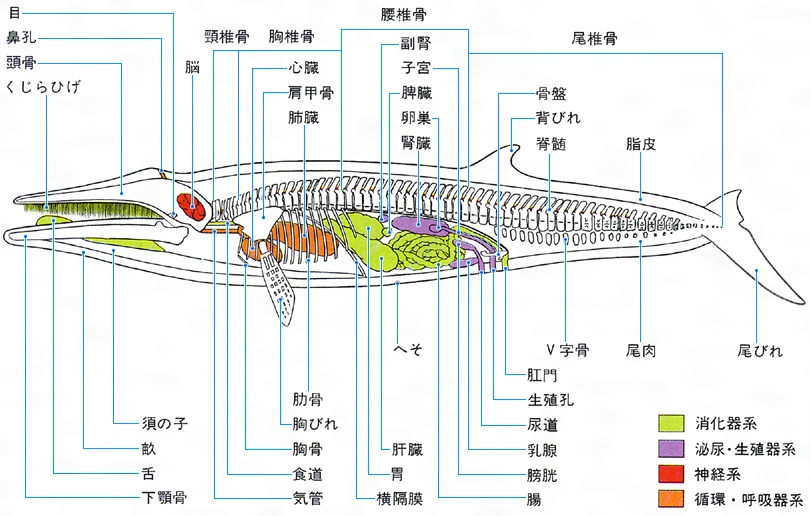
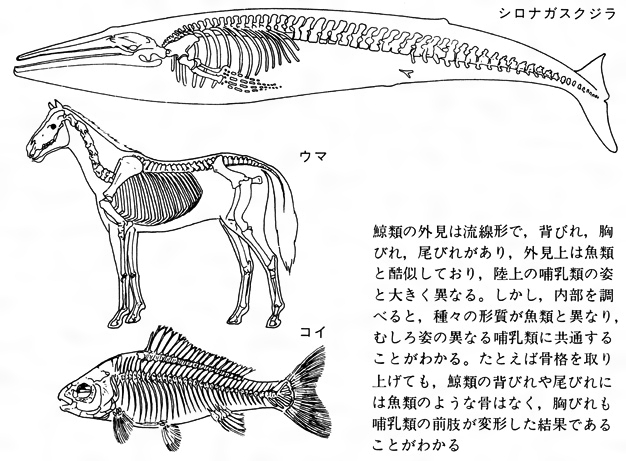
クジラ目Cetaceaに属する哺乳類の総称。鯨類(げいるい)ともいう。体長数m以上の大型種を習慣的にクジラ,それ以下をイルカと呼ぶこともあるが,この区別ははっきりしたものではない。終生水中で生活する。体は紡錘形,後肢はなく,前肢はひれとなり,尾端の皮膚は水平にのび尾びれとなる。皮膚は皮脂腺,汗腺,被毛を欠き滑らかで,真皮は厚い脂肪層となる。ヒゲクジラ類では頭部に感覚毛を残す。
分類
クジラ目はムカシクジラ類,ヒゲクジラ類,ハクジラ類の3亜目よりなる。
ムカシクジラ類Archaeocetiは,第三紀始新世初期に,地中海から東アジアに広がっていたテチス海で発生し,漸新世初めに絶滅した。最古の化石は,パキスタン産の頭骨の一部で2mくらいの動物Pakicetus属で,四肢を使って陸上歩行もした。この系統からは,始新世末期になると,イルカに似たDorudon属や全長十数mのBasilosaurus属のような進んだグループが出現した。まだ小さな後肢が残っていたが,胸びれと尾びれの形は現生クジラ類と差がなかったらしい。歯式は切歯3,犬歯1,小臼歯(きゆうし)4,大臼歯3で,合計44本と真獣類の一般型をとどめ,大臼歯と後方の小臼歯は3根で,複数の咬頭(こうとう)を残していた。鼻孔は吻(ふん)の中ほどにあった。ムカシクジラ類から現生の2亜目が派生した。
ハクジラ(歯鯨)類Odontocetiでは,顎骨が後方にのび頭骨背面を覆い鼻孔は目の位置まで後退している。左右の鼻道は皮下で合一し,1孔が開口する。最古のハクジラ類は漸新世初期のSqualodon属やAgorophius属で,歯は上下とも13~26対に増えて,後方の歯はサメの歯に似た形をし2~3根を残していた。中新世には,現生ハクジラ類の5系列(科)が出現し,同型歯となった。マッコウクジラ科は2属3種よりなり,上顎歯が退化傾向にある。頭骨の顔面部には巨大な脂肪組織(脳油)を入れるくぼみを備えている。この脂肪組織は,音響レンズであるとも浮力調節をするともいわれる。なお,コマッコウ属2種を別科Kogiidaeとして独立させることがある。アカボウクジラ科は,体長4~11mの中型種5属19種よりなる。吻が長いが,ふつう下あごに1~2対の大きな歯を残すのみである。カワイルカ科は4属5種の原始的なイルカで,浅海あるいは淡水にすむ。このグループの分類のしかたは研究者により意見が分かれ,1科4属とするものから,1上科4科4属とする意見まである。マイルカ科は小~中型種17属約33種よりなるハクジラ類中最大のグループである。ネズミイルカ科は小型種4属6種を含む。吻は短く歯は小さい。英語ではこれをporpoiseと呼ぶ。本科とイッカク科は,マイルカ科に近いグループから派生したらしい。
ヒゲクジラ(鬚鯨)類Mysticetiでは,顎骨の主要部は後縁が眼窩(がんか)前方にとどまり,頭骨背面を覆わない。鼻孔は吻の基部背面に位置し,体外に2孔が開口する。胎児期に数十対の歯胚が形成されるがまもなく消失し,口蓋にひげ板を生ずる。ひげ板の形,色,質などは重要な分類形質である。現生のヒゲクジラ類は4科6属11種で,大型種が多い。化石資料が乏しく,各科の系統的な関係は明らかではない。ナガスクジラ科は中新世中期に出現し,現生2属5種。中新世の前・中期に栄えたセトテリウム科Cetotheriidaeに近い科と考える見方がある。本科のナガスクジラ属のニタリクジラとミンククジラは分類学的に未解明で,将来種数が増える可能性がある。セミクジラ科は中新世末期に出現し,現生2属3種。アーチ状に湾曲した細い吻をもつ。コククジラ科は1属1種。
分布
クジラ類の分布は,水温,餌料,地形などに影響される。また,人間活動によって分布が狭められたり,生息数が減少している例が多い。ヒゲクジラ類は一般に夏に高緯度海域で索餌し,冬に低緯度海域で繁殖を行う。この年周期に合わせて,妊娠期間は1年前後になっている。北極圏にいるホッキョククジラと低緯度海域のニタリクジラは例外で,水塊間の回遊は行わない。ハクジラ類のうちアカボウクジラとシャチは例外的に熱帯から寒帯にまで分布するが,イルカを含めたハクジラ類の多くは大回遊をせず,一定の水温範囲に生活する。日本近海では黒潮系,親潮系のイルカと中間的な種(カマイルカ,セミイルカ)と3グループがある。マッコウクジラは雄は高緯度海域に回遊するが,雌とコドモからなる繁殖群は暖水域にとどまるので基本的には暖海種である。クジラ類には沿岸で生活する種と沖合を好む種がある。大型のクジラではセミクジラ,ザトウクジラ,コククジラが前者に属する。ハクジラ類には淡水種から沖合種まである。日本沿岸ではスナメリとネズミイルカはもっぱら大陸棚上に生息し,カマイルカとハンドウイルカも比較的沿岸に近づくが,外洋にも広く分布する。水温からみた生活環境は赤道を中心に対称的に配置されているので,同種あるいは類似した種が南北に分布することになる。このような例はナガスクジラ科,セミクジラ,アカボウクジラ科,カマイルカ属,ネズミイルカ属などにみられる。これは,かつて海水温が低下し,大洋の東側ではクジラの南北交流の可能な時期があった証拠とみなされ,その最後の機会が約1万年前に終わったウルム氷期と考えられる。南北の対応種の間の体長や体色などの亜種レベルの差はその後生じたものであろう。北極圏周辺に固有種が多いのは,赤道海域の水温低下が不十分で,北極圏の種は赤道を越える機会がなかったためであろう。今日の北太平洋のコククジラはウルム氷期の後の温暖期に北極圏を通って北大西洋とのあいだで交流があったと思われる。北大西洋では,コククジラは捕鯨によって18世紀ころ絶滅した。北大西洋にいたコククジラが氷河期に南半球に広がらなかったのは,夏を過ごす高緯度浅海域がアフリカ大陸南部にはなかったためであろう。カワイルカ類では近縁種がアジアと南アメリカの大河流域と沿岸域に離れて分布している。これはかつて分布の広がったグループの遺存種であることによる。
食性
クジラ類の食性は融通性に富んでおり,同種でも,海域や季節ごとに入手しやすいものを食べる。ハクジラ類にこの傾向が強い。外洋性の小型イルカ類は主としてハダカイワシ,アジ,サバ,イカ,エビなどを捕食する。沿岸性のネズミイルカの餌はタラ,ニシンなどが多い。体が大きく強大な歯をもっているオキゴンドウではブリ,マグロ,イカがおもな餌となる。シャチも同じく大型魚類,アザラシ,クジラ類などを捕食する。歯の比較的弱いアカボウクジラ,ハナゴンドウ,マゴンドウなどはイカが主体となる。マッコウクジラではイカ類,底生性大型魚がおもな餌である。ハクジラ類の餌の共通要素はイカ類である。イカへの依存度が高い種ほど体の巨大化と歯数の減少が進んでいる。巨大化は豊富で入手しやすいイカを常食とすることによって可能となったばかりでなく,体温の保持と深潜水に有利である。ハクジラ類には夜間に餌料生物が海面近くに浮上したときに索餌したり,暗黒の深海にすむ種を捕食する種が多く,彼らは音響探測で餌を探知する。また,ハダカイワシなどの生物発光や魚類の鳴声(ラプラタカワイルカ)を手がかりにしたり,深海性のイカがクジラの白い口唇や歯に誘われて寄ってくるのを捕食することもあるといわれる。
ヒゲクジラ類に特有の索餌器官はクジラひげと〈畝(うね)〉である。前者は口蓋から二列に垂れる三角形の角質板で,管状の髄質と外側の外被層よりなる。ひげ板の舌側の縁は索餌に伴って徐々にくずれて,髄質が剛毛となって露出し,これがからみ合って口内に大きなフィルターを形成する。ナガスクジラ科に特有な〈畝〉は下あごの先から腹部にかけて縦に並ぶ皮膚のひだで,皮下筋肉が発達して伸展性に富む。ヒゲクジラの索餌方法には3型がある。第1は,セミクジラ類の方法で,口を開けたまま餌の集団の中を泳ぎ回り,口中にたまった餌を舌で集めて飲みこむ方法である。この種のクジラでは舌は筋肉質で,クジラひげが長く,頭は体長の1/3~1/4を占めている。おもにプランクトンを捕食するが,ホッキョククジラではときにはコククジラのように海底で索餌するのが観察されている。第2はコククジラの方法で,横泳ぎをしつつ口唇とひげ板で海底の泥をすくい上げて,同じく筋肉質の舌を動かして,これを吸い込む。そのあと舌を使って泥を吐き出し,ゴカイや底生の甲殻類をこしとる。ときには,海岸の海藻をしごいて甲殻類を集めたりする。ひげ板は硬くて短い。第3はナガスクジラ科の索餌法で,餌に泳ぎ寄って口を開けると同時に畝をたるませて,口からのどのあたりに巨大なスペースをつくり,ここに一気に海水を取り込んで口を閉じる。続いて畝を収縮させて,ひげ板の間から海水を排出し餌だけを残す。この方法でプランクトン以外にサンマやニシンなどの群集性魚類もとらえる。
クジラ類の胃は,ふつう4室よりなる。第一胃は食道の膨大したもので,食物を一時蓄える。第二胃以下が消化腺を備えた真の胃である。ナガスクジラ類の胃には,0.5~1tの餌が入っていることもある。餌育実験と熱消費の推算によれば,1日当り摂餌量は,小型のネズミイルカで体重の9%前後,ヒゲクジラでは夏の索餌場で4%,冬の繁殖海域で0.4%,平均1.5~2%である。ヒゲクジラは夏季に皮下,骨,内臓などに脂肪を蓄え,冬にこれを消費する。ハクジラ類では貯蔵脂肪の周年変化は著しくない。陸上哺乳類の基礎代謝量(M:kcal/日)と体重(W:kg)の関係,M=70W0・75はクジラ類にも適用でき,体重50kgのネズミイルカで1300kcal/日,100tのシロナガスで40万kcal/日となる。これはクジラ類の皮下脂肪層が熱の散逸を防ぎ,36℃前後の体温を保持しているためである。活動に必要な熱量は上の値の約8倍で,成長と繁殖にも別に熱量を必要とする。クジラのミルクは糖が少なく脂肪が多く(水分41~54%,脂肪30~37%,残り7~22%は主としてタンパク質),熱量は3.7~4.3kcal/gである。クジラ類では餌の供給量が増加すると性成熟年齢が低下し,出産間隔が短くなる傾向がある。
遊泳
クジラ類の遊泳速度はイルカもヒゲクジラも大差なく,最高時速30~40km,巡航速度10km前後である。通常の潜水はヒゲクジラで200m,30分以内とされているが,研究が進めば,もっと深い潜水が確認されると予想される。ハクジラではスナメリのような数十m,数分から,マッコウクジラやツチクジラのように1000m以上,1時間をこえる種類まである。水中ではクジラ類は脈拍を減らし,脳や心臓に向かう血液を除き,大部分の血液は毛細血管を経ずに怪網と称する短絡路を通って静脈に戻る。潜水中は骨格筋はおもに嫌気的呼吸を行うので,大潜水後は頻繁に呼吸して酸素を補給する。潜水中は血液中に貯えられた酸素のほかに,筋肉中の酸素も使用する。このためクジラ類,とくにハクジラ類では筋肉中の呼吸色素ミオグロビンが多く,肉が黒っぽい。潜水中は水圧によって肺胞は完全につぶれて,空気は血液との接触を断たれるので,潜水病にかからない。
感覚
知覚も水中適応の結果変化した。ハクジラ類は嗅神経を失ったが,ヒゲクジラ類では残っており,嗅覚(きゆうかく)があるかもしれない。味蕾(みらい)はヒゲクジラ類にはないといわれる。ハクジラ類では,成長にともなって味蕾が退化するといわれるが,成体でも化学知覚が鋭いことが知られており,味蕾に代わって水中の化学物質を感知する器官が舌に発達している。水中の視程は最大数十mであるから視覚はあまり有効でない。カワイルカ類のように視覚が退化ないしは消失した種もある。イルカ類に見られるコントラストの強い体色斑は,悪条件下での同種の識別に効果的である。触覚は頭部,口唇,胸びれなどで鋭敏で,仲間とのコミュニケーションや索餌に役だつ。ヒゲクジラの頭部の感覚毛は水流,餌料生物などの感知に役だつ。鳴声と聴覚の役割は大きい。クジラ類は声帯を欠くが声を出す。ハクジラ類は鼻道の途中にある3対の鼻囊とその間の弁で音を出すのか,喉頭部で出すのか定説がなく,両者が別の音を出す可能性もある。鼻囊とその前方の脂肪組織(メロン)は音の反射板と音を集束させるレンズの働きをする。ヒゲクジラでは鼻囊を欠くので,喉頭部で音を出していると推定される。内耳の水中音響への適応はムカシクジラ類に始まったが,蝸牛(かぎゆう)部の構造の高周波音への適応はハクジラ類に始まっている。さらにハクジラ類では内耳を含む骨は頭骨から遊離しており,かつ空気囊に囲まれ周囲の雑音から遮断されている。音は下あごから伝わり耳に達するので,首を振って音源を探知する。繁殖期のザトウクジラのさえずり音は有名であるが,似た鳴音はホッキョククジラとセミクジラでも知られる。多くのヒゲクジラはうなり声に似た低い音(20~2000Hz)を出すことが知られている。低い音は減衰しにくく,遠く離れた個体間で交信したり,氷の状態を探知するのではないかといわれている。ヒゲクジラが音響探測を行うという確証はないが,索餌中のシロナガスクジラとコイワシクジラの付近で録音された高周波音(4~31kHz)はその可能性を示している。ハクジラ類の鳴声には,パルス状のクリックと口笛状のホイッスルとがある。これらの音を同時に出すこともでき,種類によって使用の比率が異なる。前者は数百Hzから200kHzの広い周波数域の音を含み,人間の耳にはカッカッとかギーとか聞こえる。主として音響探測に,ときには感情の伝達に用いられる。直径数mmの近距離の物体を,また300m離れた小魚の群れを探知できる。すべてのハクジラ類がこの音を出す。ホイッスルは一連の純音で,高さが連続的に変化する。マッコウクジラ,アマゾンカワイルカ,スナメリなどはこの音を出さない。複雑な音であるため,イルカの言語であると考える人もある。しかし,クジラ類が感情,発情および仲間であることを示すために声を用いていることはわかっているが,〈思想〉や〈論理〉を伝えることばをもつことを示す証拠はない。
生殖
クジラ類は単独あるいは群れで生活している。群生する場合もリーダーに相当する個体は認められていない。複数の群れが一時的に集まってできた大集団は,群れとは区別する。ヒゲクジラ類,カワイルカ類,ネズミイルカ類の群れは数頭以下で,持続的な個体間の結びつきは親子関係だけらしい。社会構造のもっとも未発達なグループである。マイルカ科では雌雄の成熟個体と子イルカを含む数十~数百頭の永続的な群れを形成するが,雌雄とも群れからの出入りが多い。これらの種では出産間隔は2~4年であるので,群れは妊娠,泌乳などの状態にある成熟雌をつねにもっている。スジイルカやマダライルカでは,生後数ヵ月で餌をとり始め,1~3歳で離乳が完成すると親の群れから離れて子どもだけの群れをつくる傾向がある。この傾向は雄に著しい。成熟すると他の成熟個体の群れに加わるが,自分の母親の群れに戻るという保証はない。この習性がさらに発達したのがマッコウクジラで,群れの基本は20~40頭の母系の血縁集団であり,繁殖期以外は成熟雄が入っていない。雄は春機発動期(10年)以後母クジラの群れを離れ,繁殖集団との競合をさけてしだいに高緯度地方に離れすむ。雌は成熟後も群れに残る傾向が強い。27歳以上の十分に成熟した雄は,繁殖期になると暖かい海にやってきて,交尾相手を求めて雌の群れを訪れて歩く。このときに雄同士が出会えば闘いが起こることもある。日本近海のコビレゴンドウの群れ(15~30頭)も母系の血縁集団であるが,常に数頭の成熟雄が含まれている。雄が雌から離れて生活することはないが,群れから群れへ移る可能性が残っている。クジラ類の群れの中で,構成員がもっとも固定しているのが,カナダ太平洋岸のバンクーバー島周辺のシャチの群れである。本種の群れは10~50頭の家族よりなり,各群れは固有の鳴声をもっている。少数の成熟雄を含む場合と含まない場合がある。発情時に一時他の群れと合流することがあっても,いずれ元の組合せに分離する。このような群れでは,群数の増加はまれに起こる群れの分裂によるだけである。社会構造の発達したハクジラでは出産率が低く,上記3種では平均5~10年に1産であり,雌は最後の出産をすませたあと,20~30年に及ぶ老年期をもつことで知られている。逆にヒゲクジラやネズミイルカ類では高死亡,高出産の傾向があり,1~3年に1産で更年期は知られていない。
脳
脳の重さはヒゲクジラでは3~7kg,体重の0.03%以下である。ハクジラ類,なかでもマイルカ科は1~6kg,体重の1~1.6%の脳をもちヒトの2%に近く,大脳の回転(しわ)も多い。これとイルカ類の複雑な鳴声とを結びつけて,イルカが高い知能と言語をもつのではないかといわれる。しかし,飼育下のハンドウイルカの問題解決能力と大脳皮質中の神経細胞の密度はゾウ程度にすぎず,大脳の大部分は聴覚中枢で占められているともいわれ,イルカ類が人間に匹敵する知能をもつ証拠は得られていない。
→イルカ →捕鯨
執筆者:粕谷 俊雄
利用
クジラは食用,工業用,工芸用と広く利用されるが,主要目的は,鯨種,利用国の国情,さらには歴史的発展段階によっても異なる。古代捕鯨(11~18世紀),アメリカ式捕鯨(17~19世紀)年代における欧米諸国では灯火用の鯨油およびセミクジラ,ホッキョククジラのクジラひげが主要目的とされ,ノルウェーにおけるミンククジラを除き鯨肉類はほとんど利用されていない。クジラひげはおもに当時の婦人服には欠くことのできないコルセットに利用され,婦人の美意識がセミクジラ,ホッキョククジラ資源の著しい減少に関係したという意外な側面がある。マッコウクジラはアメリカ式捕鯨によって多く捕獲されたが,マッコウ鯨油より生産されるろうそくは当時のアメリカの最重要輸出品となり,同国の産業発展に大きく貢献した。近代捕鯨に至ってからも欧米諸国の主利用目的は鯨油であり,国際捕鯨委員会によって適用されていたBWU制度(1972廃止)はシロナガスクジラの鯨油生産量を基準に定められていた。しかしながら1859年にペンシルベニアで石油が発見されて以来,この産業の発展により鯨油の価値は著しく低下し,欧米諸国の捕鯨撤退の大きな原因となった。
一方,日本においては,クジラはおもに食用として広く利用され今日に至っている。食用とされる部位は肉にとどまらず,脂皮,内臓,軟骨部などまでが利用される。第2次世界大戦後,GHQは国内の食糧事情悪化打開のためにマッカーサーラインを越えて捕鯨を許可し,捕鯨業は戦後の国内復興に大きく貢献した。国内市場における動物性タンパク質生産のうち,鯨肉の占める割合は1947年には47%を占め62年には生産量20万tに達したが,近年の捕獲枠減少により現在では3%を占めるにすぎない。しかしながらその需要は依然として高い水準にある。
(1)鯨肉類 ヒゲクジラ類の鯨肉は刺身,ステーキ用の尾肉赤肉(背肉,腹肉)がよく知られているが,尾肉は牛肉のロース状を呈し味および希少価値から最上とされる。胸肉,小切肉などはやや品質が劣りおもにハム・ソーセージ,缶詰などの原料となる。ナガスクジラ科鯨類特有の畝の部分は下層にあたる〈須の子〉とともにベーコン原料として利用され,〈須の子〉と〈須払い〉は缶詰原料となる。そのほか前肢基部の〈脂須の子〉,下顎基部の〈鹿の子〉は希少価値から高値で取引される。一方,マッコウクジラなどのハクジラ類の肉はその食性の関係上品質色彩ともに劣り,加工後にスープ原料などに用いられるが,食用としてはあまり利用されていない。しかし近年では鯨肉全体の供給量の減少から食用肉としての価値が見直されている。
(2)脂皮類 本来採油原料として用いられるが,ヒゲクジラでは近年鯨油価が低下し,もっぱら原形で食用とされる場合が多い。マッコウクジラの脂皮の一部は〈コロ(皮鯨)〉と呼ばれるものに加工され,関西方面のおでん材料には不可欠なものとなっている。
(3)内臓類 〈百尋〉と呼ばれる小腸をはじめとし,ほとんどの内臓類が食用となるが,これらはおもに九州地方で消費される場合がほとんどである。肝臓や膵臓などは薬品原料としても用いられる。
(4)鯨油 鯨油は鯨体の脂皮,骨類および内臓類より採油される。ヒゲクジラ類の鯨油はナガス油と総称され,脂肪酸とグリセリンからなるグリセリドを主成分とする。食用としてはマーガリン,ショートニングに用いられる。工業用としては,添加原料の反応中間原料として利用される場合もあるが概して利用途は狭く,現在では工業用としての価値は低い。マッコウクジラから生産されるマッコウ油は脂肪酸と高級アルコールからなる蠟を主成分とし,その特性として金属表面を容易に湿らせること,摩擦係数が低いこと,比重が大きく粘性が低いこと,腐食防止性があることなどから各種の工業用油剤として使用され,食用とされることはない。なお,マッコウ油は原料の部位によって体油,脳油とに区別され利用される場合もある。江戸時代には田の害虫駆除にも用いられた。
(5)その他 クジラひげ(筬(おさ))は,セミクジラなどでは良質で古くは〈からくり人形〉などの動力(ぜんまい)として利用されたり釣ざおの先に用いられ,前述のようにヨーロッパではコルセットなどに利用されていた。しかし近代になって同種の捕獲禁止によって,利用途が狭まり現在では一部が茶托,靴べらなどに加工されるが,プラスチック工業の発展によって市場が狭まり,むしろ希少価値を強調した民芸品的な色彩が強い。なお,正倉院には上代に用いられたと思われるクジラひげ製の如意(孫の手のようなもの)および幡心(仏殿内装飾品)が所蔵されており,鯨類の利用がかなり昔から幅広く行われていたことが示唆される。マッコウクジラの歯は装飾品,パイプ,印材などに用いられるが,近年希少価値からきわめて高価に取引される。また同種腸内よりまれに発見される竜涎香は香料の香気安定保留剤として高い価値をもっている。そのほか頭蓋骨中にある細長い軟骨は〈かぶら〉と呼ばれ,一部はコンドロイチン原料としても利用される。
執筆者:加藤 秀弘
鯨肉の利用史
縄文,弥生の遺跡から骨が出土していることから見ても,日本の鯨肉食用の歴史は古い。そして,天武朝以後しだいに肉食忌避の風潮が拡大する中で,鯨肉への評価は高まっていったように思われる。室町期の《四条流庖丁書》が,何よりも鯉を高貴な食物としながら,鯨だけは別格で,その上位においてもよいとしているのも,その現れではないかと思う。鯨は魚だと認識されていたから,その食用には抵抗がなかった。しかも,味は魚や鳥より濃密であることが多い。脂肪とタンパク質不足の食生活の中で,鯨が歓迎されたゆえんであろう。近世まで鯨は荒巻(あらまき)にして中央へ送られることが多く,肉は汁の実,刺身,あえ物など,かぶら骨はなますやあえ物,内臓はいろいろに調理して賞味された。天保3年(1832)刊の《鯨肉調味方(げいにくちようみほう)》は鯨の部位のすべてについてその食味と調理を記した奇書で,小山田与清の筆になるとされる。もっとも美味とされる〈尾の身(おのみ)〉は尾に近い部分の背肉で,霜降り状に脂があって軟らかく,刺身として珍重される。尾びれの部分は尾羽(おば),尾羽毛(おばけ)/(おばいけ)などと呼ぶ。黒い皮に白い脂肪層をつけたまま売られたので皮鯨(かわくじら)ともいった。これを塩漬したのが塩鯨,脂肪層を薄切りにして熱湯でゆがき冷水にさらしたのがさらし鯨である。さらし鯨は,畝と呼ばれる腹部の部分でもつくり,からし酢みそあえなどにして喜ばれる。背の赤身肉は付け焼き,みそ漬,ステーキ,カツレツなどに用いられ,松浦漬と呼ばれるかぶら骨のかす漬は佐賀県の名産として知られる。
執筆者:鈴木 晋一
信仰と捕鯨組織
海にすむ最大の動物としての鯨は,各地の民族にいろんな意味で重要な印象を与えてきた。これが一方では宗教的な畏敬とか恐怖を与え,他方ではこれを対象とする壮大な漁労活動を促すことになる。ただ,それが人間と生活の場をともにする陸上の生物ではないため,これを直接自分たちの祖先として信仰するような例は多くないが,むしろ外界から人間の世界に禍福(かふく)をもたらすものとして注目を集めた場合が多い。北太平洋,北大西洋では鯨が陸地に接近することもあるので,例えば北西インディアンやエスキモーの美術にはこれを象徴化した図柄が見られる。エスキモーやチュクチ族はアイヌの熊祭に似た鯨祭を行う。日本では紀州の和田氏が支配した太地を中心に捕鯨専業の村ができたりした。鯨の漁獲は一般に豊漁とか富をもたらすものとされ,村をあげてお祭り騒ぎをするが,それは日常的な漁労活動ではなく,年に何度もない特別な行事となるからである。この鯨を捕獲するためには多人数の協力を必要とするので,〈鯨組〉というような社会組織が構成されて,これが特別な漁労活動を支えることにもなる。一方,沿岸に近づく鯨の数が減少すると,それに反比例して,鯨を求めて遠洋に出漁する回数が増加することになる。近世,ヨーロッパの漁民が大西洋や太平洋に出漁した中には,whalerと呼ばれる捕鯨業者の活動も顕著であり,これがいくつかの新発見をもたらしている。ボニン諸島と呼ばれる小笠原諸島にもそのような捕鯨業者の来住定着が見られた。日本でも〈鯨組〉の伝統をそのままに,五島には明治初年に早くも捕鯨会社が創設されており,昭和初期の日本最初の南氷洋捕鯨にも五島出身者が多かった。
執筆者:大島 襄二
民俗
古くは魚類とみなされ,漢字でも魚偏を用い,日本でもその捕獲法や食法を記した書物は《勇魚取絵詞(いさなとりえことば)》と題されている。鯨が魚類に属さぬことを初めて明らかにしたのは,1760年(宝暦10)出版の山瀬春政著《鯨志》である。沿海部では古くから浅海に寄って行動の自由を失ったものを捕獲利用したらしく,遺跡から骨が出土する。巨大なためと小魚の群れを追ってくるため漁民から神聖視され,豊漁の兆しとしてエビスサマ,エビスクジラなどと呼ぶ土地もある。回遊性があるため海岸に大きな神社のある土地では,定期的に現れるこの動物が神もうでをするなどといって捕獲をしなかった。食用,採油,ひげの利用など,身体各部がすべて利用されるので,近世には各地に捕鯨地が発生し,〈1頭捕れば七浦が盛える〉とまでいわれた。その方式は外国に類例のない網を使用する大規模なもので,洋式捕鯨導入まで続いた。鯨は神聖視されたのと,母子の愛情が深いこととが浦人の心をとらえ,墓を建ててこれを供養する風習も広がった。
執筆者:千葉 徳爾
シンボリズム
古くから海の怪物と呼ばれてきたクラーケン,リバイアサン,摩竭魚(まかつぎよ)などについて,その正体を鯨とする説がある。またキリスト教伝説でも,旧約聖書の《ヨナ書》に出てくるヨナをのみこんだ大魚は鯨とされるが,多くの場合サメやワニあるいはイルカとの混同と思われる。ただしアリストテレスの《動物誌》や大プリニウスの《博物誌》に空気呼吸を行う事実など相当詳細な記述がなされており,古代の地中海世界でも鯨が知られていたことは確実である。中世のベスティアリ(動物寓意譚)では芳香を放って獲物を口もとへおびき寄せる生物とされ,キリスト教の象徴としては3日3晩鯨の腹にいたヨナの物語にちなみキリストとその復活を表す。現代のユング心理学では,鯨にのまれたあとに吐き出される過程を魂の死と再生の隠喩(いんゆ)と解釈している。またその巨大な口は地獄への門,その腹は地獄あるいは墓にたとえられ,悪魔の化身ともみなされた。このイメージは図像化され,大口をあけて民衆をのみこむ魔王(あるいはドラゴン)を描いたキリスト教の地獄絵を成立させたといわれる。船乗りにとっては強暴な自然のシンボルで,古地図には危険な外海を示すものとして盛んに描き込まれ,またメルビルの《白鯨》においては宇宙の破壊力が鯨に托されている。中世以降は鯨の生態が知られはじめ,その摂食行動から貪欲(どんよく)を,また巨大な生殖器から激しい性欲を象徴する生物という意味合いが強まった。
執筆者:荒俣 宏