のウイルス粒子がつくられていく。このような核酸を感染性核酸とよび、この現象をトランスフェクションTransfectionという。
カプシドはウイルス核酸を包むが、またウイルス核酸のもつ遺伝情報によって形成される。ウイルス核酸のもつ遺伝情報の数は少ないため、ウイルスゲノムが遺伝暗号としてもつことのできるタンパク質の種類は限られている。
カプシドはその外形により立方対称性と螺旋(らせん)対称性に分類される。アデノウイルスは立方対称性の例であり、タバコモザイクウイルスは螺旋対称性の例の一つである。
動物ウイルスの大多数は正二十面体対称型である。正二十面体は5個の正三角形が集まった形をとり、12の頂点をもつ。カプシドはウイルス核酸を保護し、ビリオンを宿主細胞の表面上のレセプター(受容体)に結合させることにより吸着し、細胞内に侵入する糸口となる。また、ウイルスの宿主域を決定し、ウイルスそれぞれの抗原性に関与する。
エンベロープはオルトミクソウイルス、パラミクソウイルス、ヘルペスウイルス、レトロウイルス、ラブドウイルスなどウイルス粒子の外側にある。感染した宿主細胞の細胞膜に由来する。このように螺旋対称型ヌクレオカプシドをもつ動物ウイルスはすべてエンベロープを形成する。エンベロープの表面にはスパイクspikeとよばれる突起がみられる。これは糖タンパク質でできている。エンベロープは糖タンパク質と脂質からでき、ウイルスが宿主細胞表面に吸着し、感染の際に重要な役割をもつ。
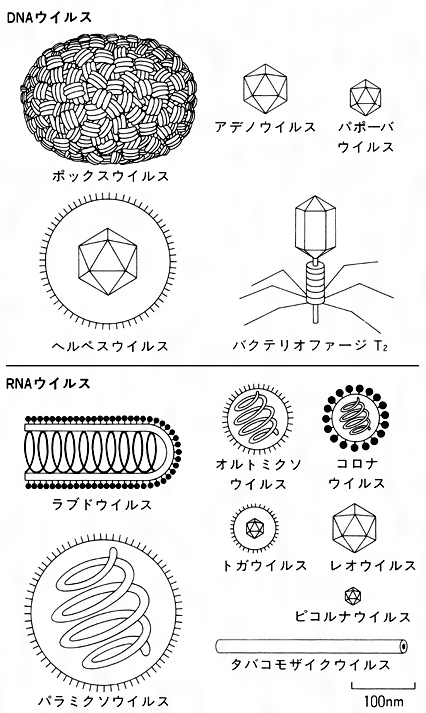
大多数のビリオンは対称構造を示す。タバコモザイクウイルスなどはエンベロープをもたない円柱状の螺旋対称型ヌクレオカプシドであり、アデノウイルスなどはエンベロープをもたない正二十面体状の立方対称型のヌクレオカプシドであり、パラミクソウイルスは円柱状の螺旋対称型ヌクレオカプシドがエンベロープに囲まれ、ヘルペスウイルスは正二十面体状の立方対称型のヌクレオカプシドがエンベロープに囲まれている。ポックスウイルスやバクテリオファージは一部のビリオンが対称構造とならない、複雑な構造をもつものもある()。
[曽根田正己]
ウイルスは宿主細胞内に侵入し、ある期間を経て細胞外に放出されるまでの間にその数を増やす。この侵入から放出までの経過は、ウイルス増生の経過でもあり、種数によってさまざまである。これはウイルス自身の性質が第一の因子となるが、素材の供給を受ける宿主細胞の形態や構造も重要な因子となる。
バルボウイルスのような小さなDNAをゲノムにもつウイルスの場合は宿主細胞への依存度が高く、ワクシニアウイルスのように大型の複雑な構造をもつ場合は核酸やタンパク質をつくるのに必要な素材だけを得られれば、自分自身の酵素活性によって自己複製が可能となる。
一般にウイルスの増生は、宿主細胞のウイルスの吸着→侵入→脱殻・解体→ゲノムの複製→素材からの合成→集合→成熟・放出(脱出)の経過をたどる。
(1)吸着 吸着はウイルス粒子の表面タンパク質と宿主細胞膜上のウイルスレセプターとの特異的な結合による。とくに、レセプターのもつ意味は重要であり寄生性を決める重要な因子となる。ヒト免疫不全ウイルス(HIV)は細胞にレセプターとしてCD4(CD=Cluster of Differentiation。分化抗原群、細胞表面の分化抗原)が発現している細胞、T細胞、マクロファージ、樹状突起細胞などに吸着し、感染が始まる。A型インフルエンザウイルスは気道上皮細胞表面のシアル酸、コロナウイルスは気道および腸管上皮細胞のアミノペプチダーゼNがレセプター因子であり、宿主細胞へのウイルスの吸着が明らかにされつつある。
(2)侵入 ウイルスの侵入はいろいろな機構によって行われる。宿主側のエンドサイトーシスendocytosis(細胞が細胞外の物質を取り込むプロセスの一つ)という取り込み作用はウイルスの侵入を助ける。また、エンベロープのもつスパイクタンパク質はエンベロープと宿主細胞膜を融合させる機能をもつ。パラミクソウイルスにはFタンパク質というスパイクタンパク質が知られている。さらに、ウイルス粒子全体が直接宿主細胞膜を分断して侵入する場合もある。
(3)脱殻・解体 宿主細胞質中に入ったウイルス解体は、まずエンベロープを失い、カプシドタンパク質が部分的になくなり、ウイルスゲノムが裸出する。しかし、カプシドタンパク質からの脱殻の細胞中の部位は種によって異なる。
(4)ゲノムの複製、素材からの合成 細胞質内に入ったウイルスは侵入前の姿はない。まず、ウイルス自身のゲノムを複製し、そのゲノムからメッセンジャーRNA(mRNA)に転写し、必要なタンパク質をリボゾームで合成させる。この経過も多様性があって種によって相異がある。DNAウイルスの複製は基本的に真核性生物のDNA複製と遺伝子発現の様式と似ている。RNAウイルスの場合は、ゲノムの複製とmRNAの合成は関連しておこる。RNAウイルスの遺伝子発現に共通なことは、リボゾームで、ウイルス特異的mRNAからタンパク質を合成する過程である。
(5)集合(アセンブリassembly) 新たに複製されたゲノムやパビリオン構成タンパク質などが集まり、組み立てられていく。この集合し、組み立てられる場所は、ウイルスゲノムの複製が行われる場所と関係がある。レオウイルスやピコルナウイルスは細胞質で、アデノウイルスやパルボウイルスでは核内で行われる。
(6)成熟・放出(脱出) 新たにできたウイルス粒子が感染性をもつビリオンに完成していくことが成熟である。エンベロープをもつウイルスでは、組立て、成熟、脱出は一体となって進行する。エンベロープをもたないウイルスの多くは感染細胞の崩壊により成熟したウイルス粒子が放出される。
[曽根田正己]
培養細胞にウイルスを感染させた場合、通常、感染細胞はしだいに変化し、最後には死滅する。このような変化を細胞変性効果cytopathic effectといい、他方、腫瘍(しゅよう)ウイルスなどが感染する場合、宿主細胞が異常に増殖をおこす。この場合はトランスフォーメーションtransformationという。この感染細胞は無限に増殖を続ける、不死化とよばれている。
[曽根田正己]
生物一般の特徴は、原核生物も真核生物も、2種類の核酸(DNAとRNA)と細胞構造をもち、遺伝によって固有の体制と代謝を維持し、増殖することである。これに対してウイルスには、体制と核酸(DNAかRNAのいずれか1種類)はあるが、細胞構造がなく、固有の代謝機構が欠けている。ウイルスは宿主細胞外では不活性であり、タバコモザイクウイルス(TMV)のように、罹病(りびょう)植物の絞り汁から結晶として単離されるなどの無生物的特徴をもつところから、ウイルス無生物説がある。しかし、細胞構造のないウイルスにも、宿主細胞の力を借りているとはいえ生物的活性や特徴があり、しかも、形質形成のための遺伝機構が明瞭(めいりょう)である。そのうえ、ウイルスは植物、動物、菌類、細菌のそれぞれの個体の生活や体制と対等な位置を占め重要で密接な関係をもっている。さらに生態学的にも多くの生物の環境における個体数の調整と保持に重要な役割をもつものと考え明瞭に生物の一員とする生物説がある。また生物的にも無生物的にもなりうるものであるので、準生物Subbiota説もある。このほか、一般生物と区別する意味で準生物という説もある。
[曽根田正己]
ウイルスは、核酸とタンパク質を含むということから、遺伝子をもつ最小構造体であるということができる。そこで、生命の起源となった原始形態は、ウイルスに近いものであったといわれていた。しかしその後、ウイルスが結晶化されて無生物説が唱えられ、さらには生物説が現れた。現在でも、ウイルスについては、さまざまな起源説が提唱されている。
ウイルス新生説は、新生物が生ずると新ウイルスが生まれ、宿主の遺伝系とウイルス遺伝系とが一つの機能を営む細胞に溶け合うようになるという考え方であり、細胞起源説は、昆虫とか細菌細胞の核酸の一部が、なにかの原因で制約が外れて細胞体制から離れて独立し、ウイルスになるという考え方である。また、退化細菌説では、ある種の寄生細菌→リケッチア→クラミジア→ウイルスという退化経路を考え、寄生細菌が宿主への依存性(寄生性)を高めるにしたがって、体制が簡単化してウイルスになるというものである。
いずれにしても、ウイルスの起源については、まだ決定的な学説が生まれていないのが現状である。
[曽根田正己]
ウイルスは、その生物的特徴を現すためには、他の生きた生物細胞が必要であり、かつては原始生物の細胞に依存したウイルスのあったことも想像される。原始生物の最初の核酸はRNAであったと考えられるから、RNAウイルスが先に、DNAウイルスが後に現れたということも考えられる。これらのウイルスは細胞に依存して増殖し、遺伝的に変異し、選択によって変遷したはずである。そこには進化があり、系統があったとも考えられる。しかし現在では、系統は近縁のウイルス群のなかの変異を示す程度であって、ウイルス全般の系統についての根拠としては問題にならない。
[曽根田正己]
ウイルスは細胞生物と異なり、系統分類学的体系を考えることはむずかしい。しかし、ウイルス粒子は微細とはいえ、その構造や性状には多様性がある。ウイルス粒子の形態、核酸組成、物理化学的性状、核酸構造などにより分類学的研究が行われ分類体系がつくられている。ウイルスの分類や命名については国際微生物学連合International Union of Microbiological Societies(IUMS)のウイルス学部会のなかに設置されている国際ウイルス分類委員会International Comittee on Taxonomy of Viruses(ICTV)において検討し、継続的に改定勧告が行われている。しかし、研究途上の部分が多く、いまだ流動的な側面をもっている。
ヒトを自然宿主とする動物ウイルスでも、その数は数百種に及ぶとされる。分類の基本は「種species」であり、種と種の類似性、相同性をもってまとめたものが「科family」である。「種」と「科」の間には「亜科」と「属」を置き体系づけている。また、いくつかの科をまとめて「目(もく)」とすることもある。
ウイルスは動物、植物、菌類、細菌など細胞生物のすべてと関係をもち、宿主細胞に感染、その力を借りて増生する。そのため、動物ウイルス、植物ウイルス、菌類ウイルス、細菌ウイルスなど、その研究はそれぞれ独自の発展をしたが、現在は共通の分類体系や命名法によって表現され、研究がされるようになった。
目名、科名、亜科名、属名、種名などの表記については、どの分類群も大文字からはじまるイタリック体の文字を使い、下記の例(単純ヘルペス1型)のように語尾を変えて表記する。
科family:Herpesviridae(~viridae)
亜科subfamily:Alphaherpesvirinae(~virinae)
属genus:Simplexvirus(~virus)
種species:Human herpesvirus
分類基準となるウイルス粒子の性状は次のようである。
(1)形態。ウイルス粒子の大きさ、形態、ペプロマー(スパイク)の有無、ヌクレオカプシドの対象性
(2)物理化学性状。ウイルス粒子の分子量、浮遊密度、沈降定数、pH、熱、陽イオン、有機溶媒、界面活性剤、放射線に対する安定性
(3)核酸。核酸の種類(DNA、RNA)、核酸ゲノムの大きさ、単鎖(1本鎖)か二重鎖(2本鎖)、線状か環状、極性(プラス+かマイナス-)、分節の数と長さ、塩基配列など
(4)タンパク質。構造タンパク質、非構造タンパク質、それぞれの性状
ウイルスはDNAまたはRNAの一方の核酸しかもたないため、増生の基本行動である核酸の複製または転写は、両方の核酸をもつ細胞生物とは異なる。RNAウイルスでは、一般にはRNAからRNAを合成するRNA依存RNAポリメラーゼによって行われる。しかし、レトロウイルス科のRNAウイルスは、RNAからDNAを合成する逆転写酵素をもちDNAを合成し、これによってRNAをつくる。DNAウイルスであるヘバドナウイルス科のウイルス(B型肝炎ウイルスが属する)は逆転写酵素をもつ。DNA and RNA reverse transcribing virusesとよばれる。RNAウイルスの多くは単鎖(1本鎖)であり、二重鎖(2本鎖)は少ない。核酸のゲノム配列が、ウイルスRNAとmRNAが同じ場合と、そうでない場合がある。配列が同じ場合を+(プラス)の極性、相補的な配列の場合を-(マイナス)の極性とよぶ。プラス単鎖RNAをもつウイルスでは、ウイルスが細胞内に侵入したのち、ゲノムRNAがそのままmRNAとして働くため、RNA依存RNAポリメラーゼ(転写酵素)はもたない。マイナス単鎖RNAでは、RNAをmRNAによみかえて転写されねばならないのでウイルス粒子内にポリメラーゼがある。マイナス単鎖RNAウイルスには核酸ゲノムが分節するものと非分節のものがある。分節ゲノムをもつウイルスには、オルトミクソウイルス科(麻疹(ましん)ウイルス、狂犬病ウイルスなどが属する)、アレナウイルス科(ラッサウイルス、リンパ性脈絡髄膜炎ウイルスなど)、ブニヤウイルス科(インフルエンザAウイルス、Bウイルス、Cウイルスなど)、レオウイルス科(レオウイルス、ロタウイルスなど)がある。
ウイルス粒子ビリオンの構造は多様性がある。ヌクレオカプシドの多くは対称型である。正二十面体のヌクレオカプシドをもつウイルス(立方対称)ではカプソマーの数は科によって固有である。RNAウイルスはらせん対称のヌクレオカプシドをもつものが多い。
ウイルスはDNAウイルスとRNAウイルスの2群に大別されるが、ICTVの第7次勧告改定では細胞病原因子を加えて体系づけされている。DNAウイルスは二重鎖、単鎖、DNA転写ウイルスの3群、RNAウイルスは二重鎖、プラス単鎖、マイナス単鎖のRNA3群に分類。微細病原因子はウロイドviroid(おもに植物病原体)と核酸(DNAやRNA)をもたないプリオンprion(おもに牛海綿状脳症の病原体)を加え分類している。
[曽根田正己]
とくにヒトを自然宿主とするウイルス科は次のようである。
(1)DNAウイルス、二重鎖
ポックスウイルス科Poxviridae、ヘルペスウイルス科Herpesviridae、アデノウイルス科Adenoviridae、ポリオーマPolyomaviridae、パピローマウイルス科Papillomaviridae
(2)DNAウイルス、単鎖
サルコウイルス科Circoviridae、パルボウイルス科Parvoviridae
(3)DNAおよびRNA逆転写ウイルス
ヘパドナウイルス科Hepadonaviridae(二本鎖DNA)、レトロウイルス科Retroviridae(プラス鎖RNAゲノム)*
*RNAウイルスであるが逆転写酵素をもつのでこれを優先する。
(4)RNAウイルス(二重鎖)
レオウイルス科Reoviridae
(5)マイナス単鎖RNAウイルス(Mononegaviralesモノネガウイルス目)
ボルナウイルス科Bornaviridae、フィロウイルス科Filoviridae、パラミクソウイルス科Paramyxoviridae、ラブドウイルス科Rhabdoviridae、オルトミクソウイルス科Orthomyxoviridae、ブニヤウイルス科Bunyaviridae、アレナウイルス科Arenaviridae
(6)プラス単鎖RNAウイルス
ピコルナウイルス科Picornaviridae、カリシウイルス科Caliciviridae、アストロウイルス科Astroviridae、ニドウイルス目(Nidovirales)コロナウイルス科(Coronaviridae)、フラビウイルス科(Flaviviridae)、トガウイルス科(Togaviridae)
[曽根田正己]
おもな病原ウイルスならびに科の特徴は次のとおりである。
(1)ポックスウイルス科。ワクチニアウイルス、牛痘ウイルス、痘瘡(とうそう)ウイルス
ビリオンはレンガ状や卵形、300×200×150ナノメートル、カプシドは複雑な形態、エンベロープをもつ、ゲノムは線状、転写酵素をもつ
(2)ヘルペスウイルス科。ヒトヘルペスウイルス1~8型(単純ヘルペスウイルス、水痘・帯状疱疹(ほうしん)ウイルス、サイトメガロウイルス、EBウイルス)
エンベロープあり、ビリオンは球状、120~200ナノメートル、カプシドは立方対称、カプソマー数162、ゲノムは線状
(3)アデノウイルス科。ヒトアデノウイルス(上気道炎、流行性結膜炎、咽頭結膜熱をおこす)
エンベロープなし、ビリオンは正二十面体、70~90ナノメートル、カプシドは立方対称、カプソマー数252、ゲノムは線状
(4)パピローマウイルス科。ヒトパピローマウイルス1~70型以上(皮膚粘膜上皮に乳頭腫(しゅ)をつくる。性器粘膜に尖圭(せんけい)コンジローマをおこす)
エンベロープなし、ビリオンは正二十面体、50~55ナノメートル、カプシドは立方対称、カプソマー数72、ゲノムは環状
(5)ポリオーマウイルス科。JCポリオーマウイルス(進行性多巣性白質脳症をおこす)、BKポリオーマウイルス
ビリオンは正二十面体、40~45ナノメートル、カプシドは立方対称、カプソマー数72、ゲノムは環状
(6)ヘパドナウイルス科。(急性・慢性のB型肝炎をおこす)
ビリオンは球状、40~48ナノメートル、カプシドは立方対称、ゲノムは二重鎖、一部は単鎖で環状、逆転写酵素をもつ。
(7)レトロウイルス科。エイズウイルス(後天性免疫不全症候群をおこす)、HTLV-1ウイルス(成人T細胞白血病をおこす)
エンベロープあり、ビリオンは球状、8~100ナノメートル、カプシドは球状、桿(かん)状。ゲノムは単鎖、プラス鎖、逆転写酵素をもつ。
(8)レオウイルス科。ロタウイルス(嘔吐(おうと)下痢症をおこす)
エンベロープなし、ビリオンは正二十面体、60~80ナノメートル、カプシドは立方対称、カプソマー数32、ゲノムは二重鎖、10~12分節
(9)フィロウイルス科。エボラウイルス(エボラ出血熱をおこす)、マールブルグウイルス(マールブルグ熱をおこす)
エンベロープあり、ビリオンは繊維状、80×490~970ナノメートル、カプシドは螺旋対称、ゲノムは単鎖、マイナス鎖、非分節
(10)パラミクソウイルス科。パラインフルエンザウイルス、麻疹ウイルス、ムンプスウイルス、RSウイルス
エンベロープあり、ビリオンは球状、多形性、150~300ナノメートル、カプシドは螺旋対称、ゲノムは非分節
(11)ラブドウイルス科。狂犬病ウイルス
エンベロープあり、ビリオンは弾丸型、90~180ナノメートル、カプシドは螺旋対称、ゲノムは非分節
(12)オルトミクソウイルス科。インフルエンザウイルスA型、B型、C型(鳥インフルエンザウイルスを含む)
エンベロープあり、ビリオンは球状、80~120ナノメートル、カプシドは螺旋対称、ゲノムは非分節
(13)ブニヤウイルス科。ブニヤウイルス、ハンタウイルス(腎症候性をおこす)、ナイロウイルス(クリミア‐コンゴ出血熱をおこす)、出血熱症、脳炎をともなう症状をおこすウイルス群
エンベロープあり、ビリオンは球状、多形性、80~120ナノメートル、カプシドは螺旋対称、ゲノムは3分節
(14)アレナウイルス科。ラッサウイルス(ラッサ熱をおこす)
エンベロープあり、ビリオンは球状、80~120ナノメートル、カプシドは螺旋対称、ゲノムは2分節
(15)ピコルナウイルス科。ポリオウイルス1~3型、コクサッキーA群(手足口病をおこす)、コクサッキーB群、エコーウイルス、A型肝炎ウイルス、ヒトライノウイルス、ヒトパレコウイルス1型、2型、エンテロウイルス
エンベロープなし、ビリオンは正二十面体、22~30ナノメートル、カプシドは立方対称、カプソマー数32
(16)カリシウイルス科。ノロウイルス(胃腸炎をおこす)、サポウイルス
エンベロープなし、ビリオンは正二十面体、27~40ナノメートル(小型)、カプシドは立方対称、カプソマー数32
(17)アストロウイルス科。アストロウイルス(乳幼児胃腸炎をおこす)
エンベロープなし、ビリオンは正二十面体型、カプシドは立方対称
(18)コロナウイルス科。ヒトコロナウイルス、SARSコロナウイルス
エンベロープあり、ビリオンは球状、120~160ナノメートル、カプシドは螺旋対称
(19)フラビウイルス科。日本脳炎ウイルス、黄熱ウイルス、デングウイルス(デング熱をおこす)、C型肝炎ウイルス
エンベロープあり、ビリオンは球状、40~60ナノメートル、カプシドは立方対称
(20)トガウイルス科。アルファーウイルス(脳炎ウイルス)、ルピウイルス(風疹ウイルス)
エンベロープあり、ビリオンは球状、60~70ナノメートル、カプシドは立方対称、カプソマー数は32または42()。
(21)プリオン(牛海綿状脳症をおこす)。ウイルスより小さい病原因子。核酸を欠き、タンパク質により構成される。
[曽根田正己]
ウイルスはもっとも微小な感染性因子で、種類も多く、その感染によっておこされる病気も多い。ウイルスは一般に感染する宿主によって動物ウイルス、植物ウイルス、細菌ウイルスに3大別される。動物ウイルスは、人以外の動物では腫瘍を形成することもかなり知られているが、人に対しては多くの感染症をおこすことが確認されているだけで、腫瘍については未確認である。
人とのかかわりでみた場合、ウイルスは宿主の個体、臓器、組織に対して、かなりえり好みをして病気をおこすということができる。たとえば、日本脳炎ウイルスが感染した場合、大部分の人は発病しない(不顕性感染)か、発病しても軽症ですみ、ごく少数の人が重症となる(顕性感染)。これは個体に対するウイルスのえり好みの顕著な例であるが、逆にインフルエンザのように、感染すればほとんど100%顕性感染をおこすウイルスもある。
次に臓器・組織に対するウイルスのえり好みについて述べる。
(1)呼吸器系の臓器を好んで侵すウイルス。ライノウイルス、アデノウイルス、コロナウイルス、コクサッキーウイルスなどで、鼻炎、咽頭(いんとう)炎、気管支炎、肺炎などをおこす。
(2)中枢神経系を好んで侵すウイルス。ポリオウイルス、コクサッキーウイルス、エコーウイルス、アデノウイルス、麻疹ウイルス、日本脳炎ウイルス、狂犬病ウイルスなどで、急性灰白髄炎、髄膜炎、脳炎、狂犬病などの病気をおこす。
(3)目を好んで侵すウイルス。アデノウイルスの8型やエンテロウイルスの70型で、流行性角膜結膜炎や、急性出血性結膜炎などをおこす。
(4)消化器系の臓器を好んで侵すウイルス。コクサッキーAおよびB群ウイルス、あるいは単純ヘルペスウイルスが口内炎をおこし、ロタウイルス、ノロウイルス、カリシウイルスなどは幼児に下痢症をおこす。
(5)皮膚に著明な病変をおこすウイルス。病名として麻疹、風疹、水痘などがあり、いずれも病名を冠したウイルスが病原である。
発病・経過の観点からウイルスによる病気を分類すると、急性感染症、持続性感染症、遅発性感染症に3大別される。
(1)急性感染症 麻疹、水痘、ポリオ、インフルエンザなどが含まれるが、これらの病気は臨床的にもそれぞれ特異な症状を呈し、病原ウイルスも別々で、病原体と病気が1対1で対応している。しかし、急性感染症のなかでも、鼻かぜという臨床症状をおこすウイルスには、ライノウイルスやコロナウイルスなどがあり、乳幼児の下痢症をおこすウイルスにはロタウイルス、ノロウイルス、カリシウイルス、アストロウイルスがあるなど、複数の病原体が一つの病気に対応するものもある。
(2)持続性感染症 前述の急性感染症では、症状が回復すれば病原ウイルスは体内から消滅するのが常である。ところが、これが体内の一部に残存し、ときに再発することがある。たとえば、単純ヘルペスに幼児期に感染しても症状を呈することなく経過し、やがて宿主の抵抗力が低下したときに、口唇ヘルペスとして水疱が生じることがある。これは、三叉(さんさ)神経節などにウイルスが潜伏していたわけであり、水疱液中にウイルスを証明することができる。これを持続性感染症という。
(3)遅発性感染症 数か月から数年という長い潜伏期ののちに発病し、遷延(遅延)性の経過をとり、予後の悪い感染症をいう。亜急性硬化性全脳炎や、進行性多巣性白質脳症がその例である。
ウイルス病(ウイルス感染症)の伝染経路には、細菌やリケッチアなどによる感染症と同様に、飛沫(ひまつ)感染、経口感染、接触感染、昆虫媒介感染などがあるほか、母から子へ伝わる子宮内感染、経胎盤感染(B型肝炎ウイルス)、産道感染(サイトメガロウイルス)がある。
ウイルス病を治療と予防の面からみると、ウイルス病には、細菌やリケッチアに対する抗生物質のように効く抗ウイルス剤はなく、わずかに免疫グロブリン製剤を潜伏期の初期に用いれば有効なものがある程度である。また、インターフェロンの抗ウイルス作用が注目され治療薬が開発されているが、ウイルス病には、ワクチンによる予防(ポリオ、麻疹、風疹、流行性耳下腺(じかせん)炎、種痘、日本脳炎、インフルエンザなど)が現在のところ最高の対策である。発病後の治療としては、対症療法と合併症予防の治療が行われるにすぎない。
[柳下徳雄]
ウイルスの遺伝学上の性状について、その概略のいくつかを述べる。
[寺川博典]
ファージ(バクテリオファージまたは細菌ウイルス)によって感染された大腸菌は、やがてファージを放出するが、それは新合成された酵素が細胞壁を部分的に分解して宿主を溶菌することによっておこる。溶菌とは、ウイルスの酵素タンパク質の働きによって細胞が溶ける現象のことである。実験的には、寒天培地上の細菌の集落(コロニー)に溶菌がおこると、ところどころに丸く透き通った斑点(はんてん)(溶菌斑)として認められる。この溶菌斑の形や大きさには特徴があり、ファージがもつ遺伝性質の表現型と考えられている。
ところが、宿主に侵入したファージDNAが、こうした溶菌の過程へ進まないで、宿主DNAの一定部位に定着し、そのまま複製されて宿主の細胞分裂と同調していくことがある。この状態にある、もとのファージDNAをプロファージとよび、宿主を溶原菌という。この溶原菌は、ビリオン(感染性をもつウイルス粒子)の生産能力を潜在的にもっていて、それを子孫に伝えていく。やがて、この潜在能力の「誘発」によってプロファージが細胞から遊離して活性となると、宿主は溶菌される。こうして放出されたビリオンは、プロファージのない同種細菌に感染し、その細菌を溶菌、または溶原菌に変えていく(溶原化)。こうしたファージの溶菌、溶原化、誘発の仕組みは遺伝学的に説明づけられている。
[寺川博典]
溶菌斑や宿主の範囲(宿主域)、あるいは温度感受性などにかかわるウイルスの遺伝子に、突然変異が生じて変異株ができることがある(もとのウイルスの系統は野生株という)。たとえば、大腸菌を宿主とするT系ファージの一つであるT4による溶菌斑には、野生株による野生型溶菌斑とは大きさの異なるr型が出現する。これは、T4ファージに含まれる別の三つの遺伝子の突然変異によっておこるものであり、r型の出現は、大腸菌の株によって異なっている。
また、野生株のT2ファージは、大腸菌のある株を溶菌しないが、変異株であるT2hは溶菌斑を生ずる。さらに別の変異株T2hrの溶菌斑の形はT2hとは異なる。このT2hrとT2(変異株との比較表記ではT2h-r-)が重感染すると、子ファージには親と同じもののほかに、T2hとT2rが一定比で生ずる。これは、親DNAの断片の結合(組換え)によると考えられている。
このT系ファージの研究によって、突然変異の仕組みや遺伝暗号、あるいは遺伝子とタンパク質構造との関係といった、生物学の基礎的問題が解かれてきたわけである。
[寺川博典]
動物ウイルスが宿主に感染した際に現れる重要なものとして、干渉因子(インターフェロン、IFN)、抗体などがある。インターフェロンは糖を含むタンパク質で、重感染の場合には、一方のウイルスの増生を阻止する働きをもっている。抗体はウイルスタンパク質が抗原となって生成されるもので、次のウイルスの吸着、ないしは増生を妨げる作用をもつ。このような抗原性、インターフェロン感受性、増殖能、病原性、宿主域などは、動物ウイルスの遺伝指標として重要なものである。
インフルエンザウイルスはS抗原(核タンパク質のこと。核酸とタンパク質の複合体)によって、A、B、C型に分類されるが、A型は表面のV抗原(糖タンパク質)によってA、A1、A2、A3などがあり、変異しやすいのも、ウイルスのもつ遺伝的性状がかわるためである。
[寺川博典]
『岡田吉美著『UPバイオロジー24 植物ウイルスと分子生物学』改訂第2版(1989・東京大学出版会)』▽『石浜明・永井美之他編『生命科学を推進する分子ウイルス学』(1995・共立出版)』▽『畑中正一編『ウイルス学』(1997・朝倉書店)』▽『児玉浩憲著『図解雑学 ウイルス』(1998・ナツメ社)』▽『大里外誉郎編『医科ウイルス学』改訂第2版(2000・南江堂)』▽『吉開泰信編『ウイルス・細菌と感染症がわかる』(2004・羊土社)』▽『山内一也著『ウイルスと人間』(2005・岩波書店)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponsored by ![]()
ビールス,バイラスなどとも発音される。核酸(DNAもしくはRNA)とタンパク質からなる,細菌よりも小さな一群の病原体。遺伝情報を担う核酸がタンパク質の外被におおわれた構造をもち,それぞれのウイルスに特有の宿主となる細菌や生物の細胞に寄生して,宿主のタンパク質合成能やエネルギーを利用して,自己増殖を行う。ウイルスは,その大きさが数十~数百nmときわめて小さく単純であることと,単独では生物としての要件である自己増殖能をもたず,寄生してはじめて自己増殖を行うことから,しばしば〈生物と無生物の間にあるもの〉と表現されることがある。また,ワクチンによる免疫療法を除くと,細菌などに対する抗生物質のように,ウイルス感染症に対する直接的な特効薬がいまだに発見されていないのも,ウイルスの独特な寄生性と自己増殖性によるものである。
19世紀に入って,L.パスツールやR.コッホたちの研究により,伝染病の多くが病原性細菌によってひき起こされていることが明らかにされた。しかし,伝染病のうちには,これらの病原性細菌が見いだされないものがあった。1892年にイワノフスキーD.I.Ivanovskiiがタバコモザイク病で,98-99年にレフラーF.LoefflerとフロッシュP.Froschがウシの口蹄疫で,それぞれ,細菌を通さない細菌ろ過器を通過させたろ液それ自体が,明確に病原性をもつことを示した。引き続いて,黄熱,狂犬病,牛痘などの伝染病の病原が,同じように細菌ろ過器を通過することが確認され,20世紀の初頭には〈ろ過性病原体filterable virus〉という概念が確立された。ウイルスという言葉はもともとはラテン語の〈毒素〉という意味で,一般に伝染病の病原因子を総称していたが,1930年代以降は,ろ過性病原体を意味するようになった。
コッホによって確立された細菌学の特徴的な手法は,培地による細菌の培養であった。初期のウイルス研究においても,さまざまな培地でウイルスを培養する試みがなされ,30年代が最も盛んであった。しかし,このような試みは結局失敗に終わった。ウイルスを分離培養するためには,適切な生きた細胞が必要とされている。現在,ウイルスの分離培養に用いられている手段は以下のようである。(1)実験動物による培養,(2)発育鶏卵(孵化(ふか)の途中の受精卵)による培養,(3)培養細胞による培養。しかし現在においても,適切な培養手段が見いだせないために研究の進んでいないウイルスも存在している。
30年代までのウイルス学は,培地による培養という細菌学の手法を適用できないという困難をかかえたまま,病原性細菌学の一部として取り扱われていた。しかしこの時期以降,ウイルス学は現在の分子生物学の発展の基盤を形づくるようになってくる。35年スタンリーW.M.Stanleyはタバコモザイクウイルスの結晶化に成功したが,このことは,結晶化したウイルスが生物か無生物かという論議をひき起こした。この問いに答えることは今日でも難しいが,スタンリーの研究は,遺伝情報の担い手である遺伝子の化学的本体の追求へと引き継がれていった。遺伝子の本体がDNAであることがはっきり証明されたのは,52年のハーシェーA.D.HersheyとチェースM.Chaseによる大腸菌に感染するT2ファージを用いた実験によってであった。そして53年のJ.D.ワトソンとF.H.C.クリックによるDNAの二重らせん構造の解明を契機として,分子生物学の飛躍的な発展がもたらされる。50年代に取り扱われたウイルスはバクテリオファージが中心であったが,60年代以降,これらの分子生物学の知見と,培養細胞によるウイルスの培養方法の確立とともに,動物ウイルスにも研究の目が注がれてきている。今日,ウイルス学が取り扱う範囲は,ウイルス自体についての形態形式,遺伝子構造と遺伝子の機能発現などだけにとどまらず,宿主細胞の側での遺伝子の構造と機能,発癌やウイルスの病原性を定めている遺伝子とその機能,ウイルスに対する防御機構などの研究にも及んでいる。
自己増殖していくための遺伝情報は,ウイルスにおいてもまた核酸によって担われているが,高等生物の遺伝情報はDNAに限られるのに対して,ウイルスの場合にはRNAのときもある。ウイルス自体の大きさが微小なため,遺伝子としてのウイルスの核酸のもつ情報量は制限されており,核酸の情報に基づいて生み出されるタンパク質の種類も限られている。このような制限のもとでつくり出されてくるウイルス粒子は,基本的には,単位的な要素(サブユニット)の規則的な繰返しによって構成されている。サブユニットは多くの場合,同一のタンパク質の集合したものであるが,数種類のタンパク質からなる複雑な集合体である場合もある。
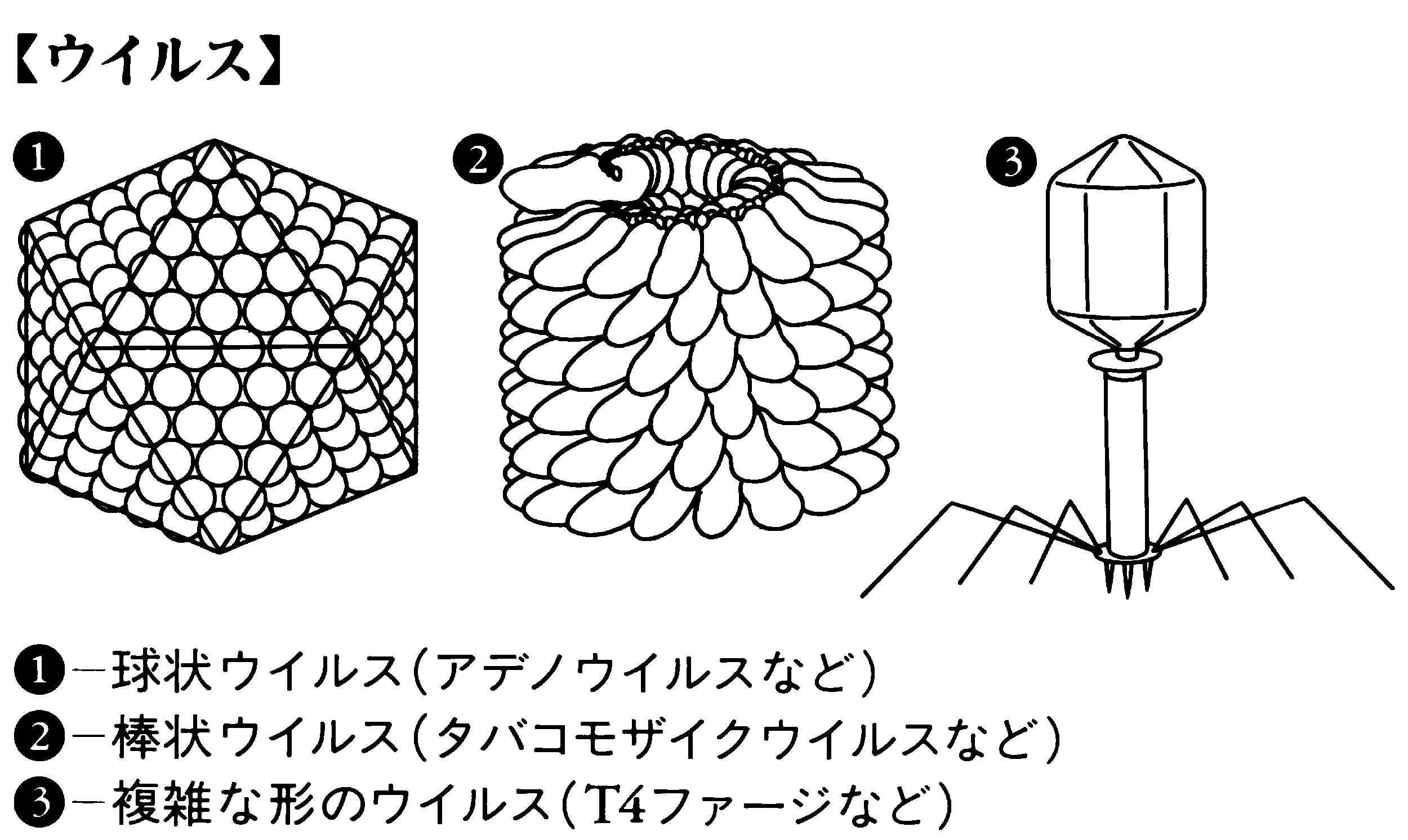
ウイルスの基本的な形態は,遺伝子である核酸をタンパク殻(カプシドcapsid)でおおっている形である。ウイルスカプシドを構成しているタンパク質サブユニットの配列方法は,正二十面体からせん構造になっている。限られた種類のサブユニットからカプシドをつくると,正二十面体からせん構造になるのである。したがって,電子顕微鏡でウイルスを拡大して見た場合,ウイルス粒子の形状は正二十面体を反映した球状,らせん構造を反映した棒状,およびこれらの形の複合した複雑な形の三つに大別されることが観察されている。複雑な形のウイルス(T4ファージなど)では,ウイルス粒子を構成しているタンパク質の種類数も多くなっており,サブユニットの種類数も多くなっている。
ある種のウイルスでは,カプシドの外側にエンベロープenvelopeと呼ばれる膜様構造物をもっている。エンベロープは,タンパク質,脂肪,炭水化物などを含んでいる。
ウイルスは,それが感染する宿主の種類によって,動物ウイルス,植物ウイルス,昆虫ウイルス,細菌ウイルス(バクテリオファージ)の四つに大別される。
1960年代以降の電子顕微鏡やX線回折によるウイルス形態学の進歩および分子生物学の発展によって,ウイルスの一元的な分類体系がつくられつつある。分類の基準となるものは,ウイルスのもつ核酸がDNAかRNAか,カプシドの形状,エンベロープの有無,ウイルス粒子が形成される宿主細胞中での位置などである。
人体に侵入したウイルスは,さまざまな病気を引き起こす。ウイルス性の疾患には,胃腸炎,肝炎,出血熱,腫瘍,髄膜炎,肺炎,皮膚疾患などが知られている。
ウイルス性肝炎は,肝炎ウイルスの感染による。肝炎ウイルスはいろいろなウイルスの総称で,A~E型までの,少なくとも5種類の肝炎ウイルスが確認されている。A,C,D,E型は核酸がRNAのRNAウイルスで,B型は核酸がDNAのDNAウイルスである。ウイルス性出血熱には,黄熱,デング熱,ラッサ熱,エボラ出血熱,マールブルグ病などがあり,これらの病原体はRNAウイルスである。ラッサ熱,エボラ出血熱,マールブルグ病の三つは,予防法や治療法が確立されていず,致命率が高く,伝染力も強いことで知られている。ウイルス性腫瘍には,Epstein-Barrウイルス(EBV)によるバーキットリンパ腫と鼻咽頭癌,ヒト白血病ウイルス(HTLV)による成人T細胞白血病が知られている。EBVはDNAウイルスであり,HTLVはRNAウイルスである。
エイズ(後天性免疫不全症候群)はエイズウイルス(ヒト免疫不全ウイルス,HIV)の感染による。細胞性免疫不全により,日和見感染症,カポジ肉腫,リンパ腫などが引き起こされる。エイズウイルスはRNAウイルスで,HTLVとともにレトロウイルスと呼ばれるRNAウイルスの仲間である。
→癌
ウイルスが感染した細胞では,細胞内でウイルス核酸の情報に基づいて,タンパク質と核酸の合成が行われ,その結果,新たに数多くのウイルスが出現することになる。この過程が繰り返されていくと,生体の側には,そのウイルスに特有の感染症状が現れる。ウイルスに対して生体側がとる防御方法は,インターフェロン系と免疫系の働きによる2種類がある。インターフェロン系の働きは,ウイルス感染の初期にインターフェロンを産生し,生体の細胞を抗ウイルス状態にさせ,ウイルスの細胞内での増殖を抑えることにある。免疫系の働きには,抗体がつくられる液性免疫と,免疫細胞が働く細胞性免疫の2種類がある。液性の免疫系では,ウイルス粒子が非自己の物質として生体の免疫系に認識され,そのウイルスに特異的な抗体がつくり出される。抗体はウイルスと結合して,ウイルスの感染力を失わせる。細胞性の免疫系では,ウイルスが感染したことによって本来の機能を失った細胞を免疫細胞が攻撃することによって,それらの細胞を生体からとり除く。
→バクテリオファージ
執筆者:川口 啓明
植物に感染するウイルスを植物ウイルスと呼び,おもに高等植物に病気を起こすものを指すが,菌類(キノコを含む)や藻類に感染するものもある。植物ウイルスは形,大きさ,宿主範囲と病徴,伝染方法,核酸の性状などに基づき約600種が同定されているが,それらは約30のグループに分類される。ウイルスによって起こる病徴はウイルスの種類によりさまざまであるが,最もふつうにみられるのは葉の緑色に濃淡のまだらが生じるモザイクで,葉脈に沿うこともあり,単子葉植物では縞になることが少なくない。ほかに,黄化,壊死(えし),輪紋,葉巻,萎縮,奇形などがある。これらの病徴は葉,茎,花,果実の各器官に現れるが,根は通常発達が悪くなる。果樹や樹木では,はっきりした病徴が認められないことも多い。植物体内では動物と異なり抗体をつくる機能がないのでウイルスに対する免疫ができず,一般にウイルスは植物が枯れるまで残存する。ウイルスが増殖する細胞・組織はウイルスによって異なるが,どのウイルスも生長点付近のごく一部を除いて植物の全身に分布するため,株分け,挿芽,球根など栄養繁殖する植物では,母株のウイルスはそのまま苗に伝達される。接木をすると組織の癒合によりウイルスは必ず伝染する。一方,種子伝染性のウイルスは,マメ類のウイルス病やセンチュウ媒介のウイルス病など一部にみられる程度で,伝染率も平均してそう高くない。
野外における最も重要な伝染方法は昆虫による媒介(虫媒伝染ともいう)である。アブラムシ,ヨコバイ,ウンカ類は口針で植物汁液を吸収して害を与えるだけでなく,吸汁の際にウイルスを伝染する機会が多い。畑作物,野菜,花のウイルス病の多くはアブラムシで媒介される。キュウリモザイクウイルス,ジャガイモYウイルスなどはアブラムシの短時間の吸汁で伝染するが,虫体内でウイルスが保持される時間は短い。イネのウイルス病にはヨコバイまたはウンカで媒介されるものが多く,イネ萎縮病ではヨコバイが病植物を吸汁するとウイルスが昆虫細胞内で増殖し,その虫は終生伝染を続ける。さらに,雌虫が保毒すると卵を通して子虫へ高率にウイルスが伝染するが,これを経卵伝染という。その他,アザミウマ,コナジラミ,コナカイガラムシ,ハムシなどの昆虫やダニによって媒介されるウイルスがある。センチュウや遊走子をもつ下等菌類によって媒介されるウイルスは,それらが生息する土壌を通して伝染する。実験的には多数のウイルスが汁液で伝染するが,農作業の際などに接触して伝染するウイルスはタバコモザイクウイルスなど一部にすぎない。ウイルス病の防除には伝染を断つことが最も肝要であるが,組織培養による無病苗の生産も実効を上げており,また一部では病原性の弱い変異ウイルスの干渉効果を利用した方法も行われる。
執筆者:奥田 誠一
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponsored by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponsored by ![]()
ビールスともいう.ウイルスはその粒子の大きさが20~30 nm の間にあり,細菌が通れない濾過器を通過する病原体と定義される.この粒子は核酸を中心としてタンパク質の殻(coat protein)で包まれている.そのほか,少量の脂質,炭水化物が含まれるものもある.核酸の量はタンパク質の量に比べて非常に少ないが,ウイルスの核酸はDNAとRNAのどちらかを遺伝物質としてもっており,この核酸が増殖に重要な役割を果たす.これはウイルスから単離した核酸部分だけでウイルス病が起こることによって示された.ウイルスはそれ自体では代謝系をもたず,宿主細胞に感染してはじめて自己増殖を行う.ウイルスは感染する宿主細胞によって,動物ウイルス,植物ウイルス,および細菌ウイルスの3種類に分類される.細菌ウイルスはバクテリオファージともよばれている.
出典 森北出版「化学辞典(第2版)」化学辞典 第2版について 情報
Sponsored by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponsored by ![]()
出典 教育ネットワーク情報セキュリティ推進委員会(ISEN)情報セキュリティ用語辞典について 情報
Sponsored by ![]()
Sponsored by ![]()
Sponsored by ![]()
出典 (株)ジェリコ・コンサルティングDBM用語辞典について 情報
Sponsored by ![]()
Sponsored by ![]()
…細菌類,菌類などは,主として細胞の形態学的特徴などから,かつては植物として扱われてきたが,もっぱら有機物質に頼るという生き方などは植物的とはいえず,さりとて全体的な様相は動物的でもなく,進化の筋道からみても,動物,植物のいずれかに含めるのは難があるとして,今日では〈第三の生物〉といわれる菌類(菌界Mycota)なるグループが設定されている。 ウイルスが生物であるかどうかは長い間論議の的であったが,ウイルスは自分ではエネルギー転換系をもたず,物質代謝の面は生物の生きた細胞に依存して自己増殖を行うため,現在では生物の範疇(はんちゆう)には含められていない。もともとは生物であったのかもしれないが,他の生物の細胞に〈寄生〉して生きるようになったため,情報のシステムだけが残ってしまったものと考えてもよいだろう。…
…ウイルスのうち細菌に感染して増殖するものの総称。略してファージともいう。…
…細菌を宿主とするウイルス(バクテリオファージ,単にファージともいう)には,感染後,細胞内で増殖して宿主を死に至らしめるものと細胞内に潜伏してしまうものがある。後者の場合,ウイルスの遺伝物質(ゲノム)は宿主染色体の一部として組み込まれており,その発現がリプレッサーと呼ばれるタンパク質の働きで大部分抑制されている。…
※「ウイルス」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponsored by ![]()
夏の暑さに体が慣れること。数日から数十日間で起こる短期暑熱順化と、数年または数世代にかけて起こる長期暑熱順化とがある。→寒冷順化[補説]近年では、冷房設備の普及にともない短期暑熱順化が起こりにくくなっ...