
精選版 日本国語大辞典 「貝類」の意味・読み・例文・類語
かい‐るいかひ‥【貝類】
- 〘 名詞 〙 貝のたぐい。
- [初出の実例]「土よりしたにゐる海にすむ、かいるいのたぐひを名につかしったがござる」(出典:咄本・正直咄大鑑(1687)青)
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponsored by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponsored by ![]()
軟体動物門Molluscaのうち、貝殻をもつものの総称。そのなかには、単板綱、多板綱、腹足綱の前鰓(ぜんさい)亜綱と有肺亜綱の大部分、および後鰓亜綱の一部、掘足綱(くっそくこう)、二枚貝綱、ならびに頭足綱のごく一部が含まれる。しかし、本項では、貝殻の有無にかかわらず、広く軟体動物門全体について述べる。
[奥谷喬司]
軟体動物は無脊椎(むせきつい)動物二十数門のなかで、たとえば触手動物門のように多様な形態をもつグループと比べればよくまとまっている門の一つではあるが、現在認められている7綱の各綱ともその生活様式によく適応し、きわめて変化に富む形態をしている。したがって各綱に共通する特徴を引き出すのはむしろ困難であるといえるが、想像される祖先型から考えられる軟体動物の特徴は次のように集約されよう。
体は柔らかく伸び縮み自在で、体節構造はなく一続きの筋肉で覆われ、ほぼ頭部と足部および内臓塊の3部分に区別され、背面を覆う外套膜(がいとうまく)から分泌される炭酸カルシウムを基質とする貝殻をもつ。体は左右相称で、頭部には目、触角および口があり、口には顎板(がくばん)と歯舌(しぜつ)がある。足は広く平らな筋肉質で、匍匐(ほふく)に適している。外套膜と内臓塊との間にできる空所は外套腔(こう)とよばれ、そこには1対の羽状本鰓があり、排出口や生殖開口もある。消化管は単純で、嗉嚢(そのう)、胃、腸からなり、大きな中腸腺(せん)が付属し、肛門(こうもん)は後端に開く。心臓はえらの根元にあって1心室2心耳、血管は開放系で血色素はヘモシアニン系である。囲心腔が真の体腔で、腎臓(じんぞう)や生殖腺と連絡している。性は分かれていて、卵生。卵は粘液質の卵嚢物質で包まれて産み出される。卵は全割卵で、螺旋(らせん)状不等分割の途中で軟体十字molluscan crossを現し、幼生はトロコフォラtrochophora(担輪子)期およびベリジャーveliger(被面子)期を通り、浮遊生活から底生生活へ移る。
分類は研究者によって多少の相違はあるが次の7綱とするのが普通で、(1)と(2)を双神経亜門Amphineura、(3)~(7)を貝殻亜門Conchiferaとまとめることができる。
[奥谷喬司]
Aplacophora 体は細長いミミズ状で、触角、目はない。背面は外套膜で覆われて外側は厚いクチクラ層を形成し、その中に多数の微小な骨片を含んでいる。腹面は足になっているが、足裏は細い縦溝になっていて匍匐の用をなさない。口には微細な歯舌をもつ。溝腹目Solenogastresと尾腔目Caudofoveataに2大別され、前者は腔腸(こうちょう)動物のヤギ類などに絡んで生活し、後者は海底質中にすむ。世界に現生約200種、日本近海に約15種が知られる。
[奥谷喬司]
Polyplacophora いわゆるヒザラガイ類である。体は背腹に著しく扁圧(へんあつ)され、輪郭は楕円(だえん)形をしている。背中線上に8枚に分かれた殻板が瓦(かわら)状に配列しているが、なかには間隔があいて飛び石状に配列するものや筋肉中に埋没している種もある。最前端と最後端のものはそれぞれ頭板および尾板とよばれ露出部分は半月形で、ほかの6枚すなわち中間板とは形態が異なる。殻板列の前後左右は硬い肉帯girdleで取り巻かれ、肉帯の上に鱗(うろこ)や小棘(しょうきょく)、針などの装飾がある。頭部には触角、目を欠くが、頭板の上には光を感じる神経の末端がある。口にはよく発達した歯舌があり、歯舌歯が磁鉄鉱のキャップをかぶっているのが特徴である。足裏は広く楕円形の吸盤状で、その左右を走る狭い溝状の外套腔に多数のえらが並んでいる。神経は梯子(はしご)状構造で、肛門は後位である。現生約1000種、日本産90種。
[奥谷喬司]
Monoplacophora 殻は小さく単純な笠(かさ)形で、殻頂は前寄りにある。頭部触角や目を欠くが、口の両側にはひげ状の肉片がある。足裏は丸い吸盤状。えらは5対または6対あり、殻につく筋肉は8対あって、神経、排出器などの配列は体節構造の名残(なごり)を示している。この仲間は、1952年に中央アメリカのコスタリカ太平洋側沖の深海から初めてネオピリナ(ガラテアガイ)Neopilina galatheaeが採集されるまで、古生代のカンブリア紀後期からデボン紀中期に栄えて以後絶滅したものと思われていた。初採集以来、中央太平洋に2種、東太平洋に7種、南大洋(太平洋、インド洋、大西洋の三大洋南部が一続きとなった海域)に2種、北大西洋に1種の合計12種が知られている。そのうち例外的に水深300~400メートルの浅海から知られる1種を除いて、2000~6000メートルの深海底から知られている。日本近海を含む北西太平洋からは、まだ1種も知られていない。
[奥谷喬司]
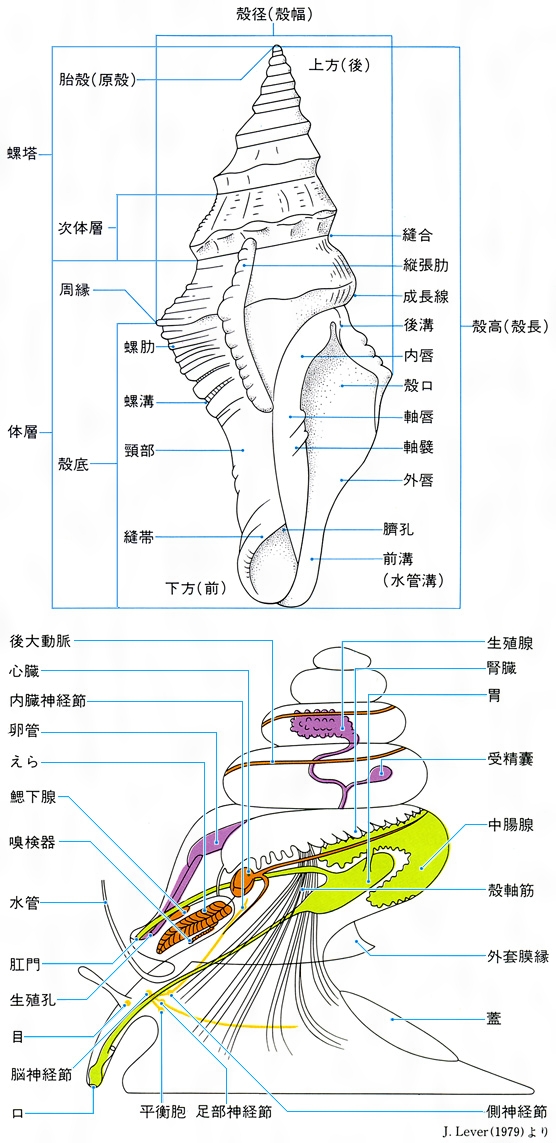
Gastropoda いわゆる巻き貝類で、一つの軸の周りに巻いた螺管からなる殻をもつ。殻は、殻頂から貝の成長の方向に向かって見たとき、時計回りならば右巻き、反時計回りならば左巻きである。圧倒的に右巻きのものが多いなかで、海産ならばキリオレガイ科Triphoriidae、陸産ならばキセルガイ科Clausiliidaeはともに科内の全種が左巻きであるほか、右巻きが主流で、なかに左巻きが混じる科もあり、奇形として右巻きの種内に左巻きの個体が出ることもある。また、ごく少数のものは超右巻きとよばれるが、それらは通常の貝のように巻き下がらず、巻き上がった右巻きのため、見かけは左巻きの殻と右巻きの特徴を有する軟体部とをもつ。巻き貝の殻は円錐(えんすい)形、紡錘形、塔形、球形、平巻きなど変化に富み、棘(とげ)やひれ状の突起をもつものや、巻き方が浅いため耳形や皿形となっているものがある。蓋(ふた)は足の後背部から分泌され、通常は角質ないし革質であるが、石灰質を沈着させているものもある。蓋には殻の巻き方とは逆の螺旋がある。蓋を欠くものも多いが、それらもベリジャー期には蓋が生じ、のちに消失したものである。一方、軟体の腹面が平らな筋肉質の足になっていて、足裏から粘液を分泌しその上をはう。頭部には1対の触角と目がある。内臓は完全に殻の中に収まっているのが普通である。
前鰓亜綱Prosobranchiaでは、発生の途中で内臓の部分が180度ねじれ、足の部分を1階に例えるならば、2階の内臓部分が1階部分と反対のほうを向くことになる。このねじれのため、成体期には、えらは心臓の前に位置し、肛門も前方すなわち口と同じほうに向かって開く。さらに、本来は左右2本が平行に走っていた梯子状の神経もねじれ、8の字形となる。後鰓亜綱Opisthobranchiaではふたたびねじれ戻り、神経は正常な配列となって、えらは心臓より後ろへ移り、肛門も体の後方へ向かって開く。後鰓類では成体期に殻があるのはむしろまれで、ベリジャー期に生ずる巻いた殻や蓋も成体期には失われるものが多い。成体期まで残る殻も薄く軽いものが多く、体内に埋没しているものもある。有肺亜綱Pulmonataは陸上生活者が多く、生活に適応してえらは失われ、かわりに外套腔に血管が集まり肺を形成する。陸上性のものでは卵は堅い殻に入れて産み出され、トロコフォラ期、ベリジャー期を経ず親の縮小型として孵化(ふか)する直達発生(ちょくたつはっせい)をする。
前鰓亜綱は雌雄異体で、体外受精するものと交尾で受精するものとがあり、性転換するものが知られている。後鰓亜綱と有肺亜綱はほとんどが雌雄同体で、体外受精するものはない。腹足綱は軟体動物のうちで現生種がもっとも多く、世界で約8万5000種、日本産は約4500種と見積もられている。
[奥谷喬司]
Scaphopoda いわゆるツノガイ類である。殻は先太りの角笛に似た管状で、普通は弓なりに湾曲し、殻表には縦肋(じゅうろく)が走っている。殻の断面は円形のものが多いが、なかには六~八角形や卵円形、あるいは台形の種もある。殻の太いほうの端すなわち殻口から太い足を出し、それで底質中に潜入する。頭部には目も触角もないが、頭糸という感覚糸の束があり、これで底質中の微生物を口に運ぶ。歯舌は簡単であるが強力。世界の大洋におよそ500の現生種があり、日本近海には70種ぐらいがすむ。小形種は殻長1センチメートル以下であるが、大形のものは10センチメートルを超える。
[奥谷喬司]
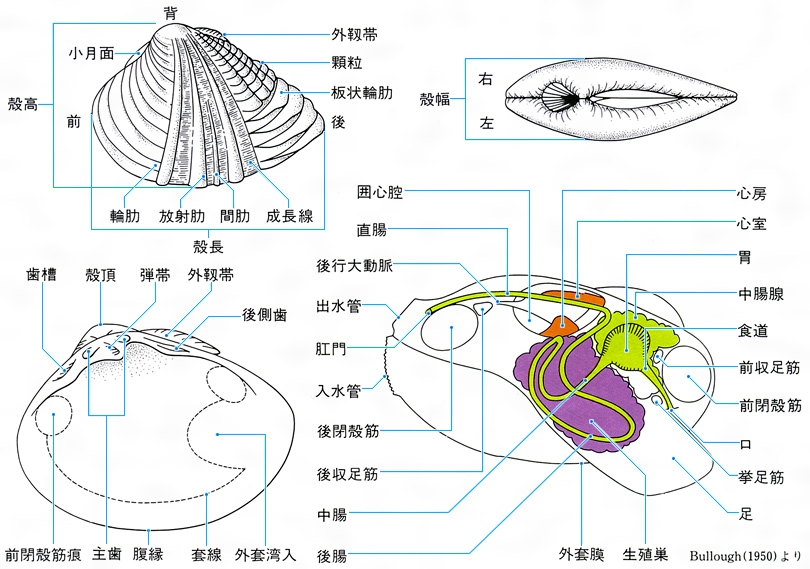
Bivalvia 体は左右相称の皿形の殻片で包まれている。左右の殻は靭帯(じんたい)で連結され、殻頂に近いところで蝶番(ちょうつがい)(鉸歯(こうし))によってかみ合わさっている。ハマグリ形を基本とすると、前後や背腹に延長した細長いものや、片側の殻で地物に固着するために左右が不ぞろいになるものなどがある。殻の表面には通常は成長線があるが、それが持ち上がって成長脈や肋となるもの、棘や鱗片(りんぺん)状などの装飾的な彫刻をもつものなど多様である。靭帯も殻の外にあるものや鉸歯の間に内在するものなど変化に富み、同時に蝶番の歯の配列や数などの形式は分類の重要な標徴となっている。
軟体部には頭部を欠き、したがって目、触角もなく、軟体動物門で口に歯舌を欠く唯一の綱である。体全体を覆う外套膜は、ちょうど殻の内面を裏打ちするようなかっこうとなり、そこから殻質を分泌する。また、外套膜の縁には触角や目がある種がある。内臓の塊は筋肉に包まれ、その腹側に斧(おの)形の足がある。二枚貝綱を斧足綱Pelecypodaとよぶことがあるのはこのためである。内臓塊から一続きの足と外套膜の間の空所は広い外套腔で、ここに左右2枚ずつの広いえらがあり、えらの形式によって原鰓類Protobranchia、糸鰓類Filibranchia、真弁鰓類Eulamellibranchia、隔鰓類Septibranchiaなどに分類される。体の後方で外套膜が癒合し、外套膜内に水を出入させる水管を形成している群がある。二枚貝は外界から呼吸水を取り入れ、えらによって呼吸をすると同時に摂餌(せつじ)する。消化管には複雑な中腸腺が発達している。神経節は集中的で、内臓や足部に大きい神経節を配している。雌雄同体や性転換するものがあるが、多くは雌雄異体で体外受精をする。一部の種では鰓室で子貝まで哺育(ほいく)する。世界に約2万5000種、日本産はおよそ1500種あり大部分が海産であるが、わずかに淡水産があってもっぱら琵琶(びわ)湖に集中している。
[奥谷喬司]
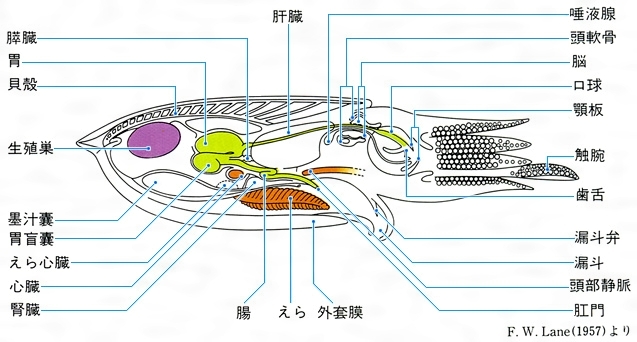
Cephalopoda 体外に殻をもつ現生種はオウムガイ科Nautilideaのみで、コウイカ科Sepiidaeの貝殻は舟形の「イカの甲」となり外套膜筋肉内に埋もれている。外套膜は袋状に内臓を包み、頭部はその前方に位置し、ここに高度に発達した目と「脳」とよばれるほど集中度の高い神経節が内在する。足は頭部の前にあり、オウムガイ類を除いては4または5対の腕に分かれ吸盤をもつ。口は環状に配置した腕の中央にあって歯舌と鋭い顎板(俗にからすとんびという)をもち、消化管はU字を描き肛門は前向きに開く。えらは1対の羽状本鰓(オウムガイ類のみは2対)をもち、根元にえら心臓がある。雌雄異体で、雄の腕のうちの1本が変形した交接腕で雌に精莢(せいきょう)を渡す。卵は卵黄が多く、部分割で胚盤(はいばん)が形成され、直達発生する。すべて海産で、現生およそ650種、日本近海には150~200種ぐらいが分布する。
[奥谷喬司]
軟体動物の貝殻は90%以上が炭酸カルシウムからなり、微量の炭酸マグネシウムやリン酸カルシウムも含まれている。炭酸カルシウムは方解石とあられ石の結晶となって存在し、これがコンキオリンconchiolinという有機物で接着されている。貝殻は通常、殻皮、殻質層、殻下層の3層からなっている。もっとも外側の殻皮はほとんどコンキオリンのみからなるため生時にすら失われやすいが、種によっては厚く毛状にさえなる。殻質層の典型的なものは、結晶が柱状に配列するので稜柱層(りょうちゅうそう)ともよばれる。殻下層は結晶配列が薄層状で、配列のぐあいによって光の屈折が異なり美しい光彩を現すので真珠層ともいわれる。貝殻は軟体動物の体を覆う外套膜から分泌される。外套膜の縁は三つのひだに分かれ、それぞれが前述の3層を分泌していく。殻下層だけはすでに形成された部分に密着している外套膜全体からも分泌補充され、貝の厚さを増す。貝殻の成長の方向は、二枚貝の場合は腹縁に沿って同心円を描くように継ぎ足されていく。巻き貝の場合は巻いた螺管の殻口の周囲に新しく継ぎ足され、螺管の長さと直径を増す。貝はほとんど絶えず成長するので微細にみると潮時に生ずる成長線もたどれるが、通常は温度の低下時や生殖時期に成長が鈍るので木の年輪のような休止帯が殻の上に残る。殻の成長には多量のカルシウムを必要とするので、巻き貝には軟体部の成長につれて内部の仕切りが吸収されてしまうものもある。淡水産の貝類ではほとんどが殻頂部を侵食される。
殻の上に形成される種々の彫刻は、一時的に成長の方向と量が変わるためにできる。たとえば巻き貝のアクキガイ科Muricidaeやフジツガイ科Cymatiidaeの一部では成長が突然に一時止まり、太い節のような縦張肋が形成される。縦張肋はかならず殻口のところにもあり、次の縦張肋が生ずるまでの間は外唇の補強ができないため、貝自身の活動はきわめて不活発である。成長の一時的な曲がりによってできた縦張肋は、のちに殻質で埋められる。アクキガイ科ではこの上に棘が生じ、本来は後溝にあたる肩部の棘がとくに強大となる。殻の表面には立体的な彫刻のほか、メラニン系やカロチン系の色素によりつくりだされるいろいろな模様や色がある。これらの色素は、殻質を分泌する外套膜縁の特定の位置にある細胞群から分泌される。殻質の分泌に伴う移動の結果、色素細胞どうしの間隔にも縮小・展開・断続がおこり、種により殻表面に不規則な文様や幾何学的な図形が描かれる。また、殻に沈着する色素は餌(えさ)によっても左右される。アワビ類HaliotisやサザエBatillus cornutusなどは、食べる藻類の種類により殻色に変化が現れる。
[奥谷喬司]
軟体動物が地球上に出現したのは、化石の産出状態から約6億年前の古生代カンブリア紀前期であると推定される。軟体動物の先祖は、環形動物や節足動物にみられるような体節構造をもち、海底をゆっくりはう動物であったと考えられる。柔らかい体を保護する石灰質の殻は、軟体動物の各綱が分化する段階で獲得したとされている。
腹足類はカンブリア紀に、二枚貝類は次のオルドビス紀に地球上に出現し、ともに海の底生動物のうちで重要な位置を占めるようになった。海洋に発生した腹足類が淡水域に進出するまでにそれから約3億年、二枚貝では2億年ぐらいかかった。腹足類には現在までに滅亡した科は意外に少なく、現存する科のなかに過去の進化系列のほとんどの段階がみられるといえる。したがってなかには「生きている化石」とよばれるオキナエビスガイ科Pleurotomariidaeなどもあるが、このように古い科において美しい色彩や彫刻があるのはむしろ例外的といえる。古生代においては腹足類に特有の体のねじれは少なく、中生代の白亜紀に入り科の数が一躍5倍となり繁栄の兆しがみえると、ねじれを経た体制の巻き貝すなわち捩神経類(れいしんけいるい)Streptoneuraが大勢を占めるに至った。その間、直神経類Euthyneuraである後鰓類の主流は、古生代の石炭紀になって現れた。他方、二枚貝類はオルドビス紀あたりから繁栄し始め、たとえば原鰓類と翼形類Pteriomorphiaの42科のうち、7科はオルドビス紀に、18科はシルル紀に、7科はデボン紀に、3科は石炭紀に、1科は中生代の三畳紀に出現した。掘足類や多板類が出現したのはカンブリア紀であるが、現在まで二枚貝や腹足類のように幅広い適応放散はみられず生息環境も限定され、種類も格段に少ない。現生では掘足綱はわずかに2科、多板綱は約10科にすぎない。
軟体動物のうちでも頭足類の歴史は複雑である。よく知られているアンモナイト(菊石)類ammoniteは、ほぼ4億年前のデボン紀に現れ、知られるだけでおよそ2万種も栄え、浅海域を中心に多様な形態と生活様式をもち、大形のものは直径2メートルに及ぶものすらあったが、中生代最後の白亜紀の終わりごろまでに、原因は不明であるが、突然全種が絶滅した。オウムガイ類は外形はアンモナイトに似たところもあるが直系の子孫ではなく、カンブリア紀後期から現れ、新生代の第三紀ごろまでにはほとんど絶滅し、熱帯西太平洋にわずかに4種だけが生き残っている。現在の海洋に栄えている二鰓類Dibranchiata(鞘形類(しょうけいるい)Coleoidea)は石炭紀に地球上に出現したが、ついに淡水圏への進出は果たせなかった。現生する軟体動物の属は、白亜紀末である6500万年前までに大部分が地球上に現れ、新生代第三紀に原始的な属で姿を消したものがあり、かわりに新しい属が加わった。淡水にすむ貝類は最近の100万年間に急激に種が増えたが、海産種は減少の傾向を示しているといわれる。
[奥谷喬司]
貝類は現在の地球上では昆虫類に次ぐ大群で、地球上のあらゆる地形に適応している。昆虫は表皮が拡張したはねを獲得することによって地上から空中に生活圏を伸ばしたが、貝類はもっぱら海洋中に繁栄し、潮間帯から水深1万メートルを超す超深海底帯にまですみ、陸上に進出した腹足類は、飛翔(ひしょう)こそできないが標高6000メートルの高山から洞穴や地下水にもすむ。海洋にすむ貝類は、大部分が潮間帯から陸棚斜面までに分布するといえ、深海にはごく限られた科がすむ。海洋の多岐にわたる条件と微地形の変化に応じて、形態、生活様式、繁殖戦略、呼吸方法、摂餌生態など多様な適応をみせている。
波の荒い岩礁にすむ腹足類や多板類の足裏は非常に広く、吸着力も強い。柔らかい砂や泥底にすむ二枚貝は、斧形の足で底質中に潜入し、殻の形や彫刻もすべて潜入生活に適応したものである。一見活発な巻き貝に比べ、二枚貝類は感覚器官や運動力に乏しいが、別の見方をすれば、軟体動物のうちで現在の地球上の海洋生活にもっとも高度に適応したとみることもできる。硬い岩礁にすむ二枚貝では、イシマテガイ属Lithophagaやカモメガイ属Pholusのように特殊な穿孔(せんこう)装置をもつもの以外は底質中に潜入生活することはできない。そのため、ワシノハガイ属Arcaやイガイ属Mytilusのように足糸という強靭な繊維質を分泌するか、イタボガキ科Ostreidaeやウミギクガイ科Spondylidaeのようにセメント質によって固い底質に固着する。貝類はこのように一般には移動力が弱いため分布域が限られている。その傾向はとくに陸産や淡水産貝類において顕著で、一つの山脈や谷、あるいは湖沼や河川系で種分化のおこっている例がある。海でも潮間帯種の分布は生物地理区ごとに種組成に強い地域性を示すことが多いが、ヤツシロガイ科Tonnidaeなどのように幼生の浮遊生活期が長く大洋の潮流にのって広域に分布伝播(でんぱ)している種もある。また、貝類の分布や生息域の拡大に人間の力があずかっているものも少なくない。日本にごく普通にみられるムラサキイガイMytilus edulisやシマメノウフネガイCrepidula onyxなどは北アメリカ原産で、おそらく船腹に付着して移入したものと思われる。逆に北アメリカには日本から移入したと思われるマルタニシCipangopaludina chinensis、オオウヨウラクガイOcenebra japonica、アサリRuditapes philippinarum、ホソウミニナBatillaria zonalisなどがみられる。このうちマルタニシはキンギョとともに、オオウヨウラクガイは種ガキについて移入したと考えられている。最近では黒海に日本産のアカニシRapana venosaが発生しているが、その移入経路はかならずしもはっきりしていない。
軟体動物のなかで、頭足類の大部分は本来の底生生活から大洋の中層や表層に進出し、魚類と比肩しうるほど遊泳生活に適応した。腹足類の一部にもこのような浮遊区にすむものがあり、たとえば翼足類Pteropodaや異足類Heteropodaなどは終生遊泳生活を送る。アサガオガイ科Janthinidaeは足裏から粘液を出し、これで浮嚢(ふのう)(いかだ)をつくり、その下につり下がって一生浮遊している。本来底生生活のものも外敵や悪環境から逃れるときは急激な跳躍や遊泳移動をする。ハマグリMeretrix lusoriaは、環境が不適になると長い粘液糸を分泌し、これを抵抗板として下げ潮(落潮流)に引かせて驚くほど大距離移動をするが、これはきわめて特異な移動法で、ほかの貝では知られていない。強い遊泳力のある頭足類では広く世界に分布する種も少なくない。
[奥谷喬司]
二枚貝類や下等な腹足類は体外に放卵・放精して水中で受精する。二枚貝類のなかでもフナクイムシ科Teredinidaeなどのように鰓室で子貝まで育つものがいくつかある。淡水性二枚貝も、鰓室内でベリジャー期に相当するグロキディウムglochidium幼生まで育ててから放出する。腹足類では卵や子貝の哺育はよくみられ、前鰓類のタニシ科Vivipariidaeやカワニナ科Pleuroceridaeも母貝の鰓室中で子貝となり、厳密な意味ではないがいわゆる胎生である。海産種には革質の卵嚢すなわちホオズキに入れて産出するものが多数ある。卵嚢からはベリジャー幼生期で孵出するものと、子貝になってはい出すものがあり、後者の場合一部の卵はほかの孵化幼生の栄養とされてしまう。後鰓類では寒天質の紐(ひも)状またはリボン状の卵塊を産む。頭足類も、卵を寒天質の卵嚢に入れて産み出す種が多い。浮遊性タコ類のアオイガイ科Argonautidaeは、雌の特殊に変化した第1腕から分泌された独特の貝殻の中で卵を哺育する。
[奥谷喬司]
軟体動物には発光するものがいくつかあり、二枚貝ではヒカリニオガイPholas dactylus、腹足類ではニュージーランドの洞穴にすむラチア属Latiaの一種などがその例である。頭足類ではイカ類の体表、腕、内臓や眼球の上などにとくに発達した発光器があり、種間の信号や威嚇のほか、海表面から降り注ぐ光量に見合う強さの光を発して背景に溶け込む護身用としても用いられる。閉眼類Myopsidaの発光は嚢中に発光バクテリアを収容する型であるが、開眼類Oegopsidaはレンズを備えた複雑な発光器をもち、おもに自身が発光する。軟体動物の発光はルシフェリン‐ルシフェラーゼ反応によるもので、昆虫類などと同様の生物発光(冷光)である。
[奥谷喬司]
軟体動物は歯舌という特有のそしゃく器官をもっている。歯舌は歯舌嚢からつくりだされ舌軟骨上にのっている器官で、軟体動物はこれを前後させておろし金(がね)で削り取るように餌を食べる。腹足類のうち、前鰓亜綱のミミガイ科Haliotidae、ニシキウズガイ科Trochidaeあるいはリュウテンサザエ科Turbinidaeなどのように海藻を専食するものの歯舌は1列の小歯数が多く、扇舌(せんぜつ)型や梁舌(りょうぜつ)型をしている。これに対して肉食性のものは全体としては餌の変化に富んでいるが、個々の科や種は餌の範囲が狭くそれに適応した歯型になっている。紐舌(じゅうぜつ)型歯型をもつタマガイ科Naticidaeや狭舌型のアクキガイ科は、酸や歯舌の作用により二枚貝などの貝殻に穿孔し、その穴から吻(ふん)を挿入して肉を食べる。同様に肉食性のものでも、フジツガイ科Cymatiidaeの多くはヒトデなどの棘皮(きょくひ)動物を襲い、エゾバイ科Buccinidaeやオリイレヨフバイ科Nassariidaeの諸種は死肉を好む。イモガイ科Conidaeの歯舌は矢状で矢舌型とよばれ、これを射て、種類により多毛類、星口類、貝類、魚類などを刺し、歯舌歯内に含まれる毒を注入し餌を殺して食べる。腹足類でも後鰓亜綱には裸鰓類Nudibranchiaのように腔腸動物などを食べるものがあり、なかには腔腸動物の刺胞や毒を体内に蓄え、それを防御用に用いる奇習をもつ種も含まれる。頭楯類(とうじゅんるい)Cephalaspideaには小形の貝類をまる飲みにするものがあり、これらの歯舌は貧弱であるが、胃中に石灰質の胃板がある。植物食性の種が多い有肺亜綱中にも肉食性種が少数いる。
二枚貝を除くすべての軟体動物は歯舌によって餌をとるが、二枚貝は水管から吸入した呼吸水に漂っているプランクトンやデトリタス(生物体の死骸(しがい)や分解途中の有機物)をえらで濾(こ)し取る。濾し取られた餌は鰓糸上の繊毛運動によって口へ運ばれ、口の前にある唇弁(しんべん)という器官で選択され、適当なものだけが食べられる。原鰓類はえらの濾過(ろか)能力が乏しいので、唇吻を殻外に出してデトリタスを吸い込む。また隔鰓類のえらは筋肉質で、その収縮力により水管から小形の甲殻類などを吸い込み、肉食性を示す。
掘足類は二枚貝と同様に砂泥底中に潜って暮らし、頭糸によって泥中の有機物を口に運ぶ。多板類は一般に腹足類と同じで、岩などに付着している微小藻類などを歯舌でかき取って食べているが、肉食性種もある。
[奥谷喬司]
腹足類や二枚貝類にはほかの動物の体外、体内に寄生生活しているものがある。ヤドリニナ科Stiliferidaeなどはその一例で、棘皮類の体外につくもののほか、体内に入り虫こぶとよく似たこぶをつくるものもある。寄生とはいえないが、ウミウサギガイ科Ovulidaeの諸種は、トゲトサカ類やヤギ類など種によってほぼ特定の寄主につき、そのポリプや共肉(各ポリプを連結する軟体部)を専食する。多くの場合、それら諸種の殻色、軟体の色や突起のようすは、餌とする腔腸動物の擬態になっていて、あたかも昆虫の幼虫が食草の保護色や擬態を示す場合と同様の関係である。ほかの貝類の表面につく貝類には、そこを単なるすみかとするもの以外にも、排出物を餌にするものや、カツラガイ科Capulidaeのように吻を寄主の二枚貝の口あたりに差し入れ、集めた餌を横取りして生活しているものもいる。成体期に自由生活する淡水性のイシガイ科Unionidaeでは、グロキディウム幼生は淡水魚のひれや体表に付着し体液を栄養としながら、自力では移動できないほど遠い距離を運ばれる。その代償として鰓室を淡水魚類の産卵床として利用させるが、なかには外套膜縁が産卵雌を誘う雄魚の体の擬態になっている種もある。サンゴ礁にすむシャコガイ科Tridacnidaeは外套膜の細胞中に鞭毛(べんもう)藻類の一種を寄生させ、その寄生種が生産する炭水化物を栄養源としている。寄生藻類を利用する貝は、シャコガイ以外にも2、3ある。
[奥谷喬司]
海辺における採集は、夏よりも春の大潮などの時期がよい。服装は水にぬれてもよい軽装で、かならず上着、帽子、軍手を着け、海藻の生えた岩の上でも滑らないように足袋(たび)や草鞋(わらじ)がよく、素足やゴム裏の履き物は適当でない。岩礁に行く場合は磯金(いそがね)やハンマー、ナイフなどを、砂浜や泥質の海岸ならば熊手(くまで)(アサリかき)や篩(ふるい)を必要とし、ほかにピンセット、ルーペ、ビニル袋や小瓶、胴乱などをもつ。
岩に固く吸い付いているカサガイ類などは殻をわずかに浮かせているとき、磯金や薄刃のナイフを差し入れてはがす。足糸でついている二枚貝も足糸をていねいに切ってとる。セメント質で固着している貝や穿孔生活しているものは、ハンマーで岩を割ってとる。転石のある海岸では、干潮時には多くの貝類がその下に潜んでいるので石をおこしてとるが、おこした石はかならず元どおりにしておくことが自然保護のうえで重要である。底質中に潜入しているものは熊手で掘り起こし、微小種は篩でふるって採集する。外洋の砂浜には波浪によって打ち上げられるものが多いが、玉石の多い海岸では貝が石の間に入ってしまい拾いにくい。台風のあとで海藻やごみが大量に打ち上げられたときは、貝類も多く漂着する。このほかテングサ干し場、エビ網やたこ壺(つぼ)の干し場、漁船の周囲、底引網漁船の水揚げ地などでは深い所の種類が拾える。またこのような所でみつかる底魚やヒトデなどに飲み込まれている貝や、ほかの動物に寄生している貝も見逃さないようにする。
淡水性貝類の採集は時季に関係がないが、水に入りやすく貝類も泥中に深く潜っていないので夏季がよい。琵琶湖のように貝類の大規模漁業のある所以外では、湖底の深部にすむものは舟や特殊な道具がないととれない。小さな種は岸辺の水草の間やスイレンの葉裏などにもいる。東南アジアなど地域によっては淡水に寄生虫の中間宿主となるものがいるので、水に入るときはこの点にも十分注意が肝要である。
陸貝は暖かい時期の雨上がりには活発になるのでみつけやすい。大きい種類は樹間、葉裏、朽ち木、石垣の間などに多く、神社やお寺の茂み、ごみ捨て場はよい採集場所である。小形種は落ち葉や石の下にいるので、それらごと集めて胴乱で持ち帰り、室内でゆっくり探すとよい。
[奥谷喬司]
巻き貝は煮てから中の肉を抜き、蓋(ふた)も保存する。二枚貝は熱湯につけ、殻が開いたら肉を抜く。貝類標本はこうして殻だけの乾燥標本とするのが普通であるが、解剖用や殻のないものは、70%以上のアルコールや5~10%の中性ホルマリンに全身をつけて保存する。標本にはかならず和名、学名のほか、採集地(産地)、採集年月日、採集者名を記した名札をつける。乾燥標本はガラス蓋付きの小箱に入れ、引き出しを利用してそれらを整理するのが最良である。乾燥標本も液浸標本も直射日光を避け、冷暗所に保存する。
[奥谷喬司]
カキの粗放的な養殖はすでに古く古代ローマ時代から行われていた。カキ養殖に革命をもたらしたのは昭和初期に始まった垂下式養殖法で、瓦や貝殻などのコレクター(付着器)に稚貝(スパット)を付着させ、これを連ねて水中につり下げるこの方法により、きわめて高密度な養殖が可能になった。さらにこのような連(れん)を筏(いかだ)につるすことによって、干出時間すなわち貝が餌をとれない時間をまったくなくし身入りをよくさせた。最近発達したホタテガイ養殖などでは人工採苗した稚貝を網の袋に入れて海中につるすが、原理的には同じである。日本における規模の大きい養殖は真珠を目的とするアコヤガイでみられ、人工採苗した稚貝を養成してから核入れの手術を施し、核の周囲に真珠層が巻くのを待って取り上げる。養成の方法はアコヤガイを入れた籠(かご)を筏からつり下げ、立体的に高密度飼育をするもので、原理はカキなどと同様である。二枚貝に比べ、巻き貝の養殖は昭和30年代ごろに始まったものである。アワビ類の養殖は、現在では人工採苗から稚貝育成までの技術が進んだが、天敵被害に対して抵抗性のできるサイズまで人手で育ててからは天然の漁場に放流するので、二枚貝類のように生涯または成長後の養殖育成とは趣(おもむき)を異にしている。
[奥谷喬司]
貝類は食用のみならず、装飾品、生活用具、貨幣、宗教の対象、楽器、染料の原料など、人間生活に益するところが非常に大きい生物群の一つであり、長い利用の歴史をもつが、なかには害をなすものもある。たとえば、人間や家畜の寄生虫の中間宿主となるカタヤマガイやモノアラガイ、刺毒をもつアンボイナガイ、咬(こう)毒をもつヒョウモンダコ、海中の構造物を穿孔破壊するフナクイムシやカモメガイ、有用貝類や養殖貝類に被害を与えるマダコやオオウヨウラクガイ、野菜や有用植物を食害するウスカワマイマイやアフリカマイマイ、発電所などの冷却水パイプを詰まらせてしまうムラサキイガイなどがその実例であるが、これらはすべて貝類本来の生態が人間活動に災いしているものである。
[奥谷喬司]
貝類のなかには魚におけるフグ類のように本来有毒のものはないが、エゾボラ類のように唾液(だえき)腺に麻痺(まひ)毒をもつものがある。また、二枚貝にも巻き貝にも餌になった生物に由来する外因性の毒が蓄積していることがあるので、食用にするときには産出の地方、季節などに十分注意する必要がある。
種によって産卵期が異なるので一概にはいえないが、貝はおおむね春先が旬(しゅん)である。貝の主成分はタンパク質であるが、リン、グリコーゲンのほかビタミン類も多い。生食する場合、高温期は生殖巣をはじめ内臓がいたみやすいので注意が要る。英語やフランス語などで、つづりにRのつかない月すなわち5~8月にはカキを食べないほうがよいという言い伝えも、このことを表している。適期に生食する場合にも、清潔な環境で採捕・育成されたもので新鮮なものを選ぶのは当然である。カキなどは軟体全部を生食するが、多くの貝では足部、閉殻筋(貝柱)、外套膜縁(ひも)のような筋肉部のみを食べ、内臓は一般に除く。頭足類の場合は外套膜が主要な可食部であるが、ホタルイカなどは全体を生食する。湯に通すだけで美味なものはミルクイガイ(ミルガイ)やナミガイ(シロミル)の水管などである。火を通す場合にも、長時間の煮炊きをすると肉の繊維が硬くなりうま味は失われる。粘液質の多いバイ類などは塩でもんでぬめりを落とす。煮てから二杯酢や三杯酢にするとあうのはエゾバイ科(ツブ類)やアカニシ、タコで、ツブ類やタニシはぬたにも向く。
二枚貝類でもハマグリは生食することは少なく、殻付きの焼きはまぐりにもするが、アサリ、シジミと同じくだしがよく出るので、潮(うしお)汁、みそ汁あるいは鍋(なべ)物の実とすることが多い。アケガイやアラスジケマンガイほか同科の二枚貝類は、各地方で各種が用いられている。洋風のチャウダーにも主として二枚貝を用いるが、カリブ海地域ではチャウダーガイという巻き貝も用いるようである。殻ごと焼くのはハマグリに限らず、ホタテガイやアゲマキガイなどの二枚貝でも行われる。一方、巻き貝の場合はサザエの壺焼きが代表的であるが、この方法はサザエのみでなくツブ類でも風味が出ておいしい。壺焼きは、貝の胃中にある餌にした石灰藻などが口に当たることがあるので、肉部だけ先に取り出しミツバなどと刻み込んで煮沸するようにも調理するが、やはりそのままで内臓まで火を通す方法が野趣があってよい。乾製品の代表的なものはイカ類のするめであるが、ホタテガイの貝柱やウミタケガイの水管、しばしばヒメガイの名で市場に出るシオフキガイそのほかも用いられる。とくにアワビの乾製品である明鮑(めいほう)や灰鮑は、中華料理になくてはならないものである。また、薫製、煮つけ、みそ漬け、粕(かす)漬け、塩辛などの加工品も多様である。
[奥谷喬司]
人類にとって貝類は、野生動物のうちでも採取がきわめて容易であるため、海産、淡水産、陸産のいずれも重要な食料として、採集狩猟生活のころから利用されてきた。それがとくに大量に消費されるようになったのは、旧石器時代後期ごろと考えられ、これは人類が海浜や入り江の近くに季節的な移住、ないしは定住生活に移行し始める時期とほぼ一致する。限られた種類の貝類を重点的に、かつ長期にわたって食用にあてていたことは、貝塚に残されたおびただしい貝殻からも知られ、海産の貝殻が内陸からも出土していることは、交易が行われていたことを示している。また今日、貝の養殖はホタテガイ、アサリ、ハマグリ、真珠貝、アワビなど多種に及んでいるが、北方海域でとりわけ多く賞味されているカキ類の養殖は、早くもローマ時代に始まっていたといわれる。
貝類は食用として貴重なタンパク源となったばかりでなく、石灰質の殻が堅固で変質しにくく、美しい色彩や光沢を有することから、腕輪や首飾り、佩具(はいぐ)などの装身具としても世界各地の民族の間で用いられてきた。日本でも、貝輪が縄文時代からもっとも一般的な装身具として愛用されてきたが、装身具のなかには呪具(じゅぐ)の性格をもつものや、貨幣としての機能を帯びるものもあり、中国では周(しゅう)代にタカラガイを貨幣に用いたことが知られている。そのほか、貝殻の神秘的な形状や色彩、光沢をよく生かしたものに螺鈿(らでん)や象眼(ぞうがん)などの貝細工も知られ、真珠は古くからペルシア湾、スリランカ、中国、東南アジアの産地をはじめ、宝石として各地で尊ばれている。より実用的な用途としては、日本の場合だけをみても、貝器類、顔料(胡粉(ごふん))、玩具(がんぐ)やゲーム(貝合(かいあわせ)、貝独楽(ごま)、碁石(ごいし))、ボタン、古いものでは法螺貝(ほらがい)などがあり、また潮の香りと海辺の旅情を伝えるものとして、土産(みやげ)品や置物にも幅広く利用されている。
[伊藤亜人]
『吉良哲明著『原色日本貝類図鑑』(1959・保育社)』▽『波部忠重著『続原色日本貝類図鑑』(1961・保育社)』▽『波部忠重監修、奥谷喬司著『学研生物図鑑 貝Ⅰ・Ⅱ』(1983・学習研究社)』▽『奥谷喬司著『自然観察シリーズ18 日本の貝』(1983・小学館)』
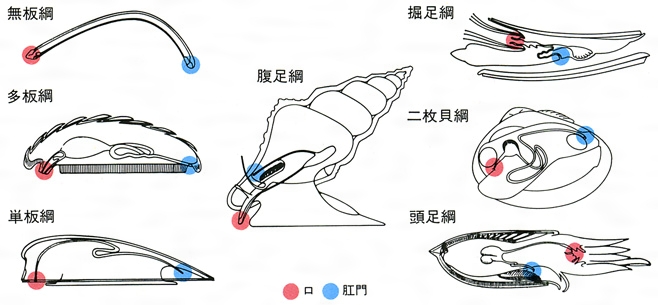
軟体動物7綱の比較
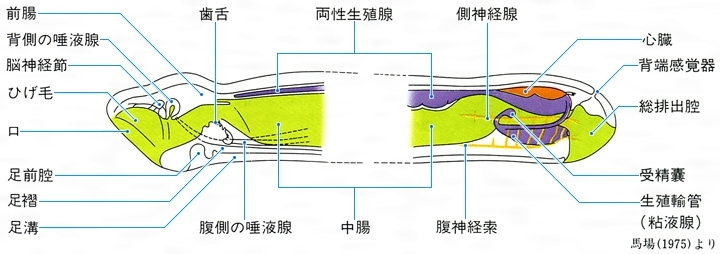
無板綱の体制模式図

カセミミズ(生態)
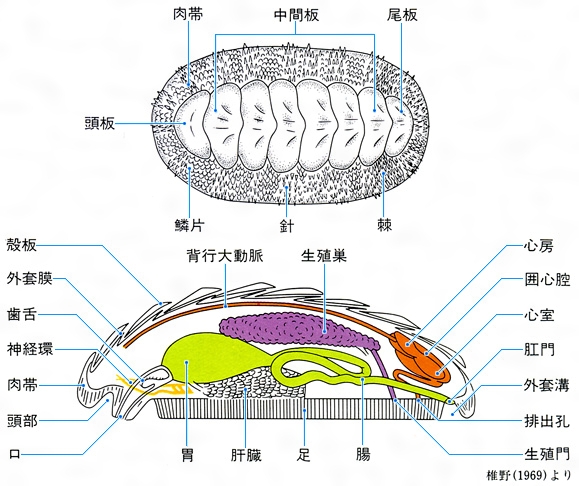
多板綱の体制模式図

ヒザラガイ(生態)
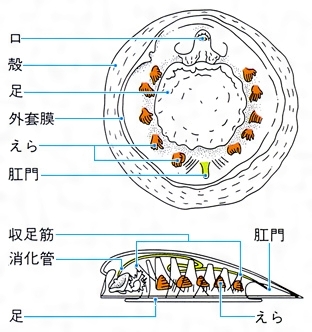
単板綱の体制模式図
腹足綱の体制模式図

バイ(生態)

エムラミノウミウシ(生態)

掘足綱の体制模式図

ヒメナガツノガイ(生態)
二枚貝綱の体制模式図

トリガイ(生態)
頭足綱の体制模式図

コウイカ(生態)

スナダコ(生態)

イタヤガイ(生態)

ウミウサギガイ類の1種(生態)

カタヤマガイ(生態)

カモメガイ(生態)

クリイロヤドリニナ(生態)

タイラギ(生態)

ハマグリ(生態)

フナクイムシ(生態)

ホラガイ(生態)

マルタニシ(生態)

モモガイ(生態)

貝輪

法螺
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponsored by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponsored by ![]()
Sponsored by ![]()
政府首脳が外国を訪問した際の会談内容や合意事項を記した外交文書。法的拘束力は持たないが,その内容は両国を事実上拘束する。類似のものに共同発表 joint statementがあるが,これはより記録的な...
7/28 化学辞典 第2版(森北出版)を更新
6/26 日本大百科全書(ニッポニカ)を更新
4/17 デジタル大辞泉プラスを更新
4/17 デジタル大辞泉を更新
2/17 日本大百科全書(ニッポニカ)を更新