デジタル大辞泉 「恐竜」の意味・読み・例文・類語
きょう‐りゅう【恐竜】
Sponserd by ![]()
Sponserd by ![]()
中生代の三畳紀後期から白亜紀まで(約2億2800万年~6550万年前)の地球上に栄えた陸生の絶滅爬虫(はちゅう)類。これまでに発掘された多くの化石によってその存在が判明している。恐竜は直立する爬虫類で、大腿骨(だいたいこつ)と関節する骨盤の深いくぼみ(寛骨臼(かんこつきゅう))の中央が穴で貫通している。仙椎(せんつい)が3個以上あって、前足の第4、第5指の指骨が減少するなどの共通性がある。
[小畠郁生]
「恐竜」という名称はイギリスのリチャード・オーエンが、1842年にギリシア語の「恐ろしい」deinosと「トカゲ」saurosの二つのことばを合成してつくった。最初は分類学上、恐竜目として提案されたが、1887年にイギリスのハリー・シーリーが、恐竜目として一括されるものではなく、これを竜盤目と鳥盤目という別個の二つの目を設けて分類することとした。1914年にドイツのフリードリヒ・フォン・ヒューネFriedrich von Huene(1875―1969)がシーリーの考えを再検討し強力に支持し、骨盤の形はそれぞれ別の祖先から受け継いだものと考える恐竜二元説はそれ以後長く受け入れられた。ところがアメリカのロバート・バッカーRobert T. Bakker(1945― )とピーター・ガルトンPeter M. Galtonが恐竜は単一祖先由来の自然の系統であるとする単系統説(1974)を発表し、1984年ごろから一般に受け入れられるようになった。分岐分類法で10以上の特徴で恐竜の定義がなされ、祖先と目された槽歯(そうし)類という分類群が消滅したからである。一方、恐竜ということばは、便利で日常的な用語としても活用されている。
竜盤目と鳥盤目という名称は、骨盤の形がそれぞれ「爬虫類の腰」と「鳥類の腰」という意味に由来する。竜盤目は、原則的には骨盤の下部の骨(坐骨(ざこつ))が上部の骨(恥骨(ちこつ))に対してある角度をなしている。すなわち竜盤類は爬虫類様式に従って配列された三つの腰の骨をもつ。ただし、ドロマエオサウルス類やテリジノサウルス類のような例外がある。また、このタイプは祖先の主竜類(恐竜、翼竜、ワニ、ラゴスクス類などを含む上位の大きなグループ)にすでに出現しているので竜盤類の定義にはほかにいくつかの特徴が使われる。一方、鳥盤類では腰の骨の配列が鳥の骨盤に似ていた。骨盤の上部の骨(恥骨)が長くて、下部の骨(坐骨)に対し並行的であった(参照)。
恐竜のなかには、きわめて大形のものもあれば、非常に小さいものもあった。重量が80トンに及ぶ大きい種類のものから、頭から尾までの長さがわずか60センチメートルという小さな種類までいた。
[小畠郁生]
竜盤目は獣脚類(獣脚亜目)と竜脚形類(竜脚形亜目)とに分けられる。獣脚類は二肢歩行性、肉食性で、テタヌラ類(テタヌラ下目)Tetanuraeとケラトサウルス類(ケラトサウルス下目)Ceratosauriaに分類され、ヘレラサウルス科Herrerasauridae、エオラプトルEoraptorを含む。テタヌラ類はさらにスピノサウルス上科Spinosauroidea、鳥獣脚類Avetheropodaに分けられ、後者はコエルロサウルス類Coelurosauria(軽い骨組の系列)とカルノサウルス類Carnosauria(大形化する系列)で構成されている。竜脚形類のほうは、草食性で大形化するグループで、古竜脚類(古竜脚下目。竜脚類と平行関係にあたるもので三畳紀に出現し、比較的小形で雑食性、二肢あるいは四肢歩行)と竜脚類(竜脚下目。ジュラ紀から白亜紀にかけ繁栄し、大形重量級で草食性、四肢歩行)からなる。なお、恐竜時代には被子植物の草は出現していなかったので、「草食」ということばを避けて一般に「植物食」という用語にする学者もいる。
次に鳥盤目は鳥脚(ちょうきゃく)類(鳥脚亜目)、装盾(そうじゅん)類(装盾亜目)、周飾頭(しゅうしょくとう)類(周飾頭亜目)に分けられる。そのうち、装盾類は剣竜(けんりゅう)類(剣竜下目)とよろい竜類(よろい竜下目。曲竜(きょくりゅう)類、装甲竜類ともいう)を含み、周飾頭類は角竜(つのりゅう)類(角竜下目)と厚頭竜類(厚頭竜下目)、さらにレソトサウルスLesothosaurusやピサノサウルスPisanosaurusを含む。すべて草食性であって、鳥脚類だけが二肢歩行性で、ほかは四肢歩行性であった。剣竜類は体の表面に顕著な突出物をもち、よろい竜類は背側全般にわたり固く武装していた。角竜類は普通、顔の前面にサイに似た角をもっていた。
近年では、属名と種名以外のリンネの分類名を使うことを避ける傾向がある。それは階級を示す用語(目、亜目、下目など)の定義が不明確のゆえである。しかしここでは混乱を避ける意味で、ある程度は従来の伝統的手法との対応を示すよう併記した。
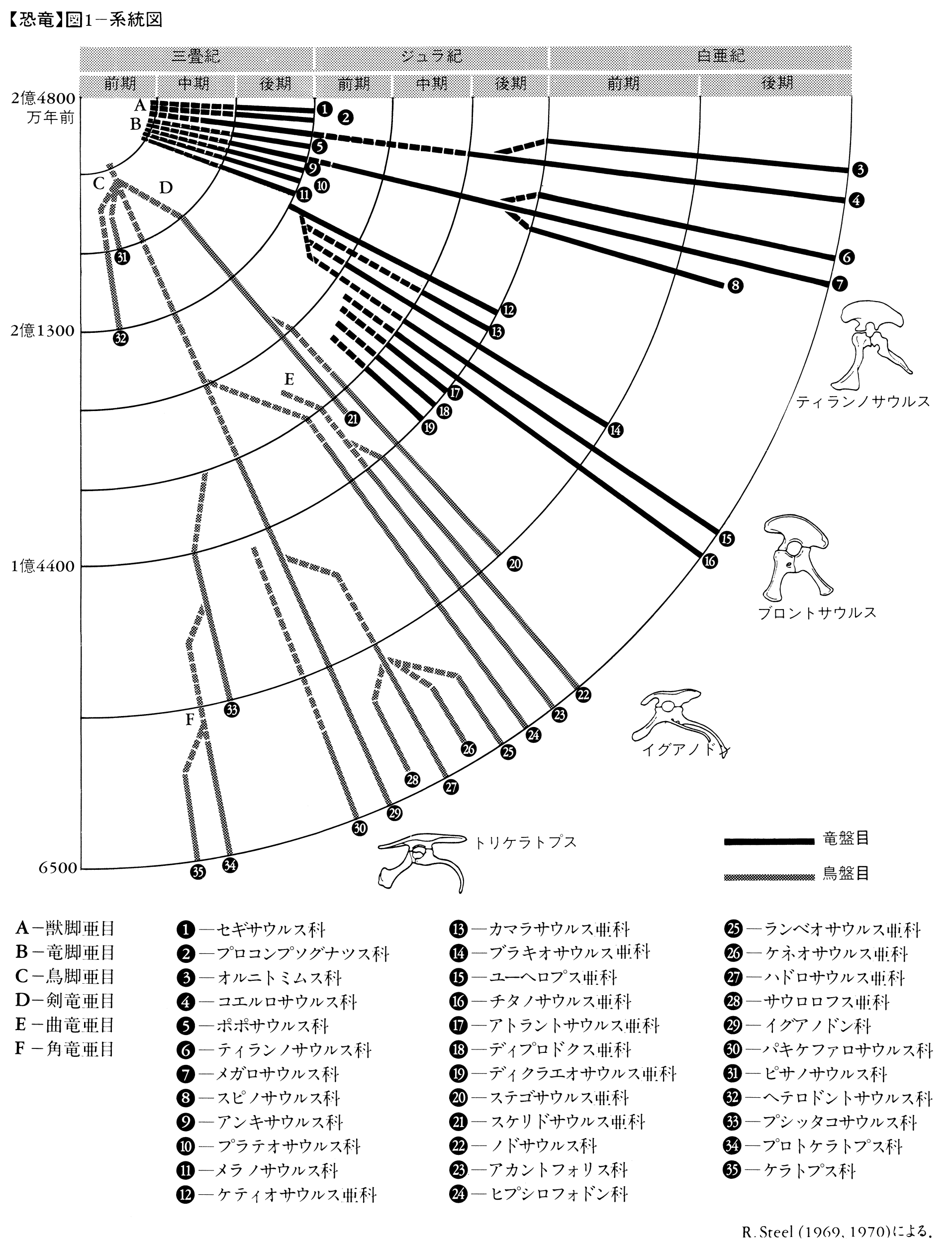
中生代を通して適応放散を繰り返した恐竜類であるが、1994年までに学界で有効な種類と認められたのは約350種である。しかしその後も年に7~8属以上の増加ぶりなので、2005年現在では625種類以上が知られている(原始的鳥類73種を含む)。なお、恐竜の系統樹を、その詳細図を、、に示す。
[小畠郁生]
かつては、恐竜は現生爬虫類との類似性から冷血(変温あるいは外温性)の絶滅爬虫類であると考えられていた。ところが、20世紀の後半に入ってから、恐竜学者のなかに、恐竜は温血動物であったのではないかという主張が現れてきた。その論点は、(1)恐竜の一般的な体形、(2)恐竜の骨の微細構造、(3)肉食竜と草食竜の個体数比、の3点からなっている。議論の数例は次のとおりである。
カナダのジョン・ハロルド・オストロームJohn Harold Ostrom(1928―2005)は、体高が高くエネルギー生産力の高い動物だけが肢(あし)をぴんと伸ばして直立した姿勢をとっているのは偶然ではなく、そうでなければ、二本肢でまっすぐ立って歩くことはできないと考えた。たとえば、ティラノサウルスTyrannosaurusのような姿勢でバランスをとって歩くことができるということから、恐竜は冷血で動きの鈍い爬虫類という考えを否定すべきであるとした。これに対し、爬虫類でも短時間の急激な運動は可能である点や、巨大恐竜が恒温であった場合には代謝が活発で過熱に悩まされたのではないかなどの反論がある。1973年にカナダの北極圏の白亜紀層からカモノハシ竜のハドロサウルスHadrosaurusが発見されたが、白亜紀当時の極地は現在よりも暖かかったので、かならずしも温血説の証拠とはならないとされた。
パリ大学のアルマン・ド・リクレArmand de Ricqlès(1938― )は種々の化石骨と現生動物の骨の内部構造を調べ、1969年に、恐竜は生理的にはトカゲよりも哺乳(ほにゅう)類に近いと報告した。恐竜の骨では、哺乳類の骨のように多数のハバース管(骨の内部にある細い管で、中に血管が通っている)がぎっしりと詰まっているのに、現生爬虫類ではまばらである。これは、活発に動く動物は生理的なつくりが優れていて、それが骨の細かい構造にも現れているからで、恐竜ではエネルギーが活発につくられ温血であったからであろうという論旨である。これへの反論としては、ある種のカメには多数のハバース管があるのに、小形哺乳類や鳥類のなかにはほとんど発達しないものもあるという例があげられていた。
1972年にバッカーの論じたところでは、コモドオオトカゲは体重45キログラムで、月に1回、自分の体重の半分のブタ1頭分を食べるが、これは60日で自分の体重分の餌(えさ)をとるという計算になる。一方、哺乳類のチーターは10日で自分の体重分の餌を食べ、ライオンは8日、野生のイヌは7日以内で食べる。つまり恒温動物では餌の必要量はトカゲの約10倍である。恒温動物は「餌動物」を頻繁に殺さないと生きていけず、一定数の草食動物を餌として生活できる動物数は少ない。コモドオオトカゲのような現生爬虫類では、一定数の草食動物に対し比較的多数の肉食動物が生きることができる。
バッカーの研究によると、古生代ペルム紀(二畳紀)の古代爬虫類が栄えた約2億9900万~2億5100万年前は、新生代の古代哺乳類が栄えた6550万~181万年前に比べ、肉食動物の数の割合がはるかに高いが、恐竜の世界ではこの比が哺乳類の場合に近く、肉食動物の比がずっと低い。今日のアフリカ大陸でカモシカの数がライオンの数よりも断然多いのと同様に、中生代白亜紀末にはカモノハシ竜が獣脚類よりも圧倒的に多かったらしい。これらを総合すると、恐竜は冷血動物というよりも温血動物であると考えるべきではないかというのであるが、異論を表明する者も多い。
1990年代の後半以降、羽毛を生やした小形獣脚類が続々と報告されるようになった。中国遼寧(りょうねい)省の白亜紀前期の地層から産出する化石生物群(熱河(ねっか)生物群)中に、2骨格の頭頂部から尾端までと尾の下側に羽毛様構造をもつ全長1メートルの獣脚類が発見され、シノサウロプテリクスSinosauropteryxと命名された(1998)。ついで、真の羽毛をもつ獣脚類2種が発見され、プロトアーケオプテリクスProtarchaeopteryxとカウディプテリクスCaudipteryxと命名された(1998)が、ともに羽毛が羽軸(うじく)を中心に対称的であって活発な飛行はできず、地上を走ったと思われる。羽毛をもつ最大のベイピャオサウルスBeipiaosaurus(1999年発見、以下同じ)の前肢は綿毛のような繊維で覆われていた。シノルニトサウルスSinornithosaurus(1999)は綿毛のような繊維をまとい、鳥型の肩帯(けんたい)(脊椎(せきつい)動物の前肢の肢帯)をもつ。前肢を上下に動かし、鳥のように羽ばたいた。走行性から飛行性への途上にあった。ミクロラプトルMicroraptor(2001、2002、2003)は全身をダウン状羽毛が包み、前後肢には羽軸と羽枝(うし)を伴うが、これは樹上からムササビのように滑空したらしい。シノベナトルSinovenator(2002)も羽ばたくように腕を自由に動かせた。これらは小形獣脚類から鳥類へと進化する段階を示唆し、彼らが温血性であったことを物語るらしい。
なお、鳥脚類恐竜のテスケロサウルスThescelosaurusの心臓があるべき場所に心臓が化石になったとされる岩塊が発見されて、賛否両論が出されたことがある。その後、病院で行った三次元CTスキャンの結果、鳥類やワニ類同様に2心房2心室であった。ただし、1本の大動脈を備え、左右の心室は完全に分かれており、ワニ類よりは鳥類や哺乳類の場合により近く、非常に高い物質代謝率が示唆されている。
[小畠郁生]
普通、動物園などで現生爬虫類は鳴き声をたてないようにみえるが、伊豆(いず)(静岡県)の熱川(あたがわ)のワニ園を訪れると、ワニは興奮したときや驚いたときに声をたてる。怒ったときにも短く甲高いほえ声をあげる。また雄が雌をよび、他の雄を追い払うときはしわがれた声を出すという。恐竜の耳のつくりはワニ類に似ているので、聴覚は発達していたらしく、よく聞こえる耳をもつので、彼ら自身もある種の声を出すことができたと思われる。多くの種類で舌骨(ぜっこつ)(舌顎骨(ぜつがくこつ))が知られる。舌骨は声帯の発達と関係が深い。たとえば白亜紀のトリケラトプスには大きい舌骨があるので、大きい声を出すことができたのであろう。肉食のタルボサウルスTarbosaurusやティラノサウルスでは餌動物を脅かすような、のどから張り上げる大きな声を想像することができる。
次に嗅覚(きゅうかく)の推察であるが、現生の哺乳類、爬虫類のあるものや鳥類を除いては、両生類以上の脊椎動物では、鼻腔(びくう)の一部が左右に膨らんで出ていて1対の袋状の器官となっており、これはヤコブソン器官とよばれる。その内面は嗅覚上皮に包まれていて、嗅神経の一部が分布している。このヤコブソン器官はヘビ類、トカゲ類ではにおいのおもな受容器として働き、引っ込めた舌の先端が正しくその開口部にあてがわれ、捕捉(ほそく)したにおいのする物質を嗅覚上皮に送り込むといわれる。この器官はワニ類にはなく、カメ類でも退化しており、恐竜類では不明とされていた。
恐竜の場合、すべて鼻孔が大きいが、ときには非常に大きいものがある。パラサウロロフスやコリトサウルスのようなカモノハシ竜には鼻道がよく発達するが、その長い理由の一つに、大きな鼻孔の奥にまで血管がよく通っており、泡のような海綿状構造があって空気をここで温めたという解釈がある。嗅覚細胞は鼻道の内側の上皮細胞にある。ほとんどの爬虫類では嗅覚上皮細胞に覆われる表面が比較的小さいため、もしカモノハシ竜などが嗅覚に頼っているとしたら、それを発達させる唯一の方法は鼻の通路自体を長くすることであった。哺乳類の上皮細胞は、あるところでは鼻腔の広がりにより、またあるところではその腔の中に薄い上皮細胞をかぶった骨、甲介骨があることで拡張されているが、カモノハシ竜には甲介骨がないので鼻道を長くするしかなかったのであろう。いずれにせよ嗅覚は、脳型の知られているすべての恐竜でよく発達していたようである。嗅覚はむろん餌や敵ならびに仲間のにおいを認めるために重要であったと思われる。恐竜はにおいに敏感であったに違いない。
恐竜の目はよく発達しており、なかには非常に大きいものがあった。多くの種類で目は強膜板の輪でじょうぶになっていた。カモノハシ竜では、強膜と角膜の間の重なり合った板からなる小さな輪の数は14個あった。これは目の保護のためか、動いて目を圧縮し焦点をあわせて物を見るためのものかもしれない。恐竜の目はよく見えたのである。その一部は、距離の遠近までよくわかる両眼視であったと思われる。多くの爬虫類は赤みか、あるいは黄みがかった目をもつ。恐竜の目の色がどうであったか、自分の保護色と同じであったかどうかは不明である。ウミガメやトカゲ、ワニ類をはじめ、現生の爬虫類や鳥類では色視細胞が哺乳類よりも発達している。彼らが色彩を見分けることに優れているのであれば、恐竜類でもその可能性が高い。さらに、恐竜類の化石は生存時には熱帯ないし亜熱帯域であったと考えられる場所から多く知られている。恐竜類の色視細胞が優れていたのならば、彼ら自身の体色がカラフルであって、彼らの体色をお互いに識別できたのではあるまいか。
[小畠郁生]
恐竜が考えていたことを想像するのはむずかしい。脳の物質そのものは残っていないが、その組織が収まっていた脳腔を調べ、その床にある窩(あな)すなわちくぼみを調べることができる。脳神経のための孔は下等な脊椎動物では位置がかなり安定していたので、孔を計測することや脳の長さを知ることができる。視葉の膨らみを調べ、耳の部分を復原し、さらに嗅部を明確にすることができる。実際の器官としての脳についてはなにもわからないが、多くの種類で脳腔の雄型の形すなわち頭蓋骨(とうがいこつ)により囲まれた空間の頭蓋内腔が研究されてきた。現生のムカシトカゲSphenodonやワニ類の脳は脳腔から想像されるよりもはるかに小さいので、恐竜の脳の考察を行う際に注意しなければならない。恐竜の頭蓋内腔雄型が知られているのは以下のものである。すなわち、獣脚類のケラトサウルス、トロオドン、アロサウルス、ティラノサウルス、古竜脚類のプラテオサウルス、竜脚類のブラキオサウルス、ディプロドクス、バロサウルスBarosaurus、カマラサウルス、鳥脚類のカンプトサウルス、イグアノドン、アナトサウルスAnatosaurus、厚頭竜類のパキケファロサウルス、剣竜類のステゴサウルス、ケントロサウルス、よろい竜類のアンキロサウルス、エウオプロケファルスEuoplocephalus、角竜類のプロトケラトプス、アンキケラトプスAnchiceratops、トリケラトプスなどである。
恐竜の脳は、身体の大きさと比べると比較的小さく、ときには異常なほど小さい。脳の構造はワニよりすこしだけ進んでいる。鳥類や哺乳類の丸い緻密(ちみつ)な脳と違い、脳の形は一般に高さが低く細長い。ワニ類のように、表面が滑らかで巻き込んでいるようなことはなかったと想像される。ティラノサウルス、プラテオサウルス、パキケファロサウルス、アンキケラトプスには長い嗅索がみられ、大脳半球の膨らみがあった。脳の大きなティラノサウルスやパキケファロサウルスでは脳下垂体が大きかった。脳の構造から、多くの恐竜はほとんど考えることができなかったらしいと思われている。脳の嗅覚は、脳型の知られるすべての恐竜でよく発達しており、鼻孔はすべて大きい。最大のティラノサウルス個体(通称「スー」)の脳のCTスキャンでは、においを感知する知覚神経にあたる嗅球が脳の半分もの容積を占めていた。
現代のゾウほどの重さの剣竜類ステゴサウルスの脳は小さな子ネコの脳よりも小さかった。巨大恐竜の脳は多くの場合、脊髄(せきずい)の直径よりも小さく、肢や尾の運動を支配する肩や腰での脊髄拡張部(神経叢(そう))よりも小さかった。たとえばステゴサウルスの場合、脳の大きさは腰の神経叢の大きさの20分の1ほどしかなかった。恐竜の成長ホルモンを分泌する脳下垂体が大きかったことは、恐竜の体の巨大化と関係が深かったかもしれない。いずれにしろ、恐竜の脳は低級であるとするのが定説であったが、20世紀後半には、ずっと大きな脳をもつ恐竜が報告されるに至っている()。
カナダで1914年に発見され1969年に発表された白亜紀のドロマエオサウルスという恐竜は、頭骨に脳が入る頭蓋腔をつくっていた部分がだいたい残っていたが、この頭蓋腔の著しい特徴は形と大きさで、ほかの肉食恐竜にはまったくみられないほど大きなものであった。これはその祖先のデイノニクスDeinonychusと同様に凶暴であったと思われる。1968年にカナダのアルバータ州でみつかった小形肉食恐竜のトロオドンは、頭蓋をみると眼窩(がんか)が異常に大きく、小さな恐竜が直径5センチメートルという大きな目をもっていた。トロオドンの頭蓋腔の容積は49立方センチメートルで、生存時は約45グラムの脳が詰まっていたとみられるので、体重45キログラムとして脳重量は体重の1000分の1以上であり、ほぼ同大の鳥エミューの脳が1227分の1の大きさであったことをしのぐ。この恐竜では、高度の行動を統御したり複雑な運動の統合調整を行ったりしたため、大脳半球が大きかった。1922年にモンゴルで発見されたサウロルニトイデスSaurornithoidesもこれと同様で、高い知能をもっていた。
[小畠郁生]
恐竜の性別ないしは性的二型性については、オテニオ・アーベルがカモノハシ竜ラムベオサウルス類Lambeosauridsのとさかの差異が性によるものではないかと1924年に提案した例のように、鳥脚類や角竜類について多く言及されてきた。こういった議論は1990年代以降活発となってきている。
マイケル・ラートMichael A. Raathはアフリカで小形獣脚類シンタルススSyntarsusのボーンベッド(骨化石包含層)を調べて、成熟した骨格には「がっしり型」と「ほっそり型」の2型があり、前者の個体数が多いので、雌であると結論している(1990)。
ケネス・カーペンターKenneth Carpenter(1949― )はティラノサウルスなどを調べ、「がっしり型」と「ほっそり型」を認め、前者の坐骨の形は卵が通過するように大きな空間を示すので、雌であるとした(1990)。ラルフ・モルナーRalph E. Molnarもティラノサウルスの頭骨の研究から性的二型性に言及し、後眼窩(がんか)骨上のうねの有無で性差の可能性を述べた(1991)。
ピーター・ラーソンPeter L. Larsonはティラノサウルスの15標本を精査し、頭骨・上腕骨・坐骨・仙骨などで「がっしり型」と「ほっそり型」を認め、前者の幅広い仙骨と角度の急な坐骨は卵の通過のためにより広い空間をなすと考え、雌の特徴とした。さらに現生の肉食鳥類の猛禽(もうきん)類でも雌が雄よりも大きいことや、現生のワニ類では尾椎(びつい)の基部についている骨の棘(とげ)を意味する第1血道弓(けつどうきゅう)と第2血道弓が、雄では同大なのに雌では前者が後者の半分のサイズしかないことなどに注目した(1993)。雄ではペニス収縮筋が第1血道弓に固定され、ペニス不必要時には体内に引っ込ませる役をしている。ティラノサウルスの「ほっそり型」でも同様と考えられた。他の獣脚類サウロルニトイデスでも「ほっそり型」では第1血道弓が第1尾椎の前にあり、第2血道弓と同形同大である一方、「がっしり型」では前者が第1尾椎の後ろにあり、後者より短く、くさび型である。「がっしり型」が雌で「ほっそり型」が雄というのは、雄間の競争が不必要な一夫一婦制の表れである可能性があり、カモノハシ竜などでは雌・雄で2型が逆の関係らしいと予想されるものがある。ティラノサウルスの「スー」が発見されたとき、第2尾椎と第3尾椎の間に血道弓がみつからなかったので、「スー」は雌であるという仮説が生まれていた。ところが、その後、血道弓が発見されたが、それで「スー」が雄であるとはいえないといわれる。ティラノサウルスの性差がワニと同じ場所に同じように現れるという保証がないからであるという。
[小畠郁生]
1976年イギリスのロバート・マックネイル・アレクサンダーRobert McNeill Alexander(1934―2016)が発表したところでは、恐竜の歩行の際の速さは秒速1~3.6メートル、1時間に換算すると約3.6~13キロメートルとされている。ほぼ人間なみである。これは、骨格化石の示す恐竜の大きさと、地層に残された足跡から考えられる足裏の長さや歩幅から計算された。現存動物と人の、歩いている場合や走っている場合の速さと歩幅、地面から腰までの高さを測った結果、動物の速さと歩幅、腰の高さの間に一定の関係があることがわかったので、恐竜の場合に当てはめてみると、竜脚類は時速3.6~4キロメートルで群れをなしてのろのろ歩き、二本肢恐竜は時速7~8キロメートルで、速いものは時速10~13キロメートル、遅いものは4.3キロメートル程度であった。2002年にイギリスのジュラ紀層の足跡化石について発表された例によると、肉食恐竜の歩行跡145メートルのあとに走行跡35メートルが続いていた。この足跡の解析結果は、時速7キロメートルで歩いていたのが、時速30キロメートルに転じたらしい。
一方、恐竜はかなりの速さで走ったとする説がある。バッカーは肢の関節を調べ、角竜類は最初の攻撃で敵を追い払うことができないと、サイのように時速50キロメートルもの速さで逃げたのではないかという説をたてた。ダチョウと形や習性が似ていたと思われる獣脚類のダチョウ恐竜オルニトミムスOrnithomimusは、ダチョウなみに時速80キロメートルを出せたのではないかと考えられているが、この恐竜のエネルギー生産力が爬虫類なみの生理機能であるとすると、時速わずかに3キロメートルを維持する程度という。肉食恐竜のデイノニクスは二本肢で、後ろ足には地面につく指が2本しかなく、もう1本は12センチメートル余りの鉤(かぎ)づめをもつ武器で、このつめで獲物を引き裂くとき、一本肢で立っていなければならなかった。優れた平衡感覚をもち知能が高く足も速そうなのに、同体重の普通の爬虫類なみとすると、最適な気温時においてさえも時速1.5キロメートルの速度しか維持できず、また、ティラノサウルスやアナトサウルスがもし冷血動物ならば時速5~6キロメートルがやっとであったということになる。恐竜の姿がたいへんな速度を出せるような構造を示しているにもかかわらず、生理的にゆっくりと断続的にしか動けないというのも不自然なことである。
これら恐竜の歩行の速さの判定は、歩幅の大きさと歩調の速さによる。脛骨(けいこつ)と大腿骨の長さの比をみると、ほぼどのくらいのスピードを出せる動物であるかがわかる。大きなアパトサウルスApatosaurusでは脛骨と大腿骨の比は現代のゾウとほぼ等しく、0.60を示す。太い肢や関節のつくりがゾウによく似て、重量を支える目的に適し、膝(しつ)関節はあまり曲がらない。竜脚類の歩く速さは遅かった。カナダのロリス・ラッセルLoris S. Russell(1904―1998)によると、ダチョウ恐竜の骨と肢の筋肉はダチョウに比べると優れており、少なくともドロミケイオミムスDromiceiomimusという恐竜にはダチョウよりも速く走れるのがいたという(1966)。ヒプシロフォドンの脛骨と大腿骨の比は1.18に達しており、きわめて速く走ることができたらしい。脛骨と大腿骨の比だけが速度判定の基準ではないが、いちおうの目安となる。恐竜の恒温動物説に基づくと、速い速度をもつ恐竜がいても不思議ではない。
実際には、恐竜の歩行や走行の速度は、普通は個体の連続した足跡の計測値を基に経験式から求める方法を使うが、これに対して、骨格や筋肉、復原像の検討から生体力学的に求める方法も考えられている。この方法によりティラノサウルスの肢の骨格と筋肉が検討されたが、2002年に発表されたその結果によると、普段は時速18キロメートル以下で歩き、時速40キロメートルが限界であったとされている。近年では、親は時速15キロメートル程度で、子は肢が長く、ラプトル(ベロキラプトルVelociraptor、ユタラプトルUtahraptorなど)級の速さを示し、親子共同で狩りをしたという説が出されている。
[小畠郁生]
恐竜の群れが化石として発見された例は、1878年にベルギーのベルニサール炭鉱の地下約300メートルの坑道から発見された草食のイグアノドンの群れである。夾炭(きょうたん)層にできていた深い裂け目が落し穴となって、イグアノドンは次々と落ち込んだものらしく、発掘されたものを総計すると、少なくとも成熟したイグアノドン29頭分もあった。同一種の恐竜がまとまって1か所から産出したという顕著な例である。不思議なことに、このときの化石群には子供の化石は含まれていなかった。
化石とくに足跡の証拠に基づくと、恐竜にはゾウのように群れをなして行動するものが多かったらしい。恐竜は普通考えられているよりも、はるかに高度な群れとしての共同生活を営んでいたものと思われる。アメリカのローランド・サクスター・バードRoland Thaxter Bird(1899―1978)がテキサス州で発見した恐竜の足跡は、合計23頭のアパトサウルスが、おそらく同一目標に向かって並行して歩いていった足跡であろうと判定されている(1944)。おもしろいことに、最初に単独で1~3頭が歩いて、後ろの群れを導いたようである。また、15頭以上のアパトサウルスの群れが歩いたと思われる跡も発見されており、ほかにも、大・中・小の3種類の足跡の群れ134個がアメリカのマサチューセッツ州でみつかっていて、これについてはオストロームが「このうち28個は歩行中の恐竜のもので、20個はみな西の方向に向かっていた。それらは一団となって同一方向に向かって歩行していたものと考えざるをえない」と述べている。このように社会性をもつ恐竜もいたらしい。
アメリカのモンタナ州北西部では、ジョン・ロバート・ホーナーJohn Robert Horner(1946― )らにより、カモノハシ竜マイアサウラMaiasauraの営巣地の跡と、子育てをしたらしい証拠が数多く発見された(1979)。さらにマイアサウラのボーンベッドが広大に存在するので、1万頭に達するマイアサウラの群れが季節ごとに南北に大移動したと推定されている。カナダのアルバータ州でも、フィリップ・ジョン・カリーPhilip John Currie(1949― )らにより、角竜類セントロサウルスCentrosaurusの数百頭の群れが洪水時の川を渡ろうとして溺死(できし)したと思われる産状が発見された(1984)。カリーは、1910年にバーナム・ブラウンBarnum Brown(1873―1963)がレッド・ディア川沿いで9体のアルバートサウルスAlbertosaurusの化石を発掘したのを引き継ぐ形で98年に同地点付近の調査を開始し、2006年には22体に達している。これはアルバートサウルスが群れをなしていたことを強く示唆しており、成体と幼体の集団であった。大形肉食恐竜の集団はアルゼンチンでも発見されている(2006)。
群れの組織と運営がどのようであったかは今後の研究課題であろう。イギリスのエードリアン・デズモンドAdrian J. Desmond(1947― )によると、竜脚類は群れとして移動しながら、胎内で孵化(ふか)した子を産み落としたが、その子はすぐに群れに入って保護されたのではないかと述べている(1975)。また一方バッカーは、群れの周りを成体が囲み、子供は群れの中で守られていたのではないかと論じた。しかしその後、アメリカのマーチン・ロックレイMartin G. Lockley(1950― )は、バードが表した竜脚類の足跡の地図を基に、足跡の踏んだ、踏まれたの関係を研究し、どの個体が先頭を歩き、どれが最後に歩いたのか、どのくらいの速さで歩いたかなどを調べて群れの構造を解析した。その結論では大小の個体はランダムに配置されていた(1987)。また、松川正樹(まさき)(1950― )が足跡のサイズ頻度分布と相対成長解析を行うと、23個体の竜脚類は2世代の集団で存在していたと判定され、大小の足跡の配置からは、大人が子供を守るようにして歩いた群れではないとする解釈が裏付けられた(1998)。
コロラド州南部のパーガトリー川のジュラ紀後期のモリソン層の竜脚類足跡は、西に向かう五つの歩行跡が適度な間隔を保っており、やがて五つの歩行跡はすべて左にカーブしていることから、5頭が同時に並んで歩いていたものと思われる(ロックレイ、1987)。松川の解析では、1世代の一つの成長段階の集団からなるが、5頭の歩行速度を計算すると、速く歩く個体と遅く歩く個体があり、最大で毎秒2.5メートルほどの差があったとされている(1998)。なお、上記と同様なカーブの歩行跡は、カナダのブリティッシュ・コロンビア州にある白亜紀前期の鳥脚類の群れと思われる足跡化石でも知られている。
富山県大山町(現、富山市)の白亜紀の手取(てとり)層群から発見された足跡を研究した松川らは、足跡はトヤマサウリプスToyamasauripusで、33頭いたことを確認した。さらに、足跡は3方向へ群れで移動し、たとえば、北西へ進む4頭は時速1.2~1.5キロメートルで歩き、歩行間隔はほぼ1メートルで平行していたことを発表した(1997)。このように、連続した足跡化石の研究などから、恐竜たちの群れの構造は徐々に明らかにされつつある。
[小畠郁生]
恐竜が草食性であったか肉食性であったかは、それらの化石の歯の形とあごの仕組みをみるとわかる()。竜盤類のあごはいわゆる「鋏(はさみ)の原理」で動き、多くの種類の歯はナイフのように鋭くとがっていた。雑食性のダチョウ恐竜と草食性の竜脚類も鋏式原理のあごの仕組みであった。竜脚類はものをかみ砕く奥歯をもたず、前方の釘(くぎ)のような切歯で、木の芽や実、小枝、木の葉を食いちぎって、かまずに飲み込んでいたらしい。このことから、消化管の一部に小石を飲み込んでもっていて、餌をつぶすのにそれ(胃石、ガストロリス)を役だたせ、強力な胃液で消化していたと推測されている。
鳥盤類はすべて草食性の歯をもっていたが、あごの仕組みも「クルミ割り方式」とよばれるもので、幅広いあご関節の位置は歯槽の線よりずっと低かったので、上下の歯がほぼ同時に接触し餌を押しつぶすようにして食べることができた。草食恐竜の餌は、当時繁茂していたシダ植物と裸子植物が主であったが、被子植物が出現した白亜紀後半は草食性の鳥盤類の全盛時なので、被子植物も餌になったであろう。カモノハシ竜ハドロサウルスが食べたものが、フランクフルトのゼンケンベルグ博物館の標本であるミイラの一つの胃の中身として残っているが、これは1922年にドイツのリヒャルト・クロイゼルRichard Kräusel(1890―1966)によって発表されたものである。それによると、胃の中身は針葉樹類のカニンガミテスの針葉のかたまりと、ほかに陸上植物の小枝や種および果実などであった。しかし、カナダで発見されたハドロサウルス類のミイラの「胃の内容物」には植物破片と木炭が混じっていたので、これらは死体中に流れ込んだのではないかという。また別の個体では胃の場所にチョウザメが発見されたが、これは死体内部を食べようとして入ってきた魚ではないかとされる。カリーは以上のような例をあげて、判断を誤らぬようにと注意を促している(1994)。
獣脚類が草食恐竜を襲ったと考えられる例は多く知られている。たとえば、アメリカ、ワイオミング州ボーン・キャビンで発見された草食恐竜アパトサウルスでは、かみ砕かれたと思われる尾椎突起と肉食恐竜アロサウルスの歯がいっしょに見つかっている。またモンタナ州で発見されたアパトサウルスの骨についた歯型とアロサウルスの歯、およびユタ州クリーブランド・ロイドで発掘されたアロサウルスと草食恐竜の骨の割合や産状の調査などから推測されるのである。
ロンドンの自然史博物館には、草食恐竜テノントサウルスTenontosaurusを襲う肉食恐竜デイノニクス数頭の動刻(動く彫刻)が展示されたことがある。テノントサウルスはデイノニクスの主要な獲物であったらしく、多くの場合デイノニクスの歯を伴って発掘されているという。デイノニクスが集団で狩りをした証拠とされるが、捕食者側の犠牲数が多すぎるなどの不自然さがあり、死後、洪水などで偶然いっしょになったとも考えられる。
1971年にゴビ砂漠を探索していたポーランド・モンゴル隊は、組み合ったまま砂嵐(すなあらし)のなかでともに力尽きて倒れてしまったものと思われる2頭の恐竜化石を発見した。これは明らかに格闘していたもので、小形で敏捷(びんしょう)な肉食恐竜ベロキラプトルが、小形の草食恐竜プロトケラトプスの頭部を前肢でつかんでいるものであった。このレプリカ(模造品)標本が、群馬県神流(かんな)町の恐竜センターと福井県勝山(かつやま)市の恐竜博物館に展示されている。
変わった食事の直接証拠もある。アメリカのニュー・メキシコ州産の獣脚類コエロフィシスCoelophysisの多数の標本のうちの二つは、体腔(たいこう)中に同種の小標本の骨をもっていた。これは赤ん坊ではなく、共食いした可能性が高いとされる。イギリスの白亜紀前期のバリオニクスBaryonyxはワニのように長い口先をもつだけでなく、魚の鱗(うろこ)が肋骨(ろっこつ)域に発見されたため、魚食向きに特殊化したのであろうと考えられている。また、アメリカ自然史博物館に展示されている標本のなかに、大形の肉食獣脚類が巨大な竜脚類に忍び寄ったと考えられる一連の足跡が展示されている。アメリカのテキサス州で発見された、獣脚類が竜脚類を襲ったのであるとされた足跡の現場には、逃げるためのスピード・アップや争いの跡はみいだせないので、襲ったことの証明にはならず、単に追いかけたり忍び寄ったりしたものと解釈される。
ある地域に生息する動物の種類別の割合は、それらの動物の食性を推定するうえで有力な方法となる。ある地質時代のある地域の恐竜の種類とその個体数の見積りはどのように行えるであろうか。将来は、地質学的・古生物学的資料を基に、堆積(たいせき)盆地ごとに、生物の多様性、種構成、優占種、生物量、生産力を具体的に分析し、群集構造や機能を考察し、個体群データから生態ピラミッド(総エネルギーと推定個体数)や食物網が議論されるようになるであろう。
[小畠郁生]
恐竜の病気は、もちろん骨化石に異変が残されていれば判別できるが、骨に影響を及ぼさない病気についてはほとんど調べることができない。しかし恐竜の歩行跡を調べると、足が不自由であった恐竜もいたことがわかる。そのような足跡は松川正樹やロックレイによる調べでは、2003年現在、世界中で10例知られているだけである。そのうち9例は、歩行跡の左足から右足までの距離と、右足から左足までの距離、すなわちステップの長さが異なる。ステップの長さが違うということは、左足から右足への運びと、右足から左足への運びのスピードが異なるということである。どちらかの足が不自由であったのだろう。肉食恐竜の獣脚類のものが5例、草食恐竜の竜脚類と鳥脚類のものがそれぞれ2例ずつある。獣脚類は狩りを行うので、けがをする割合が多かったり、縄張り争いによるけがをしたりしたのかもしれない。10例のうち残りの1例は、普通なら存在する3本の指のうち1本を欠いた足跡である。イギリスの三畳紀層から発見された獣脚類の足跡で、右後ろ足の内側の指がなかったが、けがによるのか先天性異常なのかは不明である。
さて、骨化石で病変をみつけるに際しては、病気と関係のない骨の変化、つまり続成作用(堆積物が堆積盆で沈積後、固結して堆積岩になる過程)による変化を識別しなければならない。紛れもない病変と解釈できるのは、治癒した証拠のある骨折だけである。1990年代以降、恐竜の古病理学は急速に進展しつつあるので、ここではおもにロスシルドBruce M. Rothschild(1947― )による論評(1997)を紹介しておく。骨の外皮(骨膜)の一部が引きはがされたとすると(腱(けん)が骨からとれた場合)、骨のその部分は骨片が突き出てぎざぎざとなり、そこに新しい骨が形成される。この新しくできた骨は外骨腫(しゅ)とよばれ、骨に軟骨片が含まれていると、骨軟骨腫(軟骨性外骨腫)という軟骨で包まれた外骨腫ができる。このような骨の過形成が行われた例は、トリケラトプスの下顎骨(かがくこつ)と肩甲骨、アロサウルスの肩甲骨の外骨腫で確認されている。
ハドロサウルス類の指骨関節面に、えぐられたようにみえるディボットと称するくぼみが発見されたことがあったが、これは軟骨の発育不全、つまり骨化しなかった軟骨細胞群の跡かもしれないとされている。同様な穴状構造がティラノサウルスの上腕骨でも発見されているが、これはちょうど腱が付着する場所にあったので、腱が骨から引きはがされた跡らしいといわれる。
骨折の場合には、治癒に要する時間のことを考えると、骨折後数週間以内の傷は、生時に受けたものか死後に受けた変化かの判定ができない。よく知られている骨折の例は、たとえば竜脚類の肋骨骨折、鳥脚類カンプトサウルスの腸骨の傷や尾の付け根の骨折、イグアノドンの坐骨骨折などである。獣脚類の例は珍しくなく、アロサウルスの前腕の橈骨(とうこつ)骨折や、シンタルススの蹠骨(しょこつ)骨折、デイノニクスの指骨骨折、アルバートサウルスの肋骨と腓骨(ひこつ)(下腿骨(かたいこつ))骨折、上腕骨の病変などが報告されており、ティラノサウルス類の上腕骨骨折も知られている。獣脚類の標本を調べた結果、前肢と足の骨折が27%と25%の割合でみられたが、これは獲物と格闘中に受けた損傷と思われる。これに対してトーマス・バンスThomas Vanceは前肢の骨折は交尾の最中に生じたものではないかと述べた(1989)。
一般に体重を支える主要な骨が骨折すれば生き延びられないというのが恐竜の世界と思われがちであるが、治癒して生き延びた例が報告されている。たとえば、シンタルススで脛骨(けいこつ)(下腿骨)骨折が治癒した例や、アロサウルス、アルバートサウルス、ティラノサウルスの腓骨の骨折が治癒した例である。このような病変が認められた獣脚類のうち頭骨あるいは下顎骨に異常があるものが13%、脊柱あるいは肋骨に異常があるものが27%あったという指摘もあるので、一生の間にかなりけがをすることがあったのであろう。
保存状態のよい全身骨格のみを観察するときには病的異常の度合いを実際より低く見積もりがちで、逆に病変をもつ標本を集めることばかりしていると、病的異常の頻度を高く見積もってしまう。そこで、化石産地を丹念に調査し恐竜類のボーンベッドから全標本を採集し、恐竜類の骨折を体系的かつ量的に研究したダレン・タンケDarren H. Tankeによる実例がある。カナダのアルバータ州の白亜紀後期の地層から発見した角竜類と鳥脚類ハドロサウルス類の、合計3万本の骨が対象となった。その結果では、セントロサウルスの骨の0.025~0.5%、パキリノサウルスPachyrhinosaurusの骨の0.2~1.0%に骨折がみられ、その大半は背肋(はいろく)(胸椎)の中部と後部に集中していた。これはアメリカバイソンの成体の雄の骨折分布とそっくりで、横腹に頭突きをする習性によると推測された。角や襟飾りの破損も同一種内の闘争を示す証拠と考えられ、トリケラトプス、トロサウルスTorosaurus、ペンタケラトプスPentaceratops、ディケラトプスDiceratopsなどで確認されている。一方、ハドロサウルス類では、尾骨で胴に近いほうの神経棘(きょく)骨折と尾骨の融合が目だつが、これらは交尾行動に関係があるらしい。雌の尾と、きゃしゃなつくりの神経棘の先端に雄の体重がかかった結果であると推測されている。かつてカンプトサウルスやイグアノドンでみつかった同様の傷も交尾のせいであるとされている。
運動の準備のできていないところへストレスが繰り返しかかると、骨に本来備わっている強度を超えた場合、微小破壊がおこって骨の再構築が促進され、ストレス骨折(疲労骨折)が生じるといわれる。骨の破壊を伴わないが、X線では線状にみえるストレス骨折が、角竜類の指骨でみつかっている。確認された箇所は、セントロサウルス、スティラコサウルスStyracosaurus、パキリノサウルスの第2指から第4指の指骨の足首に近いほうである。骨皮質の外側の層が肥厚する骨膜反応を伴うので、骨軟化症(ビタミンD欠乏症)を疑う必要はない。足で地面を強く踏みつけたからなのか、捕食者から逃げるために突然加速したからなのか、または長距離移動の最中に生じたものなのかは特定できない。
恐竜の歯は一生の間に何度も生え変わった。肉食のティラノサウルスの例では、波が伝わるように一定のルールで変わり、偶数番目の歯と奇数番目の歯の生え変わりの波がずれていた。子供のときには1年半に一度、大人になると2年に一度生え変わるので、50歳まで生きたとすると、一生で1250本以上の歯をもったことになる。歯の病気はめったになかったようで、虫歯が報告された例は聞かない。ただし、ラムベオサウルスのあごにみられた歯の膿瘍(のうよう)では、腫(は)れあがった深いくぼみがあって、歯列の溝が壊れ、歯が欠損していたという報告がある。獣脚類ではときどき歯が壊れていることがあり、アロサウルスの標本では歯がなくなっているものもある。ティラノサウルスの上顎骨で不正咬合(こうごう)が確認されたり、ノドサウルス類のエドモントニアEdmontoniaの前上顎骨右側に傷があったりしたとの報告もある。
恐竜では関節の感染症が確認されたことはないし、骨の感染症すなわち骨髄炎もほとんどみられない。感染症にかかることが珍しかったのか、かかってしまうと生き残れなかったのかのどちらかである。ハドロサウルス類の前肢末端部に骨折と感染がみられたという報告があるように、骨折時に骨が薄い外皮を突き破り、複雑骨折の傷が細菌に感染したのであろうといわれる。カンプトサウルスで腸骨が骨髄炎をおこした例では、同一箇所にかまれたらしい跡が残っているので、これが原因のように思われる。数少ない例であるが、膿瘍のできたディロフォサウルスの上腕骨、アロサウルスの指骨、トロオドンの頭頂骨などの報告がある。角竜類の肩甲骨で骨の外皮の感染症つまり骨膜炎がおきた例や、トリケラトプスのくちばしで、感染症が原因と思われる突起の破損が確認されている。
骨関節症はイグアノドンで2例確認されている。関節に骨棘(こっきょく)すなわち棘突起が成長しているのである。関節ではなく椎間板のスペースに椎体の棘突起が形成されているため痛みや病的状態を引き起こすことはなく、単なる変形性脊椎症である。ハドロサウルス類、よろい竜類、獣脚類、剣竜類、厚頭竜類、竜脚類では、骨関節症は未発見である。竜脚類に骨関節症がみられないのは、恐竜の体重の重さが骨関節症の主因とは思えないことを示す。安定度の高い関節では動きがもっぱら一つの可動面に限られ、骨関節症にかかりにくいのに対し、複雑な動きのできる関節はもっともかかりやすいという。恐竜で骨関節症の発生率が低いのは、一般に関節の可動範囲に制約があって、回転運動がほとんどできない蝶番(ちょうつがい)関節であったからではないかとされている。
痛風(血液中の尿酸値が高くなった高尿酸血症から急性の関節炎をおこす疾患群)は肉食恐竜2体で確認されている。アメリカのサウス・ダコタ州産のティラノサウルスの前肢骨2本が、骨組織に痛風の特徴を呈していた。骨の表面が回転楕円(だえん)状に侵食され、その縁に新しい骨ができている。アルバータ州産のティラノサウルス科恐竜の足指の骨にも同様の徴候が認められた。栄養価が高いものばかり食べていたせいであろうか。
脊椎の長軸方向に走る靭帯(じんたい)が骨化したものは汎発(はんぱつ)性突発性骨増殖症ないしはフォレスチャー病ともよばれる。ところが、脊椎靭帯の骨化はほとんどの恐竜ではあたりまえの現象となっている。角竜類、ハドロサウルス類、イグアノドン類、厚頭竜類は、幼体の時期に骨化したと思われる靭帯をもつ。こうした機構で尾が強化されていた。よろい竜類で唯一融合がみられるのは尾に骨質のハンマーをもつ種類で、効果的な武器となっていた。ノドサウルス類のよろい竜類エドモントニアの尾部が融合してよじれた尾をつくった例がある。この個体には前上顎骨の右側に傷もあり、外傷が原因で異常を引き起こしたらしい。ディプロドクスで尾椎の2~4本に融合がみられるのに脊椎関節突起(小関節面)の結合が無傷の状態に保たれていた。コンピュータ断層写真を撮った結果、このような標本では骨化した靭帯と椎骨椎体の間が分離していることがわかった。靭帯の骨化は尾椎の17~23番に限られ、尾の構造を補強し尾を持ち上げやすくしていたし、尾を防御手段として使うのに役だったのかもしれない。縄張り争いや求愛行動とかかわり、交尾にはかかせなかったのであろう。1991年までに調べられた標本のうちディプロドクスとアパトサウルスの50%、カマラサウルスの25%にこの現象が存在するので、性的二型性を表すのではないかと思われている。雌の尾が補強されれば、総排出腔を交尾しやすい位置に保つことができたであろう。
角竜類では最初の3個の頸椎が融合した例が記録されている。当初は脊椎関節症とみなされたが、複数の属の成体すべてにこの現象が認められたため、現在では発育上の現象であろうと考えられ、巨大な頭骨を支えるという力学的機能からといわれる。成体と思われる大形の角竜類とは対照的に小形で幼体と思われる角竜類ではこの頸椎の融合が不完全である。レプトケラトプスLeptoceratopsの頸椎は明らかに分節し、プロトケラトプスの頸椎は放射線で調べてみたところ分節していた。パキリノサウルスは、大形の成体標本では多様な融合が記録されている。融合は個体発生上のできごとで、成体になった証拠かもしれない。ティラノサウルスの標本で、2個の脊椎の椎体が奇妙な形で融合している例があるが、融合の仕方が特殊で変形が生じているので、かまれた傷が治った結果とも考えられる。外傷が原因で尾先が融合した例はハドロサウルス類とアロサウルスで記録されている。
かつてロイ・リー・ムーディーRoy Lee Moodie(1880―1934)は、竜脚類の尾骨融合は血管腫を示していると述べた(1923)が、発表された図では、X線写真で長軸方向の梁(はり)構造が目だったり蜂(はち)の巣状構造を示したりというような血管の腫瘍のようすはみられなかったため、ロスシルドは靭帯が骨化したものが汎発性突発性骨増殖を行った可能性が高いと述べる(1997)。獣脚類(アロサウルスまたはトルボサウルスTorvosaurus)の上腕骨にカリフラワー状の盛り上がりが認められ、これは軟骨肉腫とよばれる軟骨腫ではないかとの報告があるが詳細は不明である。
恐竜の卵も病気に無縁ではない。カール・ヒルシュKarl F. Hirsch(1921―1996)らの報告(1989)では、ジュラ紀の恐竜の卵から、殻に石灰化した層が2層あるものを発見したことから、この状態では呼吸が妨げられ、中の胚(はい)は生き延びられなかったと予想されたが、卵をコンピュータ断層撮影で調べたところ、胚は正常であったという。
[小畠郁生]
地史を調べてみると、どの生物のグループも生成、変化、発展、絶滅の歴史を示している。恐竜たちも生物のグループとして消滅の段階に達していたのであろうか。しかし、白亜紀末には、いろいろな種類の恐竜ばかりでなく、翼竜や海の爬虫類、頭足類のアンモナイトなどいろいろな種類が同時に絶滅している。したがって、そういう状態に追い込んでいった共通の理由を、まず恐竜自身の外に求めてみるという諸説がある。伝染病流行説というのがあったが、恐竜およびそれ以外の各種類の動物をすべて根絶させるという伝染病などがありえたかどうかきわめて疑わしい。白亜紀に出現した被子植物に含まれる有毒のモルヒネ、ニコチン、カフェイン、コカインなどのアルカロイドが、それを食べた恐竜に蓄積されて、その悪影響が子孫に現れてくるというアルカロイド中毒説というのがあった。しかし、魚や頭足類を食べていた翼竜も首長竜もモササウルスMosasaurusも一斉に姿を消しているし、アルカロイドを含む植物が現れてから恐竜が滅びるまでに3000万年以上もかかっていることが事実である限り、中毒説にはうなずけない。
ある恐竜化石の放射能を測定したら異常に高いカウント数を示したので、その原因になった重要なできごとが白亜紀末にあったとする説がある。また過去の地質時代に周期的に熱風の吹きまくった時代があり、とくに白亜紀末にそれが著しく、そのとき、小さな哺乳類は岩の割れ目などに隠れてその難を逃れたが、大きくて鈍重な恐竜はなすすべもなく死滅するしかなかったという熱風説もあった。1054年に中国(宋(そう))の天文学者が観測したような超新星の爆発は非常に遠かったため地球に害を及ぼさなかったが、たとえば太陽系から100光年ほどの位置にある星が爆発したとすると、地球に降り注ぐ放射線量は現在の10万倍にもなり、破滅的影響を与えたに違いない。天文学的な確率計算では、超新星爆発は銀河系宇宙で50年に1回ほどおきており、地球から100光年ほどの位置でおこる確率も数億年に1回ほどあるといわれる。恐竜は放射線を大量に浴び細胞が破壊されたとしても、ワニやカメや小形哺乳類はどうしたのであろうか。放射線を遮断する海水下のアンモナイトやプランクトンはなぜ絶滅したのであろうか。
このような説を成立させるためには、過去の地質時代のその時期(白亜紀末)に、そのような天文学上および気象学上の変化が確実にあったという十分な証拠と、それが地球上のいろいろな生物に及ぼした直接の影響を示す確実な証拠が必要である。その意味で、1980年に提唱された隕石(いんせき)説は、従来の諸説に比して地球上に残された証拠らしいものがある点で、注目すべきである。すなわち、デンマークやイタリアなどの白亜紀層と古第三紀層の境に、隕石起源と思われるイリジウムが多量に含まれる厚さ数センチメートルの地層が存在することが確認された。衝撃石英やテクタイト(衝突された地点の岩石に含まれていた物質が融(と)けて飛散したもの)も発見された。白亜紀末に大隕石が海中に落下し、津波が生じ、塵埃(じんあい)が数年にわたり太陽の光を遮り、植物は光合成作用を営めず草食恐竜がまず滅び、やがて肉食恐竜も滅びたというのである。しかし、ここでもやはり、ワニ類、カメ類、鳥類、哺乳類などの生き延びた理由については別の説明が必要となる。
そこで、恐竜が絶滅した理由については、やはり長い歴史を通じてつくられてきた恐竜自身の体に問題があったと考える一方で、周りの生物を含めて、環境からの働きかけと環境への適応はどのようであったか、個々の生物の生活の仕方ということまで含めた総合的な解釈でなければならない、ということになる。白亜紀末の世界的規模の海退は動物のすみかに影響を与えた。気候帯や四季の変化で周囲の温度が下がると、冷血の爬虫類は動きが鈍り、生理機能は低下し、冬眠状態に入ったであろう。これに対し、恐竜温血説の説くところでは、恐竜は冬眠できなかった。体温を一定に保たないと生きていられないのに、毛皮のような保温のための手段をもたなかったからである、とされた。
さらに興味深い点は、1957年にフランスのプロバンス地方より産出した恐竜の卵の産状と卵殻およびその微細構造について研究した、フランスのレイモン・デュギM. Raymond Dughi(1898―1977)とフランソア・シルグFrançois Sirugueの報告によると、白亜紀層中の下位の層準にあたる古い地層では、正常な形態を示す卵を産するのに対し、上位の層準、つまり新しい地層となると異常に厚い卵殻をもつ卵化石の産出頻度が高くなるというものであった。彼らはこのことについて、気温の急激な低下がこの病理的現象を招いたと考えた。ドイツのハインリヒ・カール・エルベンHeinrich Karl Erben(1921―1997)が1970年に発表したところでは、異常に厚い卵殻の微細構造を電子顕微鏡で見ると、病的な卵殻の大部分には発育の中断が認められ、その部分に有機質の外被が挿入されていることがわかった。恐竜の卵殻はワニ類やトカゲ類のものよりも鳥類のものに類似する。鳥類のなかでもニワトリの場合、低温などが影響したりストレスを与えたりすると、ホルモンの異常分泌のためにかなり多数の卵殻が正常卵に比べて異常に厚くなる。このような卵殻では気孔が閉塞(へいそく)されて胚が窒息し、かりに子供が成長したとしても、卵殻を破って孵化することは不可能となる。このように環境変化と、それに由来する生物体自身の生理の変化が多角的に影響したのであろう。なお現生爬虫類のなかのあるものは性染色体をもたず、性別が孵卵(ふらん)のときの温度で決定されることがわかっている。たとえば、アカウミガメの卵は低温でかえると雄になり、ワニやトカゲのある種では低温では雌になる。恐竜類でも白亜紀末の気温低下で雌雄の偏りが著しくなったかもしれない。
隕石衝突説の弱点とされたのは、提案時にはイリジウム量から想定される小惑星がつくったとみられるクレーターが特定されなかったことである。ところがメキシコのユカタン半島にみられる重力異常と地磁気異常の発見(1981)が、チクチュルブ・クレーター発見として脚光を浴びるようになったのが1991年で、深さ1100メートル、直径200キロメートルに及ぶ埋もれた衝突クレーターの存在が示唆された。隕石の大きさは10キロメートルを超えるものであった。半島は石灰岩が分布するので、これの蒸発で二酸化炭素を生み、温暖化がもたらされる。石膏(せっこう)の蒸発は二酸化硫黄(いおう)を生み、やがて酸性雨をもたらす。調査結果ではクレーターの直径は300キロメートルあるいは180キロメートルで、衝突時はフロリダ半島に達する爆風、地球を覆う地震波と津波、地球をとりまく岩石の塵(ちり)、月夜の10分の1の暗さ、衝突のもたらす長い冬、濃硫酸の雨、オゾン層の消滅、数十万年続く温暖化などが推定される。
しかし、恐竜やアンモナイトの種類が漸減する一方、原始有蹄(ゆうてい)類数が漸増する点を強調する人もいる。実際には、白亜紀末の生物種の大量絶滅は、温度低下と海退、火山活動、大陸の接続に伴う種の接触に、隕石などの激変的要素が加わって、大規模のものになったらしい。
最後に強調しておかねばならないのは、大量絶滅を免れた生物側の反応である。一般に生きていくための所要エネルギー量が大きい恒温動物では、ネズミ大の小形哺乳類と小鳥だけが生き延び、それより大きかった恒温の小形肉食恐竜や翼竜は滅び去った。食物量が少なくてすむ変温動物ではワニぐらいの大きさのものが上限で、それより大きな竜脚類や鳥盤類、海中の首長竜やモササウルス類など大形の動物は絶滅してしまったという事実がある。これは、白亜紀末の環境の大異変に直面した動物にとって、生命を維持していくのに要した最小エネルギー量がどの程度であったかを明らかに示唆しているものと思う。同程度の大きさのものでは、食物量の摂取を含め、生活の仕方の違いが絶滅か存続かを決めるのに大きく影響したに違いない。
なお、恐竜は鳥類と共有する特徴が120以上もあり、20世紀後半に盛んになった分岐分類学では鳥類は恐竜目の下位グループなので、恐竜は絶滅していないともいえることになる。
[小畠郁生]
『W・E・スウィントン著、小畠郁生訳『恐竜 その発生と絶滅』(1972・築地書館)』▽『E・H・コルバート著、長谷川善和訳『恐竜はどう暮らしていたか』(1981・どうぶつ社)』▽『デイヴィッド・ノーマン著、浜田隆士監修『恐竜』(『動物大百科 別巻』1988・平凡社)』▽『ジャン・ギィ・ミシャール著、阪田由美子訳、小畠郁生監修『恐竜のすべて』(1992・創元社)』▽『ピーター・ローソン著、三宅真季子訳『恐竜学最前線4 ティランノサウルス・レックスの性的二型性 スー物語 第3部』(1993・学習研究社)』▽『デビッド・ノーマン著、青木久美子・滝寺治子訳『恐竜』(1993・マール社)』▽『ティム・ガードム著、伊藤恵夫・舟木秋子訳『英国自然史博物館 恐竜』(1994・ほるぷ出版)』▽『デヴィッド・ランバート著、瀬戸口美恵子・月川和雄訳、瀬戸口烈司監修『恐竜解剖図鑑』(1994・同朋舎出版)』▽『佐藤哲、ネイチャー・プロ編集室構成・文『恐竜の行動とくらし8 日本の恐竜――どんな恐竜がどのような環境にくらしていたのか』(1995・偕成社)』▽『マーク・A・ノレル、ユージン・S・ギャフニー、ロウエル・ディンガス著、瀬戸口烈司・瀬戸口美恵子訳『恐竜の博物館』(1996・青土社)』▽『デイヴィッド・B・ワイシャンペル、ルーサー・ヤング著、瀬戸口烈司・瀬戸口美恵子訳『アメリカ東海岸の恐竜たち』(1998・青土社)』▽『デイヴィッド・E・ファストフスキー、デイヴィッド・B・ワイシャンペル著、瀬戸口美恵子・瀬戸口烈司訳『恐竜の進化と絶滅』(2001・青土社)』▽『小畠郁生監訳『恐竜大百科事典』(2001・朝倉書店)』▽『平山廉著、小田隆復元画『図解雑学 恐竜の謎』(2002・ナツメ社)』▽『ナショナルジオグラフィック編『恐竜の世界』(2004・日経ナショナルジオグラフィック社、日経BP出版センター発売)』▽『日経サイエンス編集部編『別冊日経サイエンス145 地球を支配した恐竜と巨大生物たち』(2004・日経サイエンス社)』▽『ヘーゼル・リチャードソン著、出田興生訳『恐竜博物図鑑』(2005・新樹社)』▽『富田京一編著『すべてわかる恐竜大事典』(2006・成美堂出版)』▽『ヘンリー・ジー、ルイス・V・レイ著、小畠郁生監訳、池田比佐子訳『恐竜野外博物館』(2006・朝倉書店)』▽『デイヴィッド・E・ファストフスキー、デイヴィッド・B・ワイシャンペル著、真鍋真監訳『恐竜学 進化と絶滅の謎』(2006・丸善)』▽『NHK「恐竜」プロジェクト編、小林快次監修『恐竜vsほ乳類――1億5千万年の戦い』(2006・ダイヤモンド社)』▽『小畠郁生著『恐竜はなぜ絶滅したのか』(岩波ジュニア新書)』▽『冨田幸光著『恐竜たちの地球』カラー版(岩波新書)』▽『フィリップ・カリー著、小畠郁生訳『恐竜ルネサンス』(講談社現代新書)』▽『平山廉著『最新恐竜学』(平凡社新書)』▽『イワン・アントノヴィチ・エフレーモフ著、清水邦生訳『恐竜を求めて――風の道』(『現代世界ノンフィクション全集16』1967・筑摩書房)』▽『ロジジェストウェンスキィ著、堀江豊訳『ゴビに恐竜をもとめて』(1974・築地書館)』▽『ジョン・R・ホーナー著、小畠郁生訳『子育て恐竜マイア発掘記』(1989・太田出版)』▽『ドン・レッセム著、加藤珪訳『恐竜発掘』(1993・二見書房)』▽『R・C・アンドリュース著、小畠郁生訳『恐竜探検記』(1994・小学館)』▽『ジョン・R・ホーナー、ドン・レッセム著、加藤珪訳『大恐竜 T・レックス新発見』(1994・二見書房)』▽『マイケル・ノヴァチェック著、瀬戸口烈司・瀬戸口美恵子訳『ゴビ砂漠の恐竜たち』(1997・青土社)』▽『ウィリアム・ナスダーフト、ジョシュ・スミス著、奥沢駿訳、真鍋真監修『失われた恐竜をもとめて――最大の肉食恐竜をめぐる100年の発掘プロジェクト』(2003・ソニー・マガジンズ)』▽『ピーター・ラーソン、クリスティン・ドナン著、冨田幸光監訳、池田比佐子訳『Sue スー――史上最大のティラノサウルス発掘』(2005・朝日新聞社)』▽『チャールズ・ガレンカンプ、マイケル・J・ノヴァチェック著、岩井木綿子・中村安子・藤村奈緒美訳『ドラゴンハンター――ロイ・チャップマン・アンドリューズの恐竜発掘記』(2006・技術評論社)』▽『ウェイン・グレイディ著、ヒサクニヒコ訳『史上最大の恐竜発掘 カナダ・中国共同プロジェクトの成果』(新潮文庫)』▽『真鍋真・小林快次編著『日本恐竜探検隊』(岩波ジュニア新書)』▽『小畠郁生編『恐竜の時代を復元する』(1977・出光書店)』▽『小畠郁生編著『恐竜時代の生物と自然』(1981・築地書館)』▽『B・クルテン著、小畠郁生訳『恐竜の時代』(1983・平凡社)』▽『シルヴィア・J・ツェルカス、スティーヴン・A・ツェルカス著、小畠郁生監訳『恐竜 地球環境からみた恐竜の進化と絶滅の物語』(1991・河出書房新社)』▽『松井孝典著『地球大異変――恐竜絶滅のメッセージ』改訂版(1998・ワック)』▽『金子隆一著『大絶滅。』(1999・実業之日本社)』▽『D・ディクソン著、小畠郁生監訳『生命と地球の進化アトラスⅡ』(2003・朝倉書店)』▽『速水格「絶滅の生態学的考察」(小澤智生・瀬戸口烈司・速水格編『古生物の科学4 古生物の進化』pp.175~193. 2004・朝倉書店)』▽『平野弘道著『絶滅古生物学』(2006・岩波書店)』▽『小畠郁生「適応放散の拡がり――極大の世界」(小林直樹・小原秀雄・柴田義松編『総合人間学の試み2――生命、生活から人間を考える』pp.57~72. 2006・学文社)』▽『ティム・ヘインズ著、池田比佐子訳、小畠郁生監訳『図説 恐竜の時代』(2006・岩崎書店)』▽『小畠郁生著『恐竜の時代』(社会思想社・現代教養文庫)』▽『E・H・コルバート著、小畠郁生・沢田賢治訳『さまよえる大陸と動物たち』(講談社・ブルーバックス)』▽『松川正樹著『恐竜ハイウェー 足跡が明かす謎の生態』(PHP新書)』▽『A・J・デズモンド著、加藤秀訳『大恐竜時代』(1977・二見書房)』▽『L・B・ホールステッド著、亀井節夫監訳『ディノサウルス』(1981・築地書館)』▽『J・C・マクローリン著、小畠郁生・沢田賢治訳『恐竜たち』(1982・岩波書店)』▽『ジョン・ノーブル・ウィルフォード著、小畠郁生監訳『恐竜の謎』(1987・河出書房新社)』▽『ロバート・T・バッカー著、瀬戸口烈司訳『恐竜異説』(1989・平凡社)』▽『グレゴリー・S・ポール著、小畠郁生監訳『肉食恐竜事典』(1993・河出書房新社)』▽『シルヴィア・J・ツェルカス、エヴァレット・C・オルソン編、小畠郁生監訳『恐竜 過去と現在』Ⅰ・Ⅱ(1995・河出書房新社)』▽『R・M・アレクサンダー著、坂本憲一訳『恐竜の力学』(1991・地人書館)』▽『小畠郁生編『恐竜学』(1993・東京大学出版会)』▽『山崎信寿著「生体力学からのアプローチ」(NHKスペシャル取材班編『これが恐竜だ! 科学的アプローチによる新しい恐竜像』所収・1995・新潮社)』▽『クリストファー・マクガワン著、月川和雄訳『恐竜解剖 動きと形のひみつ』(1998・工作舎)』▽『笹沢教一著『恐竜が動きだす デジタル古生物学入門』(中公新書ラクレ)』▽『デニス・R・ディーン著、月川和雄訳『ギデオン・マンテル伝 恐竜を発見した男』(2000・河出書房新社)』▽『デボラ・キャドバリー著、北代晋一訳『恐竜の世界をもとめて 化石を取り巻く学者たちのロマンと野望』(2001・無名舎)』▽『クリストファー・マガウワン著、高柳洋吉訳『恐竜を追った人びと――ダーウィンへの道を開いた化石研究者たち』(2004・古今書院)』▽『エドウィン・H・コルバート著、小畠郁生・亀山龍樹訳『恐竜の発見』(2005・早川書房)』▽『ダグラス・パーマー著、小畠郁生監訳、加藤珪訳『化石革命――世界を変えた発見の物語』(2005・朝倉書店)』
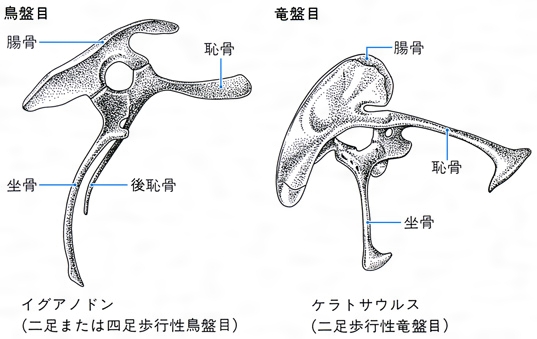
竜盤目と鳥盤目の骨盤の形〔図A〕
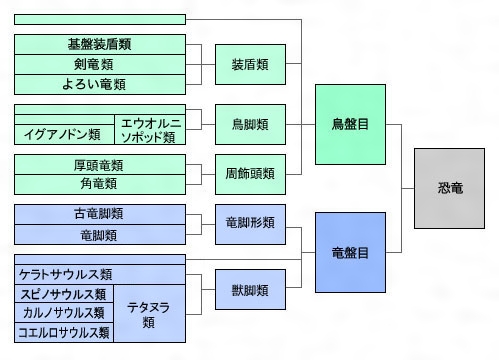
恐竜の系統樹〔図B〕

恐竜の系統樹(詳細図)〔図C-1〕

恐竜の系統樹(詳細図)〔図C-2〕
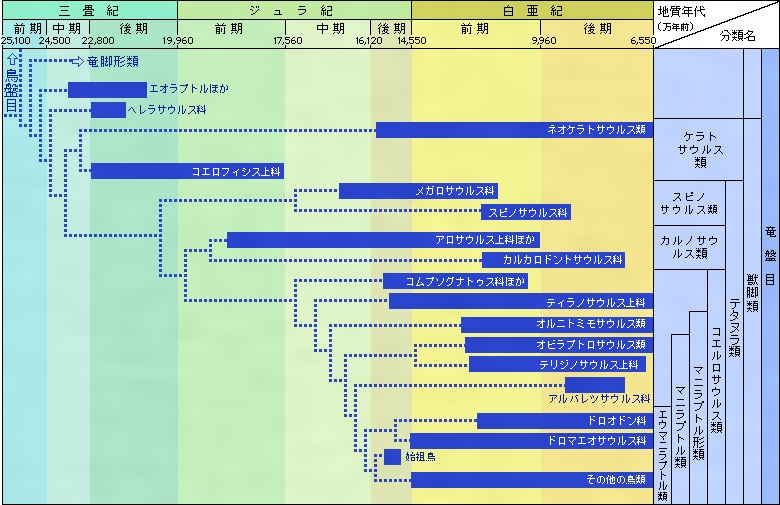
恐竜の系統樹(詳細図)〔図C-3〕
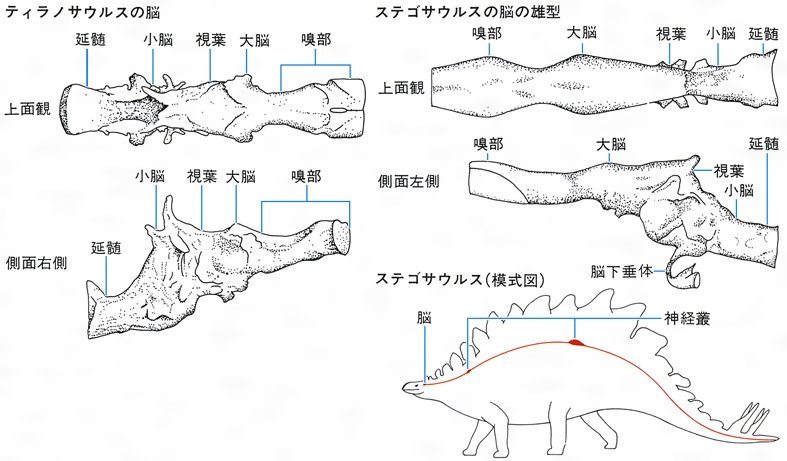
恐竜の脳〔図D〕
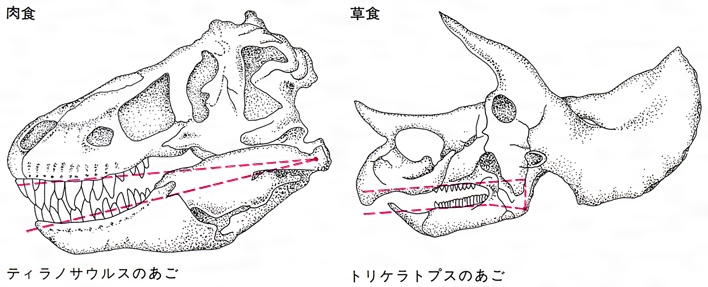
恐竜の歯の形とあごの仕組み〔図E〕

アロサウルスの骨格

イグアノドンの骨格
エオラプトルの骨格

カンプトサウルスの骨格

タルボサウルスの骨格
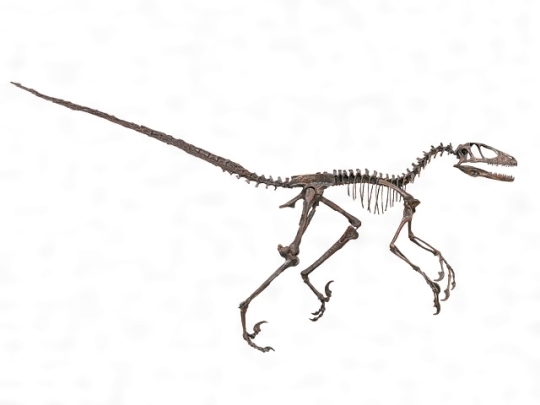
デイノニクスの骨格

ティラノサウルスの骨格

トゥオジャンゴサウルスの骨格

パキケファロサウルスの骨格

ヒプシロフォドンの骨格

プロトケラトプスの骨格
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
中生代に栄えた大型爬虫類の一群。dinosaurは1842年にイギリスのR.オーエンが提唱し,これは恐竜または恐蜴(きようえき)とも訳された。ギリシア語のdeinos〈おそろしい,ものすごい〉と,sauros〈トカゲ〉を合成した語。中生代の爬虫類全般に対して使うのは誤りである。頭蓋は幅狭く高い。頭頂の諸骨が退化し,頭骨の側頭窩(そくとうか)が左右それぞれ二つあり,それらが後眼窩骨と鱗状骨で隔てられている双弓亜綱Diapsidaの主竜(祖竜ともいう)下綱Archosauriaに属する。Archosauriaは恐竜群とも呼ばれ,ヒューネF.Huene(1948)の竜型群Sauromorphoideaにあたる。ローマーA.S.Romer(1950)は次の5目に分けた。
(1)槽歯目Thecodontia 三畳紀
(2)鰐目Crocodilia 三畳紀~現世
(3)翼竜目Pterosauria ジュラ紀~白亜紀
(4)竜盤目Saurischia 三畳紀~白亜紀
(5)鳥盤目Ornithischia 後期三畳紀~白亜紀
恐竜類はこのうちの,竜盤目と鳥盤目の2目を指す。竜盤目は恥骨が腸骨と坐骨から離れ前下方に伸びているもので,鳥盤目は恥骨と坐骨とが並行して接しているものである。
恐竜類の分類は人によりかなり違う。ローマー(1945)は竜盤目を2亜目15科150属に,鳥盤目を4亜目13科116属に分類した。オルシェウスキーG.Olshevsky(1978)は竜盤目を2亜目36科179属287種に,鳥盤目を5亜目13科121属209種に整理している。最近ではモンゴル,中国などアジアを中心に新しい恐竜が多く発掘され,獣脚亜目が細分化されたり,鳥盤目ではパキケファロサウルスの仲間を独立させる考えなどがあり,まだ混乱がある。恐竜類の2目はいずれも三畳紀後期に出現し,竜盤目は二脚歩行性のコエロフィシスCoelophysisから進化したといわれ,これは槽歯目(テコドント類)に起源すると考えられている。一つは大小さまざまな肉食性恐竜に発展し,一つは四脚歩行の大型植食性恐竜(竜脚類)に発展し,これらはジュラ紀に最大となった。鳥盤目は竜盤目とほぼ同じころに出現しており,その起源は槽歯目中の偽顎亜目に起源すると考えられている。鳥盤目のうち剣竜類と曲竜などは三畳紀にすでに分化したと考えられているがはっきりしない。その主流はファブロサウルスFabrosaurusをはじめとして,ヒプシロフォドンHypsilophodonやイグアノドン類で,白亜紀に入ってカモハシ竜類,角竜類やパキケファロサウルスなどが大量に適応放散した。それに対応して竜脚類も分化した。
恐竜類についての学問的な記載はイギリスにおけるメガロサウルスMegalosaurus(バックランドW.Buckland,1824)とイグアノドン(マンテルG.A.Mantell,1825)に始まる。ヨーロッパ大陸では,ベルギーのベルニサール炭鉱から産出したイグアノドンの骨格群集(ボーレンジャーG.A.Boulenger,1881)が重要な発見となった。北アメリカでは,コネティカット渓谷の三畳紀の足跡化石(ヒッチコックE.Hitchcock,1836)や,ハドロサウルスHadrosaurus類の研究(リーディJ.Leidy,1856)にはじまり,マーシュO.C.MarshやコープE.D.Copeらによるコロラド州のジュラ紀モリソン層の恐竜発掘と研究が活発に行われた。1958年には採掘場と露出した恐竜を恒久的に保存展示する施設をもった恐竜国立公園がユタ州にできた。カナダでは1880年代の初めにドーソンJ.W.Dawsonらによって調査された。カナダは角竜類の産地として知られるようになる。南アフリカでは海岸近くの白亜紀前期の地層から発見されたアルゴアサウルスAlgoasaurus(ブルームR.Broom,1904)の研究からはじまり,カルー盆地の三畳紀あるいはタンザニアのテンダグルのジュラ紀恐竜が有名である。テンダグルの発掘は1909-12年の4年間行われ,ブラキオサウルスBrachiosaurusやケントロサウルスKentrosaurusなど総重量250tの恐竜が発掘された。その後,1924年にイギリス隊による発掘がある。フランス隊によるマダガスカルの恐竜も重要である。南アメリカの最初の恐竜は1882年にパタゴニアで発見され,アメギノF.Ameghinoにより研究された。ラ・プラタの博物館にはパタゴニアのネウケンなどの白亜紀の恐竜化石が収集され,ヒューネによって研究された。アルゼンチンの三畳紀の地層から産したものは竜盤目で最古のものとして注目されている。
アジアでは,中国東北部アムール川沿岸より発見されたマンチュロサウルスMandschurosaurus(リアビーニンA.N.Riabinin,1925)にはじまる。本格的な調査はアメリカ自然史博物館が人類起源の研究のために送り出した調査隊が,中央アジアのゴビ砂漠でプロトケラトプスProtoceratopsや恐竜の卵を発見してからである。アメリカ隊(3回)の次はソビエト隊,ポーランド隊などが次々と発掘を行い,現在では中国の新疆ウイグル地区と並んで重要な恐竜産地となっている。南サハリンからはカモハシ竜に属する日本竜Nipponosaurusが1934年に発見され,長尾巧が記載した。日本列島では長崎県西彼杵郡高島町(現,長崎市)三菱高島礦業所二子立坑の地下917m地点の砂岩層中からカモハシ竜の上腕骨が1962年に出たと報告されている。本州初の恐竜は1979年に岩手県下閉伊郡岩泉町茂師の下部白亜系からアジア最大の竜盤目マメンチサウルスMamenchisaurusの1種が発見された。また1983年には最初の肉食性恐竜が熊本県から報告され,また群馬県からも産出した。
恐竜の脳の化石はないが,頭骨の脳腔を満たしている雄型から,脳の各部分の形態や脳から出ている神経の数・位置・太さなどを調べることによって神経系の機能について研究がされている。身体の大きさと比較するとひじょうに小さい。ステゴサウルスでは体重比が3000万g:300g,すなわち10万分の1である(ネコは100分の1)。ステゴサウルスの脳はクルミの実3個ほどの大きさしかない。構造からすると,嗅球からはじまり小脳まで際だった高まりはなく,細長くて低い。表面は滑らかでしわが発達していない。嗅覚と脳下垂体は比較的発達していて,現生のワニ類によく似ている。鼻孔は一般に大きく,ハドロサウルスではとくに鼻道が発達している。
目には強膜板が存在するが,単に目の保護のためにあるのか別の役割をもっていたかは不明である。一部のものは両眼視できたと考えられている。トカゲやカメ類は色彩の区別ができることから,恐竜類も色視の可能性が高い。耳の構造はワニ類に似ており,多くの種類に舌骨があることから,ある種の声を出すことができたと考えられている。鼓室には三半規管の機能がみられる。
歯牙は同形多生歯性である。歯数は多く,槽生で前顎骨および上顎骨の縁部,下顎の歯骨などに限られる。下顎結合は軟骨からなる。鳥盤目の前歯骨は多くは角質物に覆われくちばしを形成した。植食性のハドロサウルス類などでは歯のエナメル質が偏在する。数本並んだ歯列が何十列も並列し,各歯列の歯は萌出の時期をずらし,摩耗面で斜交してくる。萌出は車輪交換式である。総歯数は1000個をこえる。ティランノサウルスなどは各列大小2~3個の歯が舌側に萌出し,垂直交換の形式で発達する。皮膚はカモハシ竜やトロエドンTroödonなどのミイラ化石から,一部の恐竜類は不規則なうろこ状ないしこぶ状突起に覆われていたことが明らかであるが,大部分は不明である。アナトサウルスAnatosaurusでは多角形のモザイク状の薄い板ないしは結節の皮膚からできていた。体色や筋肉の構造はわからない。卵は一つの巣に複数産む。多くは固い卵殻よりなり,形は円形,長楕円形,まんじゅう形などさまざまである。卵を産んだ親の判明しているのはプロトケラトプスだけである。プロトケラトプスは円形の巣に長楕円形の卵を30~35個前後,少なくとも二重に産みつけていた。多くの場合,砂の中に産卵されている。卵殻の構造は種類によって異なる。しかし,鳥類の殻構造より弱かったと考えられている。大きさはその径が30cmをこえるものは知られていない。植食性恐竜は群れをなしていたことは明らかであるが,家族で群れを作っていた証拠はない。
剣竜類,巨大恐竜などでは仙椎の神経節が発達し,〈腰の脳〉を形成している。足および尾の運動を支配したものと考えられる。尾椎は数が多く長い。関節部の下面にはV字骨が発達する。鎖骨および間鎖骨を欠き,肩帯の構造は単純。腰帯は2系統に分かれる。四肢骨は変化が激しい。指は5本が基本で3本のものもいる。指先は鉤爪(かぎづめ)ないしひづめ状である。歩行は蹠行(しよこう)性ないし趾行(しこう)性である。二脚歩行で,肉食性獣脚亜目などには腹肋骨が発達する。外骨格の断面でみる骨質は外側は緻密(ちみつ)質が発達し,内側は海綿質構造が中心に向かってしだいに粗くなる。竜脚類ではしばしば胃石が発見される。おそらく食物の消化に役だっていたと考えられる。カモハシ竜などでは針葉樹などを食物としていたものが知られている。体の大きさは高さ18mのスーパーサウルスが最高で,体重100tと推定されている。最大長はディプロドクスの27m,最小のものではコンプソグナツスなどで,体長60cm前後。南極を除いたほとんど全世界に分布した。
恐竜の時代は中生代の全期間にわたる。三畳紀前期は,迷歯類という両生類の絶頂期にあり,河川にはワニ型動物フィトサウルスPhytosaurus類が,海には魚竜など遊泳性の爬虫類が出現した。陸上では哺乳類型爬虫類の最後の時代に入っていたが,恐竜の祖先となるテコドント類が栄え,その中から小型の,プラテオサウルスで代表される二脚歩行性のカンガルーに似た体つきの恐竜が出現した。木生シダ,シダ,トクサ,松柏類,ソテツ,イチョウなどが森林を作り,松柏類には数十mに達するものがあったと推定され,乾燥気候の地域がかなりあった。ジュラ紀には恐竜類が巨大化し,特殊化した。さらに,空域には翼竜類や鳥類が,海域では亀類,魚竜類,鰭竜(きりゆう)類などがより特殊化の道をたどっていた。北アメリカのモリソン層やアフリカのテンダグル層が代表とされる。植物は松柏類,イチョウ,木生シダやトクサが多く,常緑性森林を形成していた。三畳紀の気候より湿潤で,沼沢地が豊富だった。また,昆虫類が大発展し,現存の目(もく)の大部分の祖先が出現した。小型の原始哺乳類も出現した。中生代最後の白亜紀になると,巨大な恐竜類が絶滅し,一方,カモハシ竜,角竜や曲竜など鳥脚類の特殊化が進んだ。それに対し,肉食性恐竜に分化がみられる。海域は拡大し,海生爬虫類ではモササウルスなどが新たに加わった。高緯度地域までソテツ,シュロなど暖帯または亜熱帯性の植物が産出し,海には造礁サンゴが広範に形成されたことから,今日よりはるかに温暖な気候であった。また被子植物も現れた。1億数千万年続いた恐竜の時代は6500万年前で終わった。
恐竜の絶滅の原因については,哺乳類による淘汰説,遺伝機構崩壊説,伝染病説,放射能説,隕石説,生息環境変遷説,種族の寿命説など多くの説がある。白亜紀の最後には恐竜ばかりでなく,陸生・水生・空中を問わず大型動物である爬虫類や頭足類のアンモナイトなど,支配的な動物が全地球的に絶滅した。しかも,そうした現象は白亜紀ばかりでなく,いろいろな時代の異なる動物にもみられる共通した現象である。いくつかの説は普遍的な考えとして受け入れにくい問題がある。大きな外因は,このころ地球上ではアルプス造山運動がはじまり,海陸の分布が変化し,気候も寒冷化への兆しがあった。そしてすでに白亜紀中期から被子植物が増加するなど環境変化が目だってきたことである。一方,内因として,恐竜など動物界では形態・形質の特殊化が進んでおり,新しい環境に適応して生存することが困難であったと考えるのが一般的である。
執筆者:長谷川 善和
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
…アンモナイト類では古生代に栄えたゴニアタイト類が絶滅し,代わって三畳紀を特徴づけるセラタイト類が繁栄するようになった。中生代末にはアンモナイト,恐竜などの絶滅が起こっている。また,二枚貝類ではイノケラムス類,ヒプリテス類が絶滅し,サンカクガイ類が急速に衰退するなどの変革があり,海洋の生態系は大きく変化したと考えられる。…
※「恐竜」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
目次 飼養文化 北アメリカ 北方ユーラシア偶蹄目シカ科の哺乳類。北アメリカでは野生種はカリブーcaribouと呼ばれる。角が雄だけでなく雌にもふつうある。体長130~220cm,尾長7~20cm,...