デジタル大辞泉 「腎臓」の意味・読み・例文・類語
じん‐ぞう〔‐ザウ〕【腎臓】
Sponserd by ![]()
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
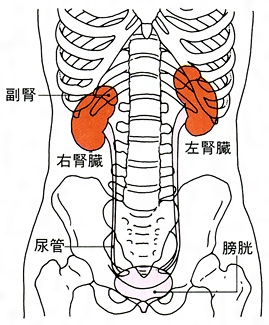
泌尿器系の器官で、血液の一種の濾過(ろか)器の機能を果たす。腎臓は、腹腔(ふくくう)の後上部壁で脊柱(せきちゅう)の両側に位置する。左右1対の腎臓は、ともにその前面だけが後腹壁を覆う壁側腹膜によって覆われているため、腹腔外部に存在する腹膜後器官とされる。
[嶋井和世]
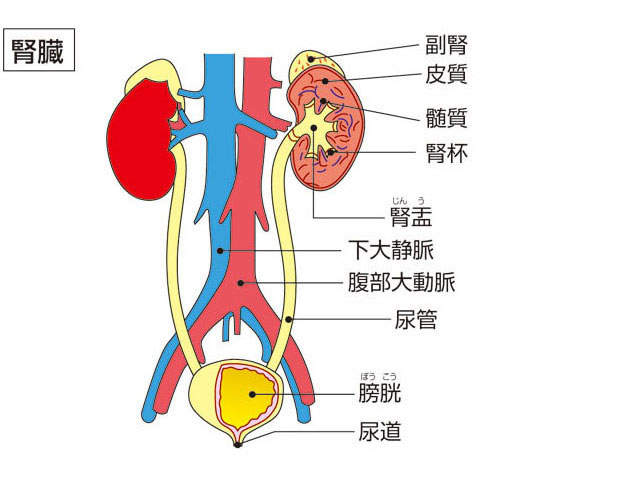
両腎ともにソラマメに似た形状で、くぼみのある部分が内側を向いている。平均の大きさは長さ10センチメートル、幅5センチメートル、厚さ3センチメートルで、重量は100グラムほどである。女性に比べると、男性がやや大きい。後腹壁における位置は左腎臓が第11胸椎(きょうつい)から第3腰椎にわたる高さにあり、右腎臓はそれより半椎体か1椎体分下がる。両腎の長軸はその延長線が上方で交差する。つまり、長軸は下方でやや開いた形となる。色調はやや暗紅色を呈し、外形では上・下端、前・後面、内・外側縁を区別できる。腎臓は上端に付着している副腎(腎上体)とともに脂肪組織に包まれており、これを脂肪被膜とよぶ。脂肪被膜は年齢とともに厚くなる傾向があるが老人になると減少する。腎臓の表面は直接、線維性の被膜で覆われ、これを線維被膜とよぶ。正常な腎臓ではこの被膜は容易にはがすことができるが、炎症のあとなどでは癒着のため、はがすことができなくなる。腎臓の内側縁の中央にあるくぼみは腎門といい、腎臓への動・静脈、リンパ管、神経、尿管が出入する。腎臓と周囲臓器との位置的関係をみると、右腎臓では前面に肝臓と、下方の一部に結腸が接し、内側は十二指腸が接触している。左腎臓の場合は、前面上部に脾臓(ひぞう)、一部は胃底に接し、下方では空腸、下行結腸などが接している。
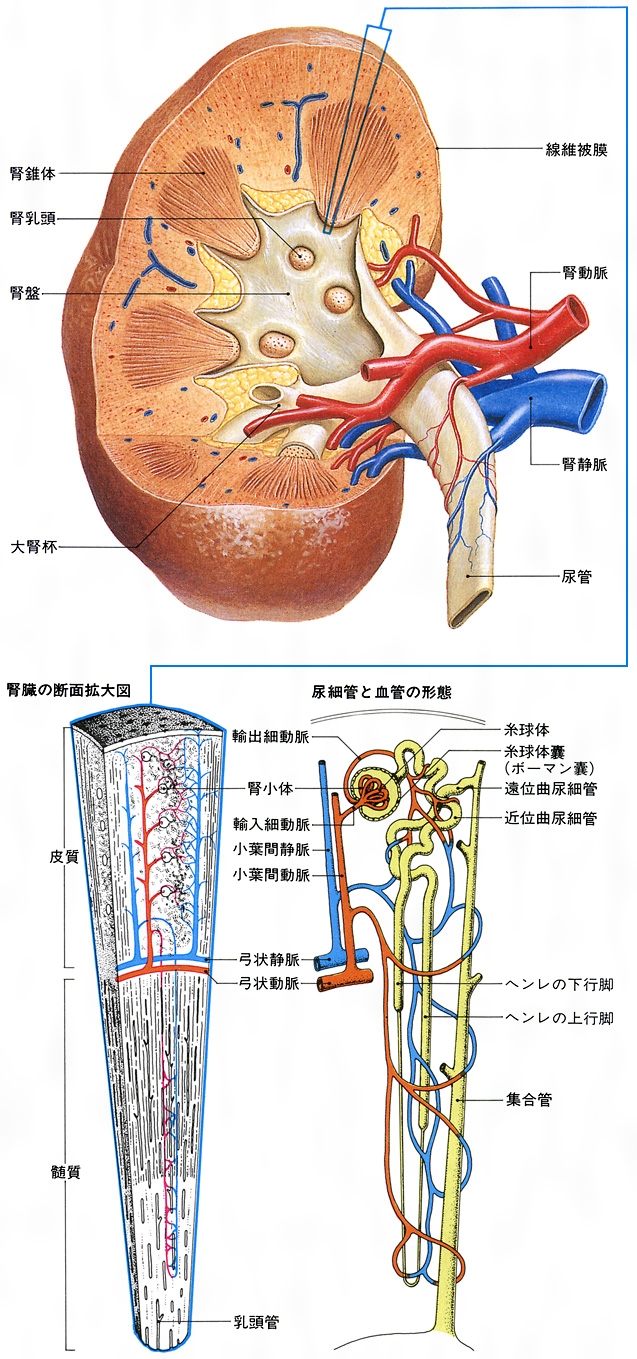
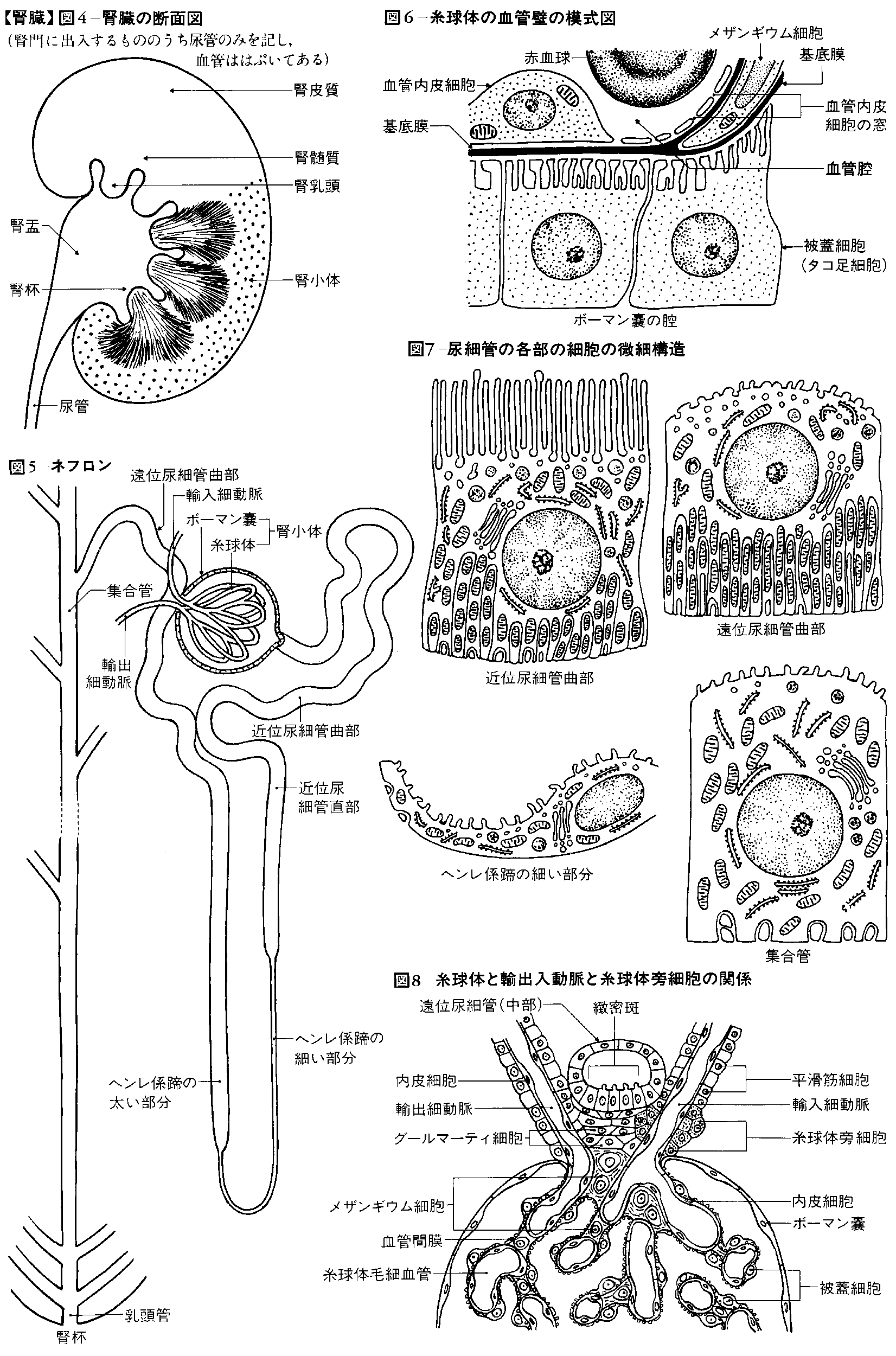
腎臓を外側縁から腎門に向かって縦断してみると、腎門の内部に広い腔があり、この腔は尿管に続いている。この腔所が腎盤(腎盂(じんう))とよばれる部分で、尿を導く部屋である。腎盤を取り囲むようにして腎臓実質がある。実質の割面では外層にあたる皮質と内層にあたる髄質とが区別できる。皮質の割面は暗赤褐色で、顆粒(かりゅう)状にざらざらしている。髄質には、先端を腎盤に向けた7~10個ほどの腎錐体(すいたい)が放射状に配列している。すなわち、錐体の底面は腎表面の方向に向いていることになる。腎錐体の突出部は腎乳頭とよび、100~700個くらいの腎乳頭口が開口している。腎乳頭が突出している腔はとくに腎杯(じんぱい)とよぶ。腎杯は腎乳頭を杯(さかずき)形に囲んでいるが、1~2個の乳頭が開く腔を小腎杯とよび、2~4個の小腎杯が集まって大腎杯となる。大腎杯は合流して腎盤に開いている。腎盤は尿管に移行する。
腎皮質内には尿を産生する腎小体が集合している。腎皮質が肉眼的にも顆粒状で赤褐色にみえるのはこの腎小体のためである。腎小体の大きさは直径200マイクロメートルほどで、毛細血管網を主体とした塊であるが、糸球体とこれを包む二重の袋状の糸球体嚢(のう)(ボーマン嚢、ボウマン嚢)とからなる。なお、1個の腎臓には100万~300万個の腎小体があるといわれる。糸球体は複雑に分岐した毛細血管網の塊で、この塊を包むのが糸球体嚢である。この嚢は内葉と外葉とによって内腔をつくっている二重膜の嚢である。この糸球体において毛細血管内の血液は濾過されるが、その後、濾過された原尿は糸球体嚢の狭い腔にためられ、糸球体嚢の輸入管口・輸出管口(糸球体嚢内で糸球体を形成するための輸入細動脈・輸出細動脈の出入口)のある血管極とは反対側に存在する尿細管極の尿細管の入口に流れる。この部分の尿細管は尿が流れ出る最初の細管で、腎小体の近くで迂曲(うきょく)して走るので、近位曲尿細管(あるいは近位迂曲管)の名がある。近位曲尿細管は皮質内を迂曲して走りながら髄質に下り、髄質に入ると急に細くなって下行脚(ヘンレの下行脚。19世紀のドイツの解剖学者F. G. J. Henleにちなむ)となる。下行脚は髄質中でU字形に反転してヘンレ係蹄(けいてい)(わな)をつくり、上行脚(ヘンレの上行脚)となってふたたび皮質に向かう。上行脚は皮質内でふたたび太くなり、屈曲走行する。この部分を遠位曲尿細管(あるいは遠位迂曲管)とよび、近位曲尿細管の部分に介在して走っている。遠位曲尿細管はふたたび髄質に向かって走り、皮質、髄質の境あたりで集合管となる。集合管は互いに結合して太くなり(径30~50マイクロメートル)、髄質内側部に入ってさらに太い乳頭管(径200~300マイクロメートル)となり、腎乳頭に至り、腎乳頭尖端(せんたん)の乳頭孔に開口する。腎小体とそれから出る1本の尿細管のことをネフロン(腎単位)とよぶ。1本の集合管には多数のネフロンが連なることになる。
[嶋井和世]
腎臓には、腹大動脈から左右両側に直角に分岐する腎動脈(左・右腎動脈)が腎門を経て腎臓内に進入する。右腎動脈が左腎動脈よりやや長い。腎動脈は腎臓内に入ると腎門で数枝に分かれ、葉間動脈となり、腎乳頭の間を走って皮質に向かう。葉間動脈は皮質と髄質との境でさらに小葉間動脈となり、腎表層方向に向かう。この走行の間に枝が出て輸入細動脈(輸入管)として腎小体に入る。この血管が糸球体を形成する。糸球体は輸出細動脈(輸出管)を出し、これはふたたび毛細血管となって皮質内の尿細管を取り囲み、ついで小葉間静脈に集まり、さらに弓状静脈から葉間静脈に入る。葉間静脈は合流して腎静脈となる。
腎臓の動脈の特徴は終動脈であるとされる。つまり、腎臓に入って数本に分岐したあとの動脈は、それぞれの分岐枝の間に吻合(ふんごう)をもたないため、各分岐動脈枝の分布区域は限定されることになる。この動脈枝を区(域)動脈とよび、区(域)動脈の分布する区域を腎区域とよぶ。解剖学上では上区、上前区、下前区、下区、後区などに区別しているが、一つの区(域)動脈に閉塞(へいそく)が生じると、その動脈の分布区域は組織壊死(えし)に陥る。
腎臓への神経は腹大動脈や腎動脈周囲の神経叢(そう)(腎神経叢)からの神経線維が交感性線維として分布するが、迷走神経から由来する副交感神経も分布する。さらに、腎臓には知覚神経も分布している。
腎臓の形態の異常のうち、もっとも発生頻度が高いのが馬蹄腎(ばていじん)である。これは、左右の腎臓が下端で融合し、馬蹄形を呈するもので、腎実質がつながっている場合と線維性組織のみでつながっている場合とがある。そのほか、左右いずれかの腎臓が先天的に欠損している単腎症、腎臓全体が発育不良の発育不全腎、あるいは骨盤部や胸部に位置の変位をおこした骨盤腎、胸部腎などの位置異常もある。
[嶋井和世]
体内の不要な不揮発性・水溶性の代謝産物または有害物を体外に排除する働きを排泄(はいせつ)というが、腎臓はもっとも重要な排泄器である。腎臓が果たす尿の排泄によって、(1)血液中の不要産物・有害物の除去、(2)血液の浸透圧調節、(3)細胞外液量調節、(4)血液のpH調節、(5)血漿(けっしょう)組成の調節、などが可能となる。また、腎臓からは数種類のホルモンが分泌され、腎臓は内分泌器官としての働きももっている。このようにみると、腎臓は、血液の性状を一定に保ち、ひいては身体の内部環境としての体液の恒常性を維持している器官といえる。腎臓は汚物処理場に例えることもできる。汚物処理場は、われわれの日常生活において生じるさまざまな汚物や塵芥(じんかい)を下水道などによって集め、焼却したり化学的処理を施してそれらを無害な形に変えているが、腎臓も、血液によって運ばれてきたさまざまな老廃物を取り出して尿として体外に捨てる(処理する)機能をもっている。汚物処理場がなければ健康的な生活を営むことができなくなると同様に、腎臓が正常に機能しないと血液中に老廃物がたまり、体液の恒常性を維持することができなくなってしまう。このような状態を尿毒症とよび、重症の場合は死に至る。また近年は塵芥をただ焼却処理するだけではなく、そのなかから再利用できるものを選別して省エネルギー・省資源の一助とすることが検討・実施されているが、腎臓はこの点では現代の汚物処理場よりもはるかに効果的に働いている。すなわち、血液からいったん取り出した物質をそのまま尿として体外に捨ててしまうのではなく、そのなかから利用できるものを再吸収して血液中に戻すという仕事も行っているのである。
[真島英信]
腎臓における尿生成の最初の段階は腎小体において行われる。ボーマン嚢の内部において輸入細動脈は毛細血管となって糸球体を形成するが、ここでは血管壁に直径50~100ナノメートルの小孔が多数認められる。これらの小孔を通して血漿のうちタンパク質を除いた成分が濾過されて糸球体からボーマン嚢に出て糸球体濾液(原尿)となる。こうした腎小体における濾過は血液中の分子の大小、および相対的圧力の関係だけに支配される現象であると考えられている。したがって、タンパク質でも分子量の小さい卵白アルブミン(分子量4万)やヘモグロビン(分子量6万8000)は濾過されて出るし、ときにはアルブミンのような大きな分子がそのまま濾過されることもありうると考えられる。実際に健康な人でも疲労すると尿中にアルブミンが検出されることがある。ただし分子の大きい物質ほど濾過速度は遅くなる。
腎臓に広く分布する糸球体から1分間に濾過されて尿細管に出る濾液の量を「糸球体濾過量」といい、腎臓の機能を表す一つの指標となっている。糸球体濾過量の値は1分間に男子で110ミリリットル、女子で100ミリリットル程度である。腎小体における濾過の決定要因としての相対的圧力に関与するのは、血圧、ボーマン嚢の内圧、および血漿の膠質(こうしつ)浸透圧である。なお、ここでいう血圧とは輸入細動脈の血圧と輸出細動脈の血圧との差であり、通常は約70ミリメートル水銀柱である。また、ボーマン嚢の内圧は条件によりいろいろに変化するが、平均約20ミリメートル水銀柱、血漿の膠質浸透圧は約20ミリメートル水銀柱である。濾過の原動力となる濾過圧は、血圧からボーマン嚢の内圧および膠質浸透圧を差し引いた値として求めることができる。したがって、濾過圧は通常70-(20+20)、すなわち30ミリメートル水銀柱程度となる。なお、糸球体における濾過においては、次のような現象が認められる。(1)糸球体毛細血管の血圧または血流量が増加すれば、糸球体濾過量は増加する。(2)糸球体の血圧は輸入細動脈の収縮によって低下し、輸出細動脈の収縮によって上昇する。(3)輸入細動脈の平滑筋は交感神経刺激により、あるいは大量のアドレナリンによって収縮し、カフェインなどによって拡張するが、拡張すると糸球体血圧は上昇し、糸球体の濾過量が増すので尿量は増加する。(4)輸出細動脈はレニン、ヒスタミン、少量のアドレナリンによって収縮するので、この場合も糸球体血圧が上昇し、尿量は増加する。
腎臓の血流量は皮膚温との関係が深い。すなわち、寒冷のために皮膚温が低下すると、体温調節反射によって皮膚血管が収縮し、発汗が抑制されるが、このとき代償的に腎臓の血管が拡張して尿量が増す。
[真島英信]
腎小体において濾過される糸球体濾液は1日に約160リットルにも達する。この糸球体濾液の成分は血漿からタンパク質のみを除いたものに等しく、その中には各種のイオン、ブドウ糖、アミノ酸など利用価値のあるものも多量に含まれているため、これをそのまま尿として排泄してしまったのでは非常な浪費となる。さらに毎日160リットルもの尿を排泄するとしたならば、それに等しい量の水を飲まなくてはならず、1日が水を飲むこととトイレに通うことだけで終わってしまうことになる。このようなむだを防ぐために、糸球体濾液がボーマン嚢に続く尿細管・集合管を流れ下る間に、ある物質は再吸収され、またある物質は血液から分泌されて最終的には尿となって排泄される。さらに、尿細管・集合管の周囲を糸球体からの輸出細動脈がふたたび毛細血管となって取り囲むため、尿細管・集合管との間で物質の再吸収・分泌が行われる。近位曲尿細管ではナトリウム、カリウムなどの電解質、ブドウ糖、フルクトース、アミノ酸、アスコルビン酸、尿素などが再吸収され、これに伴って水の大部分が受動的に再吸収される。一方、パラアミノ馬尿酸、ペニシリンなどの異物やクレアチニンなどは分泌される。こうした尿細管における再吸収・分泌は、単なる濃度勾配(こうばい)にしたがって物質が移動する受動的なものばかりではなく、尿細管の細胞が積極的にエネルギーを消費して濃度勾配に逆らって特定の物質を移動させる場合も多い。後者のような物質の移動を能動輸送という。たとえばヘンレ係蹄の太い上行脚では塩素が能動輸送によって再吸収される。また遠位曲尿細管ではナトリウム、重炭酸イオン、および水が再吸収され、カリウム、水素、アンモニアなどが分泌される。
腎動脈から腎臓に流入する血液量は1分間に1.2~1.3リットルで、心臓から1分間に拍出される血液量の約25%に相当する。このうち糸球体・尿細管を含めて腎臓において濾過に関与する血液量を「有効腎血流量」という。有効腎血流量は男子では1分間に約1リットル、女子では約0.8リットルである。この血流量からまったく濾過されない赤血球の占める体積を除いた値(すなわち血漿の容積)を求めたものを「腎血漿流量」とよぶが、これは腎臓の働きを知る一つの指標として用いられる。また、糸球体濾過量と腎血漿流量との比は濾過率とよばれ、通常は約0.2である。すなわち糸球体を流れる血漿のうち約20%が濾過されて尿細管に出ることになる。
[真島英信]
尿の大部分は水であるから、尿量は水の排泄量とみなすこともできる。糸球体濾過量が1日約160リットルに及ぶことはすでに述べたが、これに対して1日の尿量は約1.5リットルにすぎない。このことから、濾過された水の99%以上は再吸収されるわけであり、尿はそれだけ濃縮されていることになる。ヒトでは1日に最低500ミリリットルの尿で体液の恒常性を維持することができる。このときの尿は高度に濃縮されていて、その浸透圧は血漿の約5倍もある。また、尿量の最大限値は約23リットルであり、この場合の尿は、浸透圧が血漿の約10分の1という希薄な尿として排泄されるが、溶質の量は変化しない。すなわち、腎臓は溶質の排泄量を変えることなく、水の排泄量を大幅に変化させる能力をもっているわけである。糸球体から濾過された糸球体濾液の水分のうち、約75%は近位曲尿細管で再吸収され、また、5%はヘンレ係蹄に存在する対向流増幅機序という独得の仕組みによって生じる濃度勾配にしたがって再吸収される。さらに遠位曲尿細管で15%、集合管で約4%以上の水が再吸収され、合計すると結局99%以上が再吸収されることとなる。尿細管壁はヘンレ係蹄上行脚を除いてつねに水をよく透過するが、集合管壁の水に対する透過性は、下垂体後葉から分泌される抗利尿ホルモンの影響によって変化を受ける。抗利尿ホルモンが分泌されると、水はナトリウムの移動に伴って集合管壁から再吸収され、尿量は減少する。逆に、たとえば多量の水を飲んだときには抗利尿ホルモンの分泌が抑制されるため、集合管壁の水透過性が低下して水の再吸収が減少する。その結果、希薄な尿が多量に排泄されることになる。なお、下垂体の腫瘍(しゅよう)などによって抗利尿ホルモンの分泌が障害されると、希薄な尿が異常に増加し、1日の尿量は8~12リットルに及ぶ。さらに完全な抗利尿ホルモン欠乏の場合では、尿量は1日に23リットルにも達することがある(尿崩症)。
ナトリウムは尿細管および集合管の全域にわたって受動的あるいは能動的に再吸収されるため、糸球体濾液中のナトリウムの96~99%は回収される。全再吸収量の約85%は近位曲尿細管で行われ、残りはそれ以外で再吸収される。つまり、1日のナトリウム排泄量は結局1日のナトリウム摂取量とほぼ等しくなるように調節されていることになる。ナトリウムの大部分は塩素を伴って再吸収されるが、一部は水素やカリウムと交換輸送される。副腎皮質から分泌される電解質コルチコイド(アルドステロンなど)や糖質コルチコイドは、ナトリウムの交換輸送および塩素を伴う再吸収を促進するが、その作用機序にはまだ不明な点が多い。体液(細胞外液)の浸透圧は水の量とナトリウムの量との相対的な関係によって維持されている。腎臓はこの水とナトリウムとの排泄量を調節することによって細胞外液の浸透圧を一定に保つように働いている。さらに腎臓のこのような働きを調節しているのが、すでに述べたように、水に関しては抗利尿ホルモン、ナトリウムに関してはアルドステロンなどである。たとえば、いま細胞外液の浸透圧が上昇したとすると、下垂体後葉が刺激されて抗利尿ホルモンの分泌が増加する。その結果、水の再吸収が増加し、尿量が減少して浸透圧の上昇を防ぐ。同時に渇きを催して水を摂取する飲行動に駆り立てられることになる。一方、尿細管のナトリウム排泄機能が障害されるとナトリウムが体内に貯留し、それに伴って水も貯留する結果、細胞外液、とくに間質液量が増して体重増加を伴う全身的浮腫(ふしゅ)を生じる。
ブドウ糖は近位曲尿細管においてほぼ完全に再吸収される。その再吸収の機序は、小腸におけるブドウ糖の吸収機序によく似た能動輸送である。ただし、その再吸収の能力には限界があり、血液中のブドウ糖濃度がある限度以上にまで上昇して濾過されるブドウ糖が多量になると、尿細管において再吸収しきれなくなり、尿中にブドウ糖が排泄されるようになる。これが糖尿である。糖尿を生じるようになる血液中のブドウ糖濃度は、血液1デシリットル当り200~250ミリグラム以上であり、これは正常値の2~2.5倍である。糖尿は糖尿病患者に一般に認められるが、健康者でも糖質を多量に摂取した直後には尿中にブドウ糖が検出されることもある。
[真島英信]
腎臓は体液のpHの調節にも関与している。血液のpHは重炭酸イオンHCO3-の濃度と血液中の二酸化炭素CO2の濃度との比によって決まる。いま、もし水素イオンH+が増すとすると重炭酸イオンは減少して二酸化炭素が増加する。その結果pHは低下するが、増加した二酸化炭素は肺から呼気として排出され、一方の水素イオンは尿細管から尿中へと排泄されていくので、結局血液のpHは一定に保たれることになる。また、腎臓は重炭酸イオンや不揮発性の酸性物質の排泄も調節して、血液のpHを一定に保つように働いている。このように血液のpHの変動は極力抑えられるような仕組みをもっているが、種々の原因によって血液のpHが酸性に、あるいはアルカリ性に変動してしまうことがある。血液のpHが酸性になった状態をアシドーシス、アルカリ性になった状態をアルカローシスとよぶが、これらの症状は、さらにその原因によって、呼吸性および代謝性に分類される。たとえば糖尿病のような代謝においては、腎臓の排泄機能を超えるほどの酸性物質が体内にたまってしまうことがあり、体液は酸性となる。このような状態を代謝性アシドーシスという。代謝性アシドーシスの場合は、腎臓が水素イオンの排泄を亢進(こうしん)させると同時にナトリウムや重炭酸イオンの再吸収を促進して、塩基の減少を極力防ぐ役割を果たす。さらにpHの低下によって呼吸中枢が刺激されて肺における換気が促進され、多量の二酸化炭素を呼び出してpHを元に戻すように働くこととなる。逆に呼吸困難などのために肺における換気が十分に行えなくなると、血液中に二酸化炭素が貯留して体液は酸性となる。この状態が呼吸性アシドーシスである。この場合は、腎臓において酸分泌と重炭酸イオン再吸収の速度が増してpHを是正するように働くわけである。同様にして代謝性アルカローシスの場合は換気の抑制が、呼吸性アルカローシスの場合は腎臓における酸分泌と重炭酸イオン再吸収が抑制される。このように肺と腎臓は共同して血液のpHを一定に保つように働いている。
[真島英信]
これまで述べたように、腎臓は体内に生じた老廃物を排除するのみならず、血液、ひいては細胞内液をも含めた全体液の浸透圧、pHを一定に保つように働いている器官である。腎臓の排泄機能が生体の要求に十分応じられなくなった状態を腎不全といい、種々の重篤な症状を呈するようになる。腎不全の場合は尿量が減少するが、逆に尿量が増加する場合を利尿という。利尿の原因として次の三つがあげられる。
(1)抗利尿ホルモンの分泌抑制 多量の水を飲むと尿量が増加する。この機序は、全血液量の増加(血圧上昇)と血液浸透圧の低下によって直接に糸球体濾過量が増加することも考えられるが、それよりも血圧上昇や浸透圧低下に応じて抗利尿ホルモンの分泌が抑制され、水の再吸収が減少するためであると考えられる。この場合、尿の浸透圧は低くなる。その理由は、尿細管における電解質の再吸収が抑制されないためである。
(2)浸透圧的利尿 マンニット(マンニトール)のように浸透圧が大きく、しかも再吸収されにくい物質を血中に入れると、近位曲尿細管でこの物質は再吸収されないため、水の受動的再吸収が妨げられ、その結果尿量が増す。食塩、ブドウ糖などでも大量に与えればいずれも同様の機序によって利尿をきたす。糖尿病のときに尿量が増すのも、尿中へブドウ糖が多量に濾過されて出るための浸透圧利尿にほかならない。
(3)利尿剤 水銀利尿剤は、尿細管細胞による能動的な電解質の再吸収機能を抑制することによって尿量の増大をきたすと考えられる。この場合尿は酸性となり、血液中の塩素は減少してアルカローシスに傾く。カフェインなどのキサンチン誘導体は主として腎血管の拡張、腎血流量の増大によって糸球体濾過量を増加させる。また、ジギタリスなどの強心剤はその強心作用によって心臓からの拍出量の増大、血圧の上昇によって糸球体濾過量を増加させ利尿をきたす。
[真島英信]
腎臓は内分泌器官としての働きももっている。糸球体の輸入細動脈の特別な部位には血圧に反応する部分があり、腎臓の血圧または血流量が減少すると、その部分の糸球体旁(ぼう)細胞(輸入細動脈壁の中膜の平滑筋細胞が特殊化したもの)が刺激されてレニンとよばれる一種のホルモンが分泌される。また、尿細管のナトリウム濃度が低下した場合には、糸球体旁細胞に接する緻密斑(ちみつはん)の細胞(遠位曲尿細管は、血管極の近くでかならず1回糸球体に接するが、この部分の尿細管上皮細胞をいう)からも遊離される。レニンは血漿タンパク質中のアンギオテンシノーゲンを活性化してアンギオテンシンⅠとするが、アンギオテンシンⅠはさらに血液中の酵素の作用によってアンギオテンシンⅡとなる。アンギオテンシンⅡは強い血管収縮作用をもっているため、血圧上昇をきたすと同時に、副腎皮質に作用してアルドステロンおよび糖質コルチコイドの分泌を促進する。アンギオテンシンⅡの分解産物であるアンギオテンシンⅢは血管収縮作用は弱いが、アルドステロン分泌促進作用はⅡと同程度に強い。
交感神経刺激またはカテコールアミンはレニン分泌を促進し、アンギオテンシンⅡおよび抗利尿ホルモンはレニン分泌を抑制する。レニンは腎臓の血圧が低下したときに分泌され、その結果として血圧を上昇させるのであるから、このような昇圧物質は腎血流の自己調節的意味を有すると考えられる。しかし、動脈硬化、血栓などによって腎動脈に狭窄(きょうさく)を生じた場合も、狭窄部より下流にある腎臓の血圧が低下するためにレニンが分泌される。レニンはアンギオテンシンを介して全身の血管を収縮させ、血圧を上昇させるが、腎動脈はすでに機械的に狭窄されているので腎臓の血圧はほとんど上昇しない。このような場合はレニンの分泌増加が持続する結果、慢性的な高血圧症(腎性高血圧)となる。なお、腎臓からはレニン以外の組成の不明な物質が生成され、末梢(まっしょう)血管を収縮させている。また、腎臓では他の器官でつくられた昇圧物質の破壊が行われるので、腎臓を摘出すると血圧の上昇がおこる。
腎臓への酸素供給量が低下したり、血液中のヘモグロビン濃度が減少すると、エリスロゲニンとよばれるホルモンが腎臓から分泌される。このホルモンの分泌細胞はまだよくわかっていないが糸球体細胞であろうと考えられている。エリスロゲニンは血液中の基質に作用してプロエリスロポイエチンを合成し、さらに肝臓においてエリスロポイエチンとなる。エリスロポイエチンは赤血球の新生を促進する作用をもっている。すなわち、骨髄の幹細胞の分化を促進し、赤血球前駆細胞を増加させるとともに、赤芽球の細胞分裂を促進し、網状赤血球の血液中への放出も早める。また細胞当りのヘモグロビン合成量を高める作用もある。
[真島英信]
前述の腎不全をはじめ、腎臓の片側にみられる腎盂炎、腎結石、水腎症、腎結核、腎腫瘍(しゅよう)など、また両側にみられる腎炎、ネフローゼ、腎硬化症などのほか、腎損傷や遊走腎などがある。
[真島英信]
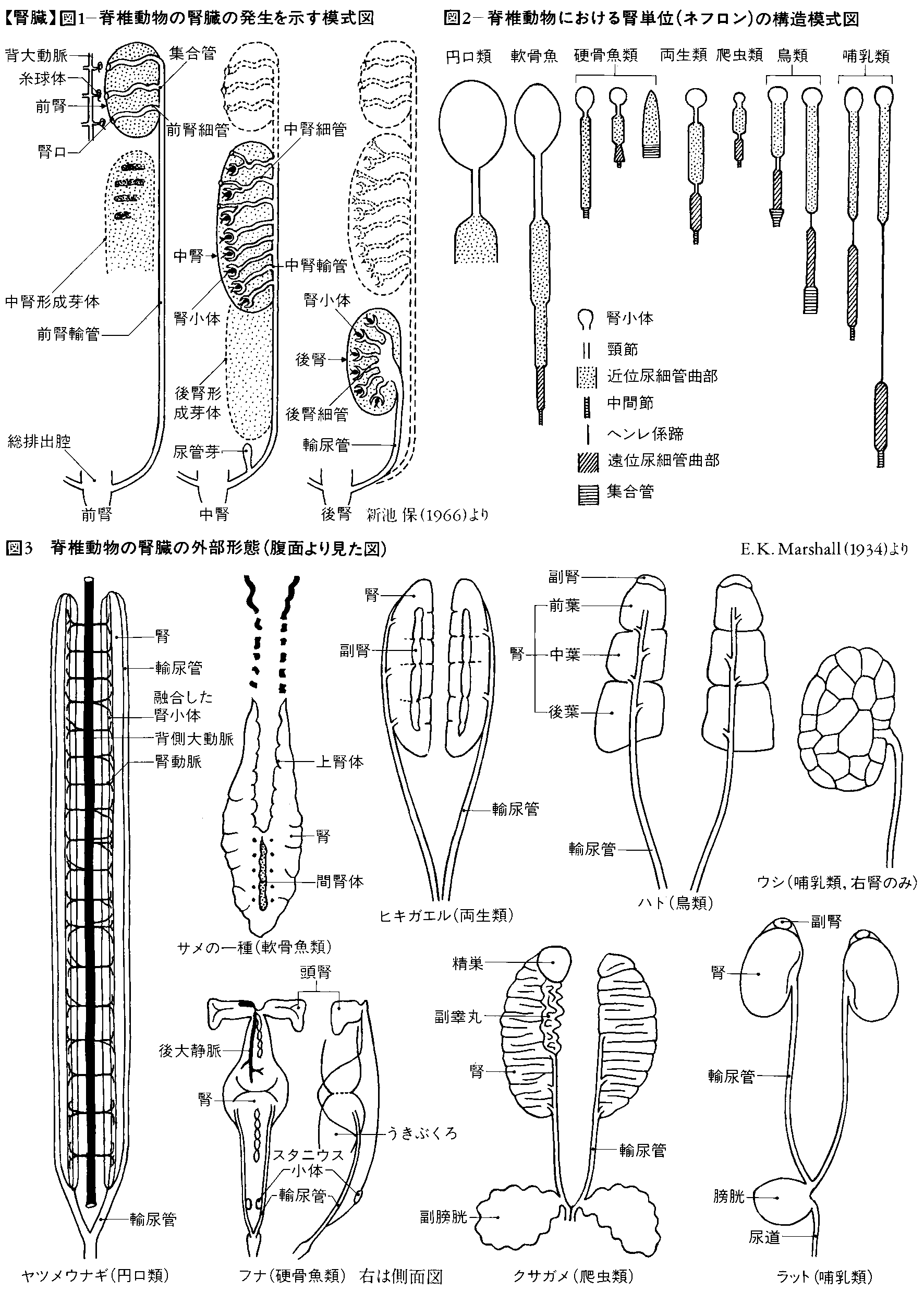
腎臓は脊椎(せきつい)動物にみられる泌尿(排出)器官で、中胚葉(ちゅうはいよう)の腎節に由来する。系統発生的にも個体発生的にも、前腎、中腎、後腎の三つに分けられる。前腎はすべての脊椎動物の胚期に現れ、前腎小管とよばれる腎管と、その近くの糸球体とからなる。前腎の形で終生働くのは円口類(ヤツメウナギ)だけで、魚類や両生類では幼生期にのみ働く。他の脊椎動物では、前腎は痕跡(こんせき)的にしか形成されず、やがて退化する。前腎に次いで中腎が現れる。魚類、両生類では中腎が終生腎として働く。体節状に配列した中腎小管は、体腔(たいこう)に開く部分から枝を出して糸球体を包み、ボーマン嚢(のう)をつくる。爬虫(はちゅう)類、鳥類、哺乳(ほにゅう)類では、中腎は胚期に一時的に出現するがやがて退化し、後腎が終生腎として働く。後腎の集合管の周囲には間葉性細胞が集まって尿細管をつくり、それぞれが糸球体と結合する。腎臓の糸球体により濾過(ろか)された状態の尿は原尿とよばれるが、続いて尿細管の上皮細胞により必要成分が再吸収されて尿となる。
[小林靖夫]
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
腎臓は脊椎動物に共通する排出器官で,老廃物,おもに窒素代謝産物の体外への放出がその主要な働きであるが,体液の浸透圧調節のための水分の排出,体液中の余剰成分の排出,体液を一定のpHに保つためアルカリ性または酸性物質の排出,有害あるいは不要物質の排出などの機能ももっており,これらの諸作用は互いに関連し合っている。
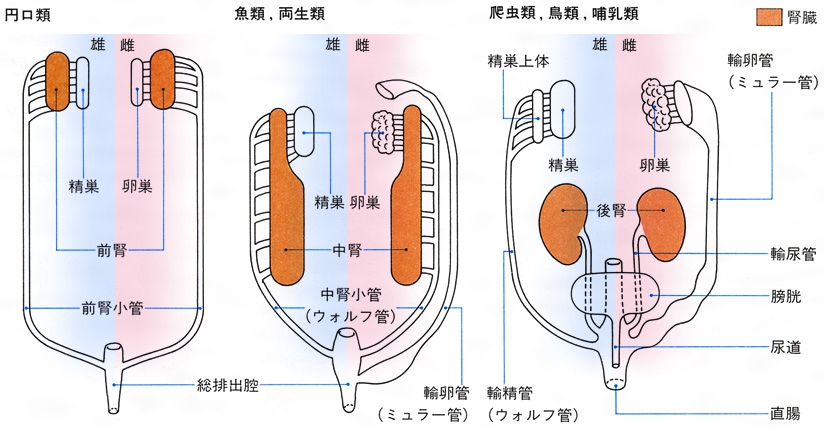
腎臓は腎管nephridiumに由来したものと考えられ,発生上の順序や位置から前腎,中腎および後腎に分けられる(図1)。最初に現れる前腎pronephrosは漏斗状となって体腔に開き,腎臓の原始型とみられる。円口類のメクラウナギでは前腎がその後に発達する中腎とともに機能を営むが,軟骨魚類と爬虫類以上では前腎が初めから退化痕跡的で作用をもたない。発生が進むにつれて前腎は退化し,その後方に中腎mesonephrosが生じる。中腎からは血管とつながった腎小体(マルピーギ小体)が形成される。円口類,軟骨魚類,硬骨魚類,両生類では中腎が生涯の腎臓として働く。爬虫類以上の脊椎動物では中腎も退化し,さらにその後方に後腎metanephrosが発達して,それが生涯の腎臓,すなわち最終腎として排出作用を営む。中腎が働きだすと不用になる前腎輸管はミュラー管Müllerian ductとも呼ばれ,雄では退化するが雌では存続して輸卵管となる。中腎輸管はウォルフ管Wolffian ductと呼ばれ,中腎が退化する爬虫類以上の動物の雄では輸精管となり,雌では退化する。
外形も動物により異なり,一般には左右1対であるが,軟骨魚類や硬骨魚類のように左右が融合したり,鳥類のように前・中・後葉と3葉に分葉したり,哺乳類の腎臓でも普通にみられるソラマメ型から多くの小腎からなる葉状腎をもつものまで多様である(図3)。
腎臓は多数の腎単位,すなわちネフロンnephronの集合したもので,腎単位は腎小体(ラテン名corpusculum renis,英名renal corpuscle)と尿細管(ラテン名tubulus renalis,英名renal tubule)とからなり,排出機能を営む一つの構造単位である。腎動脈血の供給を受けている糸球体とそれを包むようにしてボーマン囊Bowman's capsuleがあり,この両者をいっしょにして腎小体という。しかし海産硬骨魚では,腎小体の退化消失したものがある。尿細管は,腎小体に続いて頸節,近位尿細管曲部,遠位尿細管曲部および集合管の順で構成されている。海産の硬骨魚の尿細管には遠位尿細管曲部が存在しない。両生類,爬虫類,鳥類の尿細管では近位尿細管曲部と遠位尿細管曲部の間に中間節が見られ,哺乳類になるとヘンレ係蹄Henle's loopがこれに変わる(図2)。これらの尿細管の立体的配列も硬骨魚類などでは不規則であるが,両生類,爬虫類では規則的になり,鳥類,哺乳類ではより整然と配列しているので,尿細管の立体的配列構造から腎臓は皮質,髄質外層,髄質内層の3部に分けることができる。
尿生成機構の基本は,ろ過,再吸収,分泌の三つの作用である。脊椎動物の腎臓では,いったんろ過排出してそのなかから必要な物質を回収するといった,一見非能率とも思える方法で尿を生成しているが,これは脊椎動物が環境に適応しつつ長い過程で獲得したものにちがいない。腎小体ではろ過が行われ,ボーマン囊内の糸球体ろ過液である原尿の成分濃度は,タンパク質を除いて血漿(けつしよう)のそれとほぼ等しい。その後尿細管の各部位で,有用成分の再吸収や水の再吸収によって濃縮が起こり,さらに分泌も行われて終末尿が作られる。哺乳類ではヘンレ係蹄での対向流機構によって尿が濃縮されると考えられている。
このような腎単位の分化や機能は,それぞれの動物の環境適応,とくに浸透圧調節と深いかかわりがある。尿生成の最初の過程は糸球体でのろ過であり,体内に浸入した過剰の水を除く必要から,糸球体は淡水に生息する脊椎動物の腎臓に初めて発生したと考え,スミスH.W.Smithは脊椎動物の淡水起源説を提唱した。しかし,近年では血液中の2価イオン調節のため生じた器官で,脊椎動物もやはり海洋起源だとする考えが一般的である。
尿の生成には腎血流量,血圧,血中成分などの変動が関係し,その調節には神経やホルモンが複雑に関与している。なかでも,脳下垂体後葉ホルモンは,尿細管での水の再吸収を増加して尿量を減少させる。副腎皮質ホルモンのアルドステロンは,尿細管でのナトリウムの再吸収を増加させる。そのほか副甲状腺ホルモンは,リン酸塩とカルシウム代謝に関与し,プロラクチンは硬骨魚の浸透圧調節に関係して,淡水中で腎臓に働き,多量の薄い尿を作る。
→排出
執筆者:小川 瑞穂
腎臓は,腰椎の高さにおいて脊柱の左右に位置する1対のソラマメ型の器官であり,後腹壁にある脂肪組織(脂肪被膜)と繊維被膜に包まれている。腹膜には包まれていないで,それよりも後ろにある。つまり腹膜後器官である。大きさは,こぶし大よりやや大きく,成人男子で11cm×5.5cm,重量130g程度である。小児期から成人まで重量を増し,老化とともに減少する傾向にある。女子ではこれよりもやや小さく,また左右では右のほうが小さい。成人男子では,左腎は第11胸椎~第3腰椎の高さに位置するが,右腎は肝右葉に圧迫されるため,これよりやや低い。外側縁は鈍円状にふくらみ,内側縁は陥没してその中央に腎門があり,腎動脈,腎静脈,尿管,神経がここから出入りする。腎実質は,腎門から深くえぐりとられたようにへこみ腎洞を形成する。ここには,腎動静脈および尿管の上端がふくらんでできた腎盂(じんう)(腎盤)と,その間をうめる脂肪組織が存在する。腎盂は2~3個の大腎杯に分かれるが,それはさらに7~14個の小腎杯に分かれており,腎実質から突出している腎乳頭の先端に連結し,腎実質で生成された尿を受ける(図4)。
腎臓の断面を観察すると,表面は脂肪組織と繊維被膜に包まれており,実質のうち表面に近い部分を皮質,内部を髄質と呼ぶ。皮質は,原尿の生産装置である腎小体と,その回りにとぐろを巻く尿細管とから成り,髄質はおもに直行する尿細管と集合管と呼ばれるさらに太い管より成る。集合管は,最終的には,腎乳頭に集まり,乳頭を中心とする腎錐体という構造をつくる。また各腎錐体の間には皮質が入り込んでおり,これは腎柱と呼ばれている。
腎門から入った腎動脈は,腎洞から腎実質に達し,錐体間を走る葉間動脈に分枝する。これはさらに弓状動脈に分かれて皮質と髄質の間を弓状に走る。これから皮質へ小葉間動脈を出し,さらにそれらは輸入細動脈となって糸球体に入り,輸出細動脈となってこれから出,ついで皮質尿細管周囲の毛細血管網を経て,小葉間静脈,そして弓状静脈に注ぐ。また髄質には弓状動脈から直細動脈が出て毛細血管網となり,直細静脈を経て弓状静脈に注ぐ。これらの弓状静脈は葉間静脈へ注ぎ,さらに腎静脈に集められ,腎門より出る。腎動脈や腎静脈は腎臓の大きさに比べて大きい。これはこれらの血管が腎臓を養うためのみならず,何よりも腎臓で尿を作るために,ろ過するための血液を送っているからである。また,腎臓内部では尿生成のために血管がよく分岐し,毛細血管がきわめてよく発達している。
腎皮質中には,腎小体(マルピーギ小体。ラテン名corpusculum Malpighii,英名Malpighian corpuscle)と呼ばれる多くの球状構造が見られる。血液はここでろ過され,原尿となった後,これに続く尿細管部分において分泌および再吸収を受けて集合管に注ぐ。すなわち集合管は多くの尿細管を集める。ついで多数の集合管が腎乳頭に集まり乳頭管となり腎杯に開く。
一つの腎小体およびそれから出た尿細管は,集合管に注ぐまで,蛇行したり直行したり,あるいはその管の太さや構造を変えるものの,分岐することのない1本の管を形成している。この腎小体とそれに続く1本の尿細管の系を尿の生産と排出の機能単位・構造単位とみなして腎単位あるいはネフロンと呼んでいる。ネフロンは,片側の腎臓につき約100万といわれている(図5)。
尿の生成は,腎小体にはじまる。腎小体は直径約0.2mmの球状体で,血管の特殊な糸だまを丸いふくろで包んだ構造をしている。前者は糸球体glomerulusと呼ばれる構造で,また後者はボーマン囊と呼ばれる。ボーマン囊は尿細管の続きで,両者の連結部を尿管極という。また,これと反対側には2本の細動脈の出入りする血管極が存在する。この二つの細動脈は小葉間動脈に由来し,血管極から入る輸入細動脈と,血管極から出て尿細管周囲の毛細血管網へと続く輸出細動脈からなる。血管極からボーマン囊へ入った輸入細動脈は吻合をもった複雑な毛細血管網となり,もつれた糸だま状を呈するので糸球体と呼ばれる。この部分では,血管は平滑筋層を失い,動脈と動脈の間に存在する毛細血管という特殊な構造となっている。
糸球体を通る血液は,その血液の圧力により物理的にろ過されて原尿となり,ボーマン囊に受けとめられるが,その間に血液尿関門とよばれる特有な構造が存在する。
糸球体の毛細血管の内皮細胞には径500~1000Åの小孔が無数にあいており,窓あき型の毛細血管を形成している。この内皮細胞の外側には,基底膜が厚い完全な連続層をつくっている。その厚さは成人では200~400nmであり,中央部の緻密(ちみつ)層およびそれをはさむ内外の疎性層から成る。さらに,この外面にはボーマン囊の内葉に由来する被蓋細胞(タコ足細胞)から出る突起の末端終足が接している。タコ足細胞の終足は長さ約1μm,太さ約100nmの細胞質突起で,基底膜に接する底部が平らにひろがっている。互いにかみ合い,その間にすきま(スリット)とよばれる小さい数多くの穴を形成している。このすきまはスリット膜とよばれる薄い膜に架橋され,その上面を粘液多糖類の糖衣がおおう。基底膜はところどころで2層に分かれて,その間にメザンギウムmesangiumと呼ばれる結合組織成分が存在し,そこにメザンギウム細胞が存在する。この細胞は食べ込み能をもつとも,収縮能をもつともいわれるが,その働きは明らかではない(図6)。
上述した血液尿関門では,単純な限外ろ過が行われ,コロイド粒子(径約30Åのヘモグロビン分子より大きなもの)はせき止められ,それより小さな物質は通過する。細かく見ると,窓あき内皮細胞には巨大分子を止める能力はなく,基底膜が限外ろ過の主役を演じる。また,この基底膜を通過したものでも,さらに細かい障壁であるすきま,およびその上にある糖衣によってさえぎられる。以上のようにしてろ過されて原尿が生成される。両側全糸球体の毛細血管の長さの総和は50km,その表面積は1.5m2に及ぶ。糸球体でつくられた原尿の量は,1日に170~200lにも達する。ところが,1日に排出する尿量は2l足らずである。これは,原尿が後にボーマン囊から尿細管に入り,これを通る間に糖やアミノ酸とともに,99%までが再吸収されることに起因する。
尿細管は,長さ10~20cmにも及び,皮質,髄質を往復する特異な走行を示す細い管で,その間に太さや構造および機能が変化する。
ボーマン囊から出た尿細管は,腎小体の回りにとぐろを巻き,近位尿細管曲部を形成する。この部分は主部とも呼ばれ,送られてきた水分の75%を再吸収する。この部分の細胞では,刷子縁(さつしえん)と呼ばれる微絨毛(びじゆうもう)が,ブラシ状に配列した構造が上縁に見られ,また細胞基底部は深いひだをなし,また隣の細胞のひだとも深くかみ合い,基底陥入といわれる構造をなしている。これら二つの構造は,主部における活発な再吸収機能と深く関連し,物質の能動輸送を行う形質膜の面積をいちじるしく増加させている。また,基底部には基底陥入の間をミトコンドリアが縦に並び,能動輸送に必要なエネルギーを供給していると考えられている。主部の細胞には,このほかコロイド状異物のとり込み能があり,細胞内にしばしば大小の水解小体が見られる。
近位尿細管曲部は次にヘンレ係蹄(ヘンレのわな)と呼ばれる部分へと移行する。この部分では,尿細管は一度髄質へ下行し,そこでUターンして上行し,再び皮質にもどる。すなわちループを形成している。このループは近位尿細管直部に始まり,ついで急に径15μm程度の細い管(ヘンレ係蹄の細い部分)となり,髄質内でUターンし(ここまでをヘンレ係蹄の下行脚,これより後を上行脚ともよぶ),上行する。なお,ヘンレ係蹄の細い部分は,この折れかえりの後に再び太くなり(ヘンレ係蹄の太い部分),遠位尿細管直部となる。
下行脚の太い部分(近位尿細管直部)の構造は近位尿細管曲部とよく似ている。またヘンレ係蹄の細い部分は,かなり扁平な細胞からなり,表面には短い微絨毛が散在し,またミトコンドリアも小さいものが細胞内にばらつくに過ぎない。上行脚の太い部分では少数の短い微絨毛が見られ,また,基底部には基底陥入と,これに沿って配列する細長いミトコンドリアが見られ,後述するナトリウムイオンの能動的吸収に関与しているといわれている。
ヘンレ係蹄の上行脚となって再び皮質にもどってきた尿細管は,介在部へと移行するが,この部分は一般に遠位尿細管曲部と呼ばれている。介在部の細胞は,尿細管を通じて最も発達した基底線条(基底陥入とそれに応じて配列するミトコンドリア)を示すが,微絨毛は低くまばらである。介在部は,集合細管を経て集合管に至るが,その間,血管極のごく近くで必ず一度,輸入細動脈に接近し,ここに緻密斑とよばれる特殊な構造を形成する。この部には,扁平につぶれたような核をもつグールマーティ細胞Goormaghtigh's cellが層をなして密集しているが,これは後述する腎臓の内分泌と深く関与している(図7,8)。
糸球体での原尿の産生が,物理学的なろ過であったのに対して,尿細管では生体に必要なものを再吸収し不必要なものを捨てるという選択的な,そして能動的な,いいかえれば生物学的な処理がなされる。
腎小体から送られてくる原尿の水分と電解質の75%は主部で再吸収される。この部での水の吸収にさいしてはナトリウムイオンNa⁺と塩素イオンCl⁻が能動的に吸収され,これに伴って受動的に水が吸収されるというのが特徴である。また生命にたいせつなブドウ糖やアミノ酸などが選択的に再吸収を受け,他方,パラアミノ馬尿酸や少量のクレアチンなどが排出される。
ヘンレ係蹄では,原尿の5%に当たる水分が再吸収される。ここでの特徴は,対向流機構による皮質髄質浸透圧こう配である。これは,ヘンレ係蹄の上行脚においてナトリウムポンプが働きNa⁺の能動的な再吸収が行われるのに対し,水分がこの部であまり吸収されないために起こるものである。この結果,再吸収されたNa⁺が出ていくために尿細管周囲の間質の組織液,およびここを流れる毛細血管の血液は高張となり,他方,ヘンレ係蹄の中を流れる尿はNa⁺のみが再吸収されたために低張となる。このようにして皮質と髄質において濃度こう配ができる。したがって,この濃度こう配の中を下行する下行脚および集合管では,受動的に水が再吸収されることになるのである。
介在部では原尿の15%の水の再吸収が行われるが,この部での水の再吸収は,脳下垂体後葉の抗利尿ホルモン(ADH)によって促進される。これは,視床下部の視索上核および室旁核(しつぼうかく)にある浸透圧受容器による体液調節と深く関係している。すなわち,体液の水分量が増加し浸透圧が低下すると,バソプレシンの分泌が減少し,尿量が上昇するのである。なお,この部分での水の再吸収は電解質の再吸収を伴わないため,尿は高張となる。
以上のようにして生じた尿は,集合細管を経て集合管に合流し,ここで原尿の4%の水分が吸収された後さらに合流をくり返して乳頭管となり,腎乳頭の先端で小腎杯に開口する。乳頭管の数は各腎乳頭当り20~30個である。
腎臓のたいせつな機能としては,老廃物の処理のほか体液の調節がある。体液の水分量や電解質の調整を行っているのである。これらのことは,総括的にみれば,体内の不要物の処理ともいえよう。これらの調節には,種々のホルモンが関与して複雑な系をなしている。たとえば,Na⁺は体液中に多くの割合を占める重要な電解質であるが,これは副腎皮質から出るアルドステロン(鉱質コルチコイド)によって調節されている。このホルモンは,遠位尿細管でのNa⁺の再吸収を促進する。なおアルドステロンは,副腎皮質に対する種々の刺激によって出るほか,腎臓の糸球体旁装置から分泌されるレニンを介して分泌が調節されている。
カリウムイオンK⁺は,Na⁺とは裏腹に行動する。すなわち,K⁺の血漿中の濃度が増加すると,アルドステロンの分泌が増加してNa⁺の再吸収が起こり,逆にK⁺は尿中に排出される。また,K⁺の血漿濃度の増加に伴い,遠位尿細管と集合管におけるK⁺の分泌増進(K⁺の尿細管壁への直接作用による)が起こるといわれている。
そのほか,腎臓の重要な機能として,血液の酸塩基平衡の調節があげられる。これは,血中の炭酸水素イオンHCO3⁻の濃度に応じて炭酸水素ナトリウムとして水素イオンH⁺が排出または再吸収されることや,H⁺がリン酸やアンモニアと結合して排出されることを介して調節されている。
輸入細動脈が糸球体の血管極にはいる直前の部分では,中膜の平滑筋細胞がやや球状ないし立方状の細胞になっていることが多い。この細胞の細胞質は,ヘマトキシリン-エオジン染色ではただ明るく見えるだけであるが,ゲンチアナ紫,エチル紫,中性赤などによって分泌顆粒(かりゆう)様の構造を染め出すことができる。この上皮様細胞は,動脈の壁からはなれて結合組織の中に小島を作っていることもある。これらの上皮様細胞をまとめて糸球体旁細胞juxtaglomerular cellとよぶ。そして緻密斑,グールマーティ細胞,糸球体旁細胞をまとめて糸球体旁装置juxtaglomerular apparatusと称する。
糸球体旁細胞は,血管中膜の平滑筋細胞の特殊化したものである。電子顕微鏡で見る糸球体旁細胞は,細胞質中にオスミウム好性の大型の分泌顆粒をもち,その中にレニンを含む。レニンは血漿中のアンギオテンシノゲンにはたらき,アンギオテンシンという血圧上昇因子をつくるのに必要な一種のタンパク質分解酵素であることが知られている。
→尿
腎臓がもつ尿排出,体液の調節,内分泌機能がなんらかの原因でおかされると,種々の症状を呈し,次のような病気が発生する。
まず症状としては,糸球体でのろ過と尿細管での再吸収とのバランスが崩れると,多尿になったり,逆に乏尿になる。これは腎臓を支配する内分泌系の異常(抗利尿ホルモンの分泌減少など)や腎臓が関与したレニン-アンギオテンシン系の異常から起こる。また糸球体のろ過の異常によってタンパク尿が,炎症によって血尿や膿尿が起こる。体液の調整機能の異常によって浮腫(水腫)や高血圧(腎臓が原因の高血圧を腎性高血圧という)などが起こる。
腎臓の病気に際しては,以上のような諸症状が組み合わされて表れるが,おもな病気としては腎炎,ネフローゼが最も重要である。このほか,感染による腎盂腎炎,腎結核などや,悪性腫瘍(腎癌),結石(腎結石を含め尿路の結石を尿路結石と総称する)などによって,尿が停滞して起こる水腎症などがある。
執筆者:藤田 尚男
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
Sponserd by ![]()
…そのほか胎盤からは種々の下垂体ホルモンや性ホルモンが分泌される。(12)腎臓 旁糸球体細胞はレニンという酵素を分泌する。これが血中のアンギオテンシノゲンに作用して,アンギオテンシンIを生ずる。…
…この過程が排出である。脊椎動物には排出器官として発達した腎臓があるが,無脊椎動物にも系統群によってそれぞれ特有の排出器官があり,排出器官では,原理的にはろ過,再吸収,分泌の三つの過程を経て尿が作られる。脊椎動物の腎臓を例にとると,まず第1段階として腎小体(マルピーギ小体)で,小動脈よりなる糸球体からそれを包むボーマン囊へ体液がろ過されるが,これは小動脈内血圧と外側の圧の差によって起こる限外ろ過であって,血球と大部分のタンパク質を除く血液成分がこし出される。…
…玉(ぎょく)玉(たま)【町田 章】
[語源,習俗]
ヒスイをあらわす西欧語jadeはもとejadeと称したが,語頭のeが冠詞の一部と誤解されて脱落した。ejadeはスペイン語のpiedra de ijada(〈横腹の石〉の意)から由来しており,この石,つまりヒスイは腎臓の痛みを治癒させるといわれていた。石とは関係のないijada(横腹)からjadeの語は生じたわけである。…
※「腎臓」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
《〈和〉doctor+helicopterから》救急専用の医療機器を搭載し、医師・看護師が乗り込んで患者のもとに急行し、病院などに搬送する間に救命医療を施すことのできる救急ヘリコプター。...