
デジタル大辞泉 「化石」の意味・読み・例文・類語
か‐せき〔クワ‐〕【化石】
1 地質時代の生物の遺骸が地層中に保存されたもの。巣穴・足跡などの
2 消滅するはずの古いものが、そのまま残っていること。また、そのもの。「封建時代の
3 石になること。石のようになって動かないこと。
「―するまで此処を離れまい」〈露伴・露団々〉
Sponserd by ![]()
翻訳|fossil
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
過去の地質時代における生物(古生物)の遺骸(いがい)や遺体と生活の痕跡(こんせき)(生痕)を化石という。すなわち、古生物の骨、歯、貝殻などの遺物や遺体、それと足跡(あしあと)、這(は)い跡、巣穴などの生活の記録のうち、堆積(たいせき)物の中に埋没したものが、長い地質時代を経たにもかかわらず消滅することなく保存され、のちに掘り出されてきたものが化石である。英語ではfossilというが、これはラテン語のfodere(掘るという動詞)からきたことばで、「掘り出されたもの」という意味である。そのため、化石には本来石のように固いという意味はないが、長い年月、地層の中に埋没している間に、続成作用や地殻運動の影響を受けて変質したために固くなる場合が多い。このような変化を石化作用とよんでいる。つまり、化石はかならずしも固いとは限らず、植物の葉や動物の皮膚や筋肉などの軟らかい組織の化石も知られている。氷河時代の氷漬けのマンモスや、中生代の乾燥気候のもとでミイラと化した恐竜の皮膚の化石や、こはくの中に封じ込められた昆虫の化石などの例もある。また新しい地質年代の地層からは、ほとんど現在の生物と変わらない状態の化石が発見されることがある。このようなものを準化石sub-fossilとか、半化石semi-fossilとよんで区別することがある。人の手が加わって人類遺跡や貝塚から発見されたものは化石とはよばない。
[大森昌衛]
化石ができるためには、普通次のような条件をもつことが望まれる。
(1)化石となる古生物の個体数が多いこと(広い地域に繁栄したこと)。
(2)古生物の遺骸や遺体および生活の記録が消去される前に堆積物の中に埋没すること。
(3)古生物の遺骸や遺体が、続成作用や地殻運動による熱や圧力の影響を受けても、分解、溶解することのない骨・歯・貝殻のような固い組織でできていること。
(4)化石が発掘されて人の目に触れる条件にあること。
化石は、これらの条件を基準にしてその成因を推定する。
[大森昌衛]
人類が最初に化石を発見した記録はさだかでないが、遠く石器時代にまでさかのぼる。クロマニョン人の遺跡から化石の巻き貝でつくった首飾りやターバンが発見されていることが、この事実を物語っている。また、スウェーデンの先史時代の古墳(前10世紀)からは、遠くから運んできたと考えられる古生代や中生代の化石が掘り出されている。しかし、これらはいずれも、偶然の機会に石器時代や先史時代の人間生活に、化石が持ち込まれたにすぎない。
人類が最初に化石について考察した記録は、紀元前7世紀にさかのぼる。古代ギリシアの哲学者クセノファネスやアナクシマンドロスが、貝化石や植物化石、魚類化石を発見して、人間の先祖と結び付けて説明した記録が残っている。しかし、化石の科学的な成因についてはとうてい思い付かなかったが、リディア王国サルディスの哲学者サンタスXanthus(前500ころ)が、内陸から貝化石が産出することに注目して、これらの地方はかつて海底にあったものが陸地に変わったと説明していることが注目される。中世に入っても、化石の科学的説明にはみるべき進歩もなかったが、11世紀のアラビアの科学者イブン・シーナー(アビケンナ)や13世紀のスコラ神学者・生理学者アルベルトゥス・マグヌスが、化石を「大地の造形力によってつくられたが、生命力をもつに至らなかったもの」と記している。このころまでは、化石は自然の戯れの産物か、神秘的な造形力がもたらしたものという考えが支配的であった。化石の成因について最初に科学的な説明を与えたのは、イタリアのレオナルド・ダ・ビンチと、同じく16世紀のイタリアの医師・科学者・詩人であったフラカストロで、「過去の海域に生息した動物の遺骸が乾陸化したために内陸から発見されるようになったもの」と考えた。これより先12世紀に中国の朱子(朱熹(しゅき))が、『朱子全書』(1170ころ)のなかで高山の泥から発見した貝殻の化石について、「この泥は昔は海にたまっていた泥で、貝は海に生息していたものが、低い土地が高くなり(隆起)、柔らかいものが固い石に変わったもの」と説明していることが注目される。一方、16世紀に、ドイツのアグリコラは、化石を鉱物として扱い、石化作用の仕組みについて科学的に説明するきっかけをつくっている。
[大森昌衛]
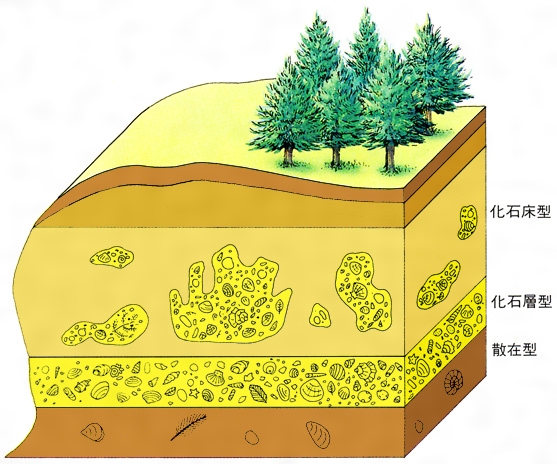
古生物の遺骸、遺体や生活の記録が堆積物の中に埋没して化石として発見されるまでの一連の過程を、化石化作用という。そのため、化石化作用の解明には、古生物の生体や生活に関する研究や、堆積作用の内容に関する研究、続成作用や地殻運動に関する研究などが必要とされる。化石の産状(産出の性状)は、化石化作用の内容によって異なるが、大きく分けて化石床型、化石層型、散在型の三つに分類される。また、化石が古生物の生息時の状態で堆積物の中に埋没したものを自生の化石または現地性の化石といい、化石の埋没状態や地層の特徴から古生物の生息時の姿勢や環境などの古生態を推定することができる。これに対して、古生物の遺骸や遺体が水や大気で運ばれて、生息地域とは異なった地域で堆積物の中に埋没したものを他生の化石または異地性の化石という。このような化石は、化石を包含している地層の研究(堆積学や地層学)によって、古生物の生活域を復原しなければならない。生痕化石のような生活の記録の化石は、化石が発見された場所にその記録を残しているため、化石の形態や産状から直接その古生物の生態や生活環境を推定することができる。1940年にロシアのエフレモフJ. A. Efremovは化石の産状や成因に関する研究をタフォノミーTaphonomyとよび、古生物学の重要な研究分野としている。taphoは埋没を、nomyは学問を意味する。
化石の産状の研究には、堆積学や地層学の研究が並行して行われねばならない。前述の化石の産状の分類も、化石を包含する地層の単位ごとに行われ、地層中における化石の分布と堆積構造との関係や、化石の破損状態や埋没姿勢などを総合して、化石の成因や古生物の生態、生活環境などを推定する。デンマークのステノやイギリスのW・スミスは、化石の産状と地層の関係を研究して地層累重の法則と、動物群類同の法則を発見した。この法則は、化石の内容によって地層の年代を決定できることを明らかにしている。つまり、化石が、地層の年代を決定するための時計としての役割を果たすことが認められたのである。
化石のなかには、地球上の広い地域にわたって特定の地層のみから産出するものがあるため、この化石の発見によって容易に地層の年代を決めることという。また、化石のなかには特定の堆積環境を示すものがある。サンゴの化石は透明な暖海域を、立ち木の化石は陸域を示している。このように環境を指示する化石を示相化石といい、地層堆積時の環境推定に役だっている。
[大森昌衛]
化石化作用の内容によって、化石の性状も異なる。古生物の遺骸、遺体やその一部が保存されたものを体化石(たいかせき)body fossilという。これに対して、生活の記録の化石を生痕化石trace fossil(生活化石または痕跡化石ともいう)といい、排泄(はいせつ)物や卵の化石、胃石なども含まれる。排泄物の化石を糞石(ふんせき)(コプロライト)といい、軟体動物や魚類、恐竜類、哺乳(ほにゅう)類などの糞石が知られている。化石の石化作用の過程で、化石を構成する物質が再結晶したり、外から注入した物質で置き換えられたりすることがある。このような化石を鉱化した化石とよぶ。石化作用によって、もとの化石が砂や泥で置き換えられたものを石核または鋳型の化石(いがたのかせき)という。また、化石の輪郭や彫刻が堆積物の表面に押し付けられたものを、印象化石impression fossilという。つまりクラゲや昆虫および植物の葉などは、本体は分解しても堆積物の表面に外形や彫刻が、生体が分解して残った炭素でくま取られて残ることがある。これが印象化石である。
化石には肉眼で識別できる巨視化石(大形化石ともいう)のほかに、ルーペや、岩石の薄片や研磨面を顕微鏡下で観察して初めてその存在を識別できる微化石microfossilがある。有孔虫、放散虫などの原生動物のほか、珪藻(けいそう)類、花粉、胞子などの植物化石は微化石の例である。また、高倍率の顕微鏡や電子顕微鏡などによって識別できるコッコリスやバクテリアなどの超微化石もある。このほか岩石の化学分析によって識別できる化学化石chemical fossilや、化石とは無縁の偽化石(ぎかせき)pseudofossilがある。
[大森昌衛]
動物化石の記録は、原生代後期にさかのぼるが、末期(約6億年前)には硬組織をもたない後生動物が急激に発展している。
[大森昌衛]
古生代カンブリア紀には、小形の三葉虫や古盃(こはい)動物・軟体動物・腕足動物などの硬組織をもった後生動物が出現している。これらはカナダのバージェス動物群や、中国南部の澄江動物群(ちょうこうどうぶつぐん)Chengjiang faunaで代表されている。また原生代後期に出現しているオーストラリアのエディアカラ動物群の内容となっている後生動物の形態は、現生動物はもとより、バージェス動物群の内容とも著しく異なっている。バージェス動物群の動物がカンブリア紀初期に、骨格や殻などの硬組織をもつようになった過程や要因については、いまなお明らかでないが、ロシア台地や中国南部などのカンブリア紀初期の地層から硬い殻をもった小有殻動物群small shell fossils(SSFと略称)が発見されており、硬組織をもった最初の動物群として注目されている。
[大森昌衛]
(1)原生動物 原生動物の出現は原生代後期にさかのぼる。原生動物門は普通、単細胞で、肉質虫類(アメーバ、有孔虫、太陽虫、放散虫など)と鞭毛虫(べんもうちゅう)類、胞子虫類、繊毛虫類の四つの亜門に大別されるが、化石として重要なのは肉質虫類である。とくに、有孔虫類には古生代後期の標準化石となっている紡錘虫(ぼうすいちゅう)類(フズリナ)や、中生代・新生代の標準化石となっている大形有孔虫(高等有孔虫)が含まれる。また、小形有孔虫や放散虫は珪藻(けいそう)化石とともに石油の原物質となっており、これらの遺骸(いがい)や遺体が大量に堆積(たいせき)して生物岩をつくっている場合が多い。エディアカラ動物群のなかには、複数の核をもつ肉質虫類の仲間が大形化した特殊の動物があると考えられている。
(2)海綿動物 骨片の化石は原生代後期の地層から産出しているが、体制を残したアティコカニアAtikokaniaがカンブリア紀の地層から報告されている。日本では北上高地の古生層や白亜系(石灰海綿)、各地の第三系(珪質海綿)から発見されている。
(3)腔腸(こうちょう)動物 この化石の記録も原生代後期にさかのぼるが、古生代以降の海域に発達し標準化石となっているものが多い。しばしば暖海域に群生し礁を形成するため、地質時代の石灰岩のなかには、化石サンゴ礁を構成し、古環境や古生態復原の貴重な研究対象となっているものもある。古生代には床板(しょうばん)サンゴ類や四放サンゴ類が、中生代には六放サンゴ類が発達している。またヒドロ虫類の仲間の層孔虫類には、古生代・中生代の標準化石となっているものが多い。造礁性サンゴは100メートル以浅の透明な温暖海域に発達するため、造礁性サンゴのつくる石灰岩の分布によって、古気候や古環境を復原した研究例が多い。ヒドロ虫やクラゲ、ウミエラの仲間の化石はエディアカラ動物群から報告されている。
(4)扁形(へんけい)動物・紐形(ひもがた)動物・円形動物・袋形(たいけい)動物・環形動物 これらの仲間は、キチン質のほか硬組織を欠くため化石としては保存されにくい。わずかに、古生代後期や古第三紀の昆虫化石に寄生している扁形動物や、魚類の化石に付着している円形動物が発見されているにすぎない。これらの動物の体化石の例はきわめて少ないが、這(は)い跡、掘り跡、排泄(はいせつ)物などの生痕化石(生活化石)の記録が多い。エディアカラ動物群に相当する中国の化石群からは、ミミズの仲間の貧毛類やゴカイの仲間の多毛類の化石のほか、紐形動物と思われる生痕化石が産出している。
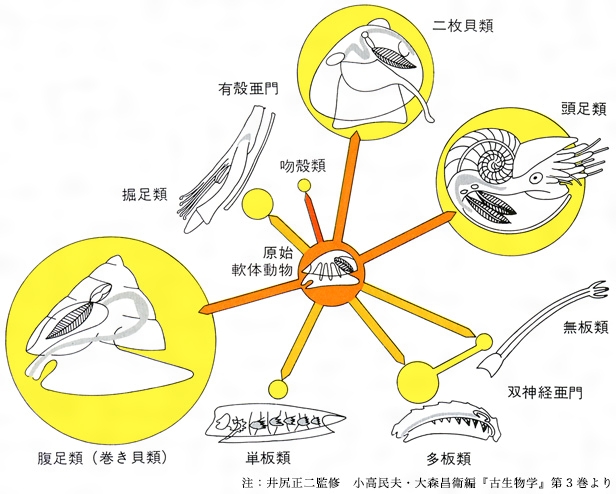
(5)軟体動物 古生代カンブリア紀以降のあらゆる地質年代に生息し、殻(から)をもったきわめて多様な種に分化発達しているため、化石の記録がもっとも多い仲間となっている。軟体動物は、殻をもたない双神経亜門と殻をもつ有殻亜門に2大別され、体制の差によって多くの綱に細分されている。
双神経亜門の動物体には、石灰質の骨片が発達し、まれに骨片の化石が発見される。多板類(ヒザラガイ)は、8枚の石灰質の殻板をもち、古生代オルドビス紀以降から化石が知られている。殻板の接合部の彫刻が、古生代、中生代、新生代の順に複雑になっている。単板類は、古生代のカンブリア紀からデボン紀にかけて産出する。口縁が楕円(だえん)形で、全体が丸い笠(かさ)形をした1枚の殻で、殻の内面には5、6対の筋痕(きんこん)が残っている。1952年に太平洋の深海からこの仲間の遺存種であるネオピリナNeopilinaが発見されてから、この動物の体制や構造が明らかになり、原始体節動物から軟体動物が進化してきた系統分類上の位置が定められた。ロシアの白海沿岸にあるエディアカラ動物群に相当する化石群からは、キンベレラkimberellaとよばれる原始的な単板類の化石が報告されている。
腹足類(巻き貝類)は軟体動物のなかでももっとも繁栄した仲間で、化石の種類ももっとも多い。この仲間は殻体のほかに、軟体を殻体に納めたときに殻口を保護する蓋(ふた)と、口腔(こうこう)内に消化を助ける数個ないし数百個の微小な小歯群からなる歯舌(しぜつ)がある。歯舌の形は、腹足類の系統分類の重要な目安となっている。殻の形態や歯舌の型によって、古腹足類、中腹足類、新腹足類の三つの目(もく)に分類され、それぞれが古生代のカンブリア紀、オルドビス紀、白亜紀に出現し、進化の度合いもこの順に高くなっている。この仲間には、いろいろの地質時代の標準化石となっているものも多く、また殻の形態や殻表彫刻が地質年代の順に変化していくみごとな生系列をつくるものも多く知られている。
二枚貝類(斧足(おのあし)類ともいう)は、巻き貝類に次いで化石の種類が多い仲間である。古生代後期以降に繁栄し、標準化石となっているものも多い。とくに、中生代の海域に発達したサンカクガイ(トリゴニア)、イノセラムス、厚歯(あつば)二枚貝などが注目される。
掘足類(ツノガイの仲間)は、オルドビス紀の海に出現し、石炭紀以降に属や種の数が増しているが、化石の産出例に乏しく、殻の形態変化も少ないため、地層の序列や年代決定および進化の研究対象としてはあまり重要なものとは考えられていない。
古生代前期の海には、左右の双殻をもちながら前後に開口した空隙(くうげき)の後方に足を伸ばして、ツノガイに似た生活型をもっていたと考えられている仲間の化石が報告されている。この仲間を吻殻(ふんかく)類Class Rostroconchaとして独立させ、二枚貝類と掘足類の中間型に位置させている研究者もある。
頭足類は、箭石(やいし)類やアンモナイト(菊石)類など化石によって地層の序列や年代決定、および進化についての研究の重要な資料となっているものが多い。また、この仲間の子孫には直角石の遺存種として知られているオウムガイや、イカ、タコなどの仲間があるため、これらの生理、生態、組織、解剖などの生物学的研究によって、化石種の復原に関する興味ある研究が進められている。
(6)節足動物 この仲間は動物界では著しい発達を遂げたグループで、原生代後期の海域に出現し、古生代には三葉虫類、剣尾類(カブトガニの仲間)、広翼類(ウミサソリの仲間)、ウミグモ類などが発達して、重要な標準化石となっている。古生代前期末には陸域に進出し、唇脚(しんきゃく)類(ゲジなど)、倍脚類(ヤスデなど)、結合類(コムカデなど)や各種の昆虫類が出現している。昆虫類は現在著しい発達を遂げ、人間生活ともきわめて関係深い動物となっている。この仲間の化石は、石炭層や湖成層と、これらに伴って産出するこはく中に包埋された完全な標本が発見されている。
カンブリア紀の海には、奇妙な姿の原始的な節足動物の多様な種が出現し、短い期間に姿を消している。なかでも小形の貝虫類や、ウミサソリに近い貝甲類などの化石は、地層の序列や年代決定に重要な情報を提供している。軟甲類(エビ、カニの仲間)も古生代中期の海に出現しているが、化石として重要なものは中生代ジュラ紀以降に報告されている。
(7)コケムシ動物(蘚虫(せんちゅう)または蘚苔虫(せんたいちゅう)ともいう) これらの仲間は、1ミリメートル前後の個虫が集まって群体をつくり、水中ではほとんどが付着生活をしている。オルドビス紀に発達し、古生代末期に姿を消した胞孔目、変口目、隠口目の仲間には、古生層の序列や年代決定の研究上重要な化石種となっているものが多い。
(8)腕足動物 これらの仲間は現在の動物界ではきわめて小さい位置を占めているにすぎないが、地質時代には世界各地の海域に発達し重要な化石となっている。原生代末に出現し、古生代には著しく発達したため、殻が大形化し左右に翼状に伸長したもの(スピリファ)や、殻表に長い棘(とげ)などの彫刻をもつもの(エキノコンカスなど)がみられる。これらは中生代、新生代を通じ絶滅の道をたどっているが、現在の海に住むシャミセンガイやホオズキガイの仲間は、腕足動物の遺存種である。
(9)棘皮(きょくひ)動物 ウミユリ、ウニ、ヒトデ、ナマコの仲間も先カンブリア時代末に出現し、カンブリア紀の海に発達している。
(10)触手動物・箒虫(ほうきむし)動物・有鬚(ゆうしゅ)動物・毛顎(もうがく)動物 前口動物と後口動物の中間的特徴をもつこれらの仲間の化石の記録は乏しく、わずかにカナダのロッキー山脈のカンブリア紀中期のバージェス頁岩(けつがん)から毛顎動物の化石が発見されているにすぎない。このほか古生代後期の地層から箒虫動物が掘った住み穴の化石(生痕化石)と思われるものが報告されている。
[大森昌衛]
原索動物(げんさくどうぶつ)の化石として、古生代前期の標準化石となっているものに筆石(ふでいし)類Graptolithinaまたは筆石動物Graptozoaがある。キチン質の硬質部をもち剣盤から始まる群体を形成する。カンブリア紀中期から石炭紀前期までの海底に固着し、または海藻その他に付着して浮遊し、ときに浮袋をもつものもある。しばしば黒色頁岩中に産出する。
カンブリア紀の5億4000万年~5億3000万年前のバージェス動物群や澄江動物群からは、脊椎動物の起源を示すフサカツギ(羽鰓(うさい)類)やナメクジウオ(頭索動物)などの原索動物の仲間の化石が発見されている。
とくにカンブリア紀以降の地層から産出するコノドント(錘歯(すいし)類)は2ミリメートル以下の微小化石で、どの動物の器官であるかが決められないまま、軟体動物の歯舌とか、下等魚類の歯、軟骨魚類の楯鱗(じゅんりん)などと考えられていたが、1970年にアメリカの石炭紀の石灰岩から産出した、ナメクジウオに似た魚類化石の胃の中に、コノドントの群体が発見されたため、原脊椎動物の濾過(ろか)摂食器官の一部と考えられている。一方、1974年にイギリスのモリスR. Morrisは、バージェス動物群から産出したコノドントを歯としているアノマロカリスAnomarocalisやその仲間と考えられるオドントグリフスOdontogriphusとよぶ化石を研究して、触手動物の仲間と考えた。
[大森昌衛]
脊椎動物の化石には、骨や歯のほかに角(つの)、爪(つめ)などが知られている。魚類の場合には体表を覆う各種の鱗(うろこ)や、海生魚類の平衡器官である耳石(じせき)や、コイ・フナなどの淡水生魚類の咽頭歯(いんとうし)のほか、排泄(はいせつ)物や卵の化石も報告されている。
(1)魚類 最古の魚類は、原索動物から進化した無顎類(むがくるい)(ヤツメウナギ)の仲間とされている。カンブリア紀初期のナメクジウオのような甲皮に覆われない魚形の祖先から、著しく特殊化した甲皮に覆われた魚類として進化したものである。この仲間はデボン紀前期にかけて発達していくが、デボン紀末には姿を消している。一方、シルル紀に出現した有顎魚類は、板皮類として発達したが、デボン紀末には甲皮類とともに姿を消している。
軟骨魚類の最古の化石はデボン紀中期から知られているが、これ以前に板皮類のあるグループから由来したものと考えられている。この仲間の多くは肉食性で、中生代後期に硬骨魚類の真骨類が出現するまでは、軟体動物や甲殻類などを食べて、当時の海洋における生態的地位の頂点を占めていた。
デボン紀に出現した硬骨魚類は、空気呼吸のための肺と陸上歩行のための四肢を発達させて、両生類を経て爬虫(はちゅう)類へ進化していった。硬骨魚類は両生類を分化した内鼻孔魚類と、現在の海洋に発達している条鰭(じょうき)類の二つのグループに分けられている。
(2)両生類 この種の最古の化石は、グリーンランド東部のデボン紀後期の地層から発見されたイクチオステガIchthyostegaである。現生の両生類は、有尾類(イモリ、サンショウウオなど)、無尾類(カエルなど)、無足類(アシナシイモリなど)の三つのグループに分類されているが、有尾類は三畳紀、無尾類はジュラ紀、無足類は第三紀になって、それぞれ最初の化石が知られている。
(3)爬虫類(はちゅうるい) 石炭紀中期に両生類から漸進的に進化し、古生代末期から中生代にかけて著しい発達を示している。この仲間は、角質化した皮膚や石灰質の硬い殻に包まれる有羊膜卵をもつことで両生類と区別される。そのため、爬虫類は骨格や歯・角・爪の化石のほか、卵殻や糞(ふん)の化石、また乾燥気候のもとでミイラ化した皮膚の化石が発見されている。また、この仲間は、胃の中に小石を飲み込んで消化を助けているため、骨格化石に伴って胃の中の石が発見されることがある。これを胃石Gastrolithという。
卵殻の化石や胃石は、体化石に伴って発見された場合には動物の所属を決めることができるが、単独で発見されることが多いため、所属を定めるのが困難な場合が多い。最古の卵殻の化石は、アメリカ合衆国テキサス州のペルム紀(二畳紀)前期の赤色岩層から発見されているが、動物の種類は明らかではない。
恐竜類の足跡(あしあと)の化石も報告されている例は多いが、体化石に伴わない場合は動物種を決めにくい。しかし足跡の形態や配列、方位などによって、二足恐竜、四足恐竜のいずれであるかを知ることや、歩行の姿勢、状態、移動の方位などを定めたり、歩行時の環境や年齢、性別、体格などを推定することができる。排泄物から、狩猟動物の行動を推定しているし、野生動物の保護係は、排泄物を健康状態を知るための手掛りとしている。そのため、この種の研究を足痕学pedalogyとか糞形学coprologyとよんでいる。足痕化石や糞石によって古生物の生活や行動を推定する研究を古足痕学paleopedalogyとか古糞形学paleocoprologyとよび、古生痕学(または化石生痕学paleoichnology)の重要な一分野となっている。
恐竜という名称は初め俗語であったが、1970年代以降は恐竜類をまとめて、鳥類や翼竜類を含めて恐竜亜綱とし、学名として扱うようになった。しかし意見は定まっていない。1980年代以降は、恐竜の骨の化石が哺乳類の骨の化石と同じ特徴をもつことなどから、恐竜類も温血性であったという意見があり、温血動物説と冷血動物説とに分かれている。
(4)鳥類 ドイツのジュラ系から発見された始祖鳥の化石によって、鳥類の出現の時期が考えられてきたが、1990年代になって中国の三畳紀後期の地層から、羽毛をもった翼竜類の化石が発見され、鳥類の起源はさかのぼるものと考えられている。近代的な鳥類の化石は、白亜紀から発見された陸鳥と水鳥の仲間の化石である。
鳥類の化石の地質時代における発見数から考えて、白亜紀後期と、新生代に2回の発展の時期があったもので、とくに第三紀中期以降に著しく多様化している。
(5)哺乳(ほにゅう)類 哺乳類は温血性であることと、歯が分化し胎盤が発達したことによって、爬虫類と区別される。
爬虫類は多数の同形の歯をもち、一生に何度でも生え換わる(多生交換)。これに対して、哺乳類は、切歯、犬歯、臼歯(きゅうし)などに分化しており、一生に一度交換(二生性)するだけである。爬虫類の歯のこのような分化は、古生代のペルム紀から中生代の三畳紀にかけて南半球に存在したゴンドワナ大陸に生息した獣弓類Therapsidaの仲間に始まっている。この仲間の体形も哺乳類に似ているものが多い。
中生代の陸域を爬虫類が支配していたため、小形の原始哺乳類(原獣類)の出現をみたので近代型哺乳類は発達しなかった。現生のカモノハシ(単孔類)は、爬虫類的形態を残し鳥類のように卵を孵化(ふか)し哺乳を行う。この起源については意見が分かれているが、多丘歯類(ジュラ紀末から新生代初期にかけて生存した、齧歯(げっし)類に似た歯をもつ小形の原獣類)に似ているという説もある。ジュラ紀の原獣類には三錐歯(さんすいし)類や梁歯(りょうし)類などの化石がよく知られている。
中生代に出現した、現生哺乳類に関係ある化石哺乳類には、汎(はん)獣類、後(こう)獣類がある。汎獣類は相称歯類と真汎獣類とに二分され、後者が正獣類の根幹をなすグループと考えられている。いずれも白亜紀末には姿を消している。
後獣類には、オーストラリア、ニューギニアなどに現存する有袋類が含まれる。有袋類の化石は北アメリカの白亜紀にさかのぼり、南北両アメリカの新生代第三紀以降の地層から化石が発見されている。オーストラリアにおける有袋類の化石の記録は、中新世後期または漸新世後期にまでさかのぼる。有袋類の起源はジュラ紀の汎獣類にさかのぼり、正獣類とは隔離されて発展したため、生態学上や形態学上は正獣類と同様の適応放散を示している。
正獣類の最古の化石は、モンゴル国(外蒙古(もうこ))の白亜紀後期の地層より発見された食虫類の仲間デルタテリディウムDelthatheridiumとされているが、中国東北地区から産出したジュラ紀後期のエンドテリウムEndotheriumを食虫類とする考えもある。中生代後期に出現した食虫類を起源として、新生代に入ると正獣類は急激に発達し、多くの種類に分化している。
爬虫類から哺乳類へ進化する過程については、化石の資料に乏しいため、学者の間での意見が分かれている。イギリスの動物学者ハクスリーは、爬虫類的特徴を多く残している哺乳類を下等動物と考え、単孔類・有袋類・正獣類の順に進化したとする単系統進化を主張している。これに対し、イギリスの人類学者ブルームらは、単孔類を有袋類、正獣類(真獣類)と区別し、二つの進化系列が存在するという多系統説を提唱している。また、アメリカの古生物学者シンプソンらは、爬虫類時代にすでに哺乳類に通ずる多くの系列に分かれていたと考えている。
哺乳類の化石には新生代の重要な標準化石となっているものが多く、とくに奇蹄(きてい)目(ウマ、サイなど)、偶蹄目(シカなど)、長鼻目(ゾウなど)、霊長目などの系統進化については詳細な研究が行われている。
[大森昌衛]
貝殻や骨の化石のなかには、明らかに奇形を示しているものや、内部組織が破壊されたり、寄生生物によって変形、変質しているものが知られている。このような病気、奇形の化石を対象として、古生物の発生、組織、生理学上の異常を研究する分野を古病理学paleopathology(または化石病理学)とよんでいる。貝殻の奇形・病形の化石のほか、脊椎動物の骨格化石の組織が骨髄炎などによって破壊されたものが報告されている。巨視化石の場合には、巻き貝に穴をあけられた二枚貝や、魚竜・首長竜などの歯の跡が残ったアンモナイトやウニの化石、肉食恐竜に尾をかまれたアパトサウルスの化石なども発見されて、古動物界における捕食―被食の関係を知るための貴重な資料となっている。これらはすべて、生痕化石(生活化石)に含めて扱われている。
動物界における食物連鎖の関係は、次の三つの段階を経て飛躍的に発達したことが考えられる。第一は、原生代後期~古生代前期における後生動物の発達期であって、この時期には、沿岸域におけるプランクトン、海藻類、後生動物の間の捕食―被食関係の出現による、複雑な食物連鎖の関係が成立したと考えられる。第二は、古生代前期末における陸生動物の出現によって、陸域における食物連鎖が発展し始めた時期である。第三は、ヒトの出現した時期で、道具の使用による人間の生産活動によって自然界の食物連鎖の頂点にヒトがたつと同時に、人為的な営力が加えられ始めたことである。
[大森昌衛]
しばしば系統分類上で二つ以上の分類群に関係し、所属の決められないものが化石として発見されている。このようなもののなかには、中間型の動物種として、系統発生の新しい系列の起源を示すものが多い。始祖鳥はこの例である。また現生の生物界には知られていない化石生物として、新しい分類群として扱われる。コノドントはこの例で、長い間系統分類上の位置不明のコノドント動物門として扱われてきたが、現在では原索動物の摂食器官の化石とする意見が多い。
系統上の位置が不明で、かつ化石としての判定に疑問のあるものを「問題のある化石」(プロブレマティカProblematica)として扱うことがある。また生物起源であることがわかっても正体不明の微小な化石を、アクリタークAcritarchsとよんでいるが、これは原生代の地層から多く発見され、古生代からも報告されている。アクリタークとはギリシア語のacritos(「不明」または「混乱した」の意)とarche(「起源」の意)の合成語である。
[大森昌衛]
植物の微化石には、花粉・胞子、植物性有孔虫などの殻、車軸藻(しゃじくも)類の生卵器などがある。また、イネ科植物などの葉につくられた珪酸体のような、分解耐性があり一定の形状をとる生物起源物質などもある。微化石は、生産量が多く、地層中の時間的・空間的分布も広いので、環境変化や時代決定、地理分布の検証などの指標にしやすい。形態を研究するためには、化石を含む資料から酸などの化学処理によって周囲の岩石や鉱物を除去し、光学顕微鏡や電子顕微鏡で観察するのが一般的である。
植物の大形化石には、植物体の部分や性状、保存状態によってさまざまなものがある。植物細胞には、細胞膜の外側にセルロースを含む細胞壁があり、表皮細胞にはクチクラ、維管束の木部細胞にはリグニンが沈着する。これらの化学物質は分解耐性が高いので、化石化したときの植物組織の保存性は、動物組織に比べて非常に勝っている。
一般に、保存状態により印象化石、圧縮化石、植物遺体、鉱化化石という区別がある。
印象化石は、いわゆる「押し葉」のような化石で、植物体の組織は失われているが、外形や葉脈などの形態を知ることができる。
圧縮化石は印象化石よりも圧縮と変性の程度が低いもので、植物の組織は炭化して失われているものの、表皮のクチクラなどは残っており、表皮細胞や気孔の形状などがわかる。生殖器官からは花粉や胞子が取り出せることもある。
植物遺体はさらに圧縮の程度が低く、植物体を立体的な形状として取り出せるものである。保存の程度は、炭化した黒色のものから、炭化程度の低い褐色のもの、まだ緑色をしているものまで、さまざまである。
鉱化化石は、植物体が鉱物質によって徐々に置換され、細胞や組織が立体的に保存されたもので、鉱物質の「漬け物」といえる。このため、立体的な外観だけでなく、切片をつくることで解剖学的な研究が可能である。分類に役だつ内部構造の観察だけでなく、保存のよい化石では、2億5000万年前の受精過程すら知ることができる。また、組織内に菌類や昆虫など他の生物が保存されていることもある。
植物化石の研究法は、化石情報を取り出す目的と、可能な技術によって異なる。現在では、さまざまな波長を用いた顕微鏡や、X線スキャナ、化学分析機器も開発改良と利用が進んでいる。さらに、地球科学や生物学全般の進歩に呼応した研究目的の展開と、それに沿った化石情報の収集と利用も進んでいる。しかし、化石情報がもっとも利用されるのは、生物の系統進化過程の復元である。以下に、植物化石から明らかにされてきた、初期の藻類と植物の系統について概説する。
[西田治文]
酸素発生型光合成は、地球の生物生産と物質循環において重要な役割を果たしている。酸素発生型光合成を行うもっとも原始的な生物は、原核生物のシアノバクテリアcyanobacteria(藍藻(らんそう)類、藍藻植物)である。この仲間の化石記録は、始生代にさかのぼる。現生のシアノバクテリアが、その遺体と炭酸塩鉱物とともに形成するストロマトライトと同様の構造物が、35億年前の地層から報告されている。また、オーストラリア西北部や、南アフリカの35億~20億年前の地層からは細胞壁のある球状や連鎖状の原核生物の化石が発見されている。水中に増加した酸素により酸化された鉄が堆積(たいせき)したとされる縞(しま)状鉄鉱層は、世界各地の原生代前期の地層中に認められ、堆積性の鉄鉱資源となっている。
真核細胞生物の出現は約20億年前にさかのぼると考えられている。その証拠として、中国の19~17億年前の地層から、真核藻類とされる大型の単細胞生物の化石がみつかっている。オーストラリア中部の約9億年前のビタースプリングス層からは、単細胞緑藻類に似た化石が報告されているが、これらの類縁については議論が続いている。
[西田治文]
生物界を動物と植物とに分類する二界説でとらえると、植物には細菌や菌類、藻(そう)類も含まれる。一方、五界説をとると、植物は陸上に生活する緑色植物、具体的にはコケ植物、シダ植物、種子植物をさし、これらはまとめて陸上植物と俗称される。現在の植物界において、多様性が高く、人類との関わりがもっとも大きいのも陸上植物である。
陸上植物の祖先は、緑藻類である。最初の陸上植物は、現在の車軸藻類に近い淡水の緑藻類が、陸上生活に適応することで分化したものと考えられている。車軸藻類の化石記録は、シルル紀以降にみられる。
陸上の環境は水中と異なり過酷である。強い紫外線、乾燥、重力などは、植物が上陸するにあたっての大きな障害となる。そのため陸上植物には、このような障害を克服する独特の組織や器官が発達した。表皮、通道組織、乾燥耐性のある胞子などである。このような組織の破片が、最初に化石としてみつかるのは、オルドビス紀で、はっきりした植物体の化石はみつからないものの、このころから植物は陸上に進出を始めたと考えられている。
最初の陸上植物とされるのは、シルル紀のクックソニアCooksoniaで、約4億2000万年前のものである。体には現在の陸上植物に一般的な、根・茎・葉という体制の分化がみられず、針金のような軸が立体的に二叉(にさ)分枝を繰り返す単純な姿である。軸の先端には、胞子嚢(のう)が1個ずつつく。シルル紀からは、同じような姿のステガノテカSteganotheca、親指ほどの軸に数センチメートルもある細い葉が密生し、現在のヒカゲノカズラ類の祖先とされるバラガナチアBaragwanathiaも知られている。
これらの初期陸上植物は、すべて胞子嚢をつける胞子体で、しかも現在のコケ植物の胞子体のように、小形で配偶体に寄生するものではない。このため、コケ植物というよりは、現在のシダ植物に近いといえるが、デボン紀の植物化石を詳細に検討した研究からは、この時代にはコケ植物でもシダ植物でもない陸上植物が存在した可能性も指摘されている。
現生植物間の形態や遺伝子比較からは、コケ植物からシダ植物が派生したと結論できる。コケ植物のような体制をもった最古の化石は、デボン紀前期のスポロゴニテスSporogonitesであるが、陸上植物の初期分化は今後も新たな化石の発見により明らかにされるはずである。
[西田治文]
デボン紀の植物は、クックソニアに似た単純な体制を引き継いでいるが、徐々に多様化が進んでいる。特筆されるべきことは、通道組織の構造、胞子嚢の位置と形態、植物体の外見などに違いが生じてきたことである。デボン紀の植物化石産地としてもっともよく知られているスコットランドのライニーから産出するライニー植物群Rhynie floraにおいても、この多様化のあとがうかがえる。
ライニー植物群から最初に記載されたのは、リニアRhynia(2種)、ホルネオフィトンHorneophyton、アステロキシロンAsteroxylonである。このうちアステロキシロンは、葉脈を1本しかもたない針のような葉をもち、胞子嚢は軸の先端ではなく、それぞれの葉の付け根上側につく。この特徴は、シダ植物の小葉類と共通する。シルル紀のバラガナチアも含め、小葉類がかなり早い時期に分化したことがわかる。同じデボン紀のゾステロフィルムZosterophyllum、ドレパノフィクスDrepanophycusも小葉類またはその祖先とされる。
リニア種とホルネオフィトンは互いによく似るが、通道組織に分化がみられる。リニアの1種は現在の維管束植物と同じ型の仮道管をもつのに対し、ほかの1種(後にアグラオフィトンAgraophytonと改称された)とホルネオフィトンはコケにみられる簡単な通道組織しかもたない。シルル紀のクックソニアやステガノテカもこの型である。デボン紀初期までに維管束植物とそれ以外の陸上植物がはっきり分化した。小葉類は維管束植物である。
プシロフィトンPsilophyton、トリメロフィトンTrimerophyton、ペルティカPerticaは、維管束をもつが、小葉類と異なり、胞子嚢が軸の先端につく。さらにリニアと異なり、胞子嚢が集合する傾向がある。また、立体二叉分枝が主体であった体制から、軸に強弱ができ、側軸には分枝した軸が扁平(へんぺい)に集まる傾向がみられる。この扁平な軸の集合が癒合して葉が形成されたと考えられ、小葉類がもつ単脈針状の小葉に対して、大葉とよぶ。維管束植物には少なくとも系統の異なる2つの型の葉が生じた。
デボン紀中期のイビカIbykaでは、植物体の側軸が等間隔に集合する傾向がみられる。イビカは等間隔の節を特徴とし、シダ植物の楔葉(けつよう)類(トクサ類)の祖先といわれる。分子系統解析の結果も含め、維管束植物は単系統であることがわかっているが、その初期分化はデボン紀におこったといえる()。
[西田治文]
デボン紀は陸上の生物相が急激に豊かになった時代である。その理由の一つが、森林の出現である。森林をつくる樹木は、二次成長して樹幹を形成する植物で、最初はシダ植物に出現した。デボン紀には小葉類、トクサ類と、絶滅群の前裸子植物が、並行して二次成長する能力を獲得した。前裸子植物は、針葉樹のような樹幹をもちながら、胞子繁殖するシダ植物で、アルカエオプテリスArchaeopterisはその代表である。最初の森林はデボン紀後期につくられ、地上に空間構造が生まれて動物の多様化も促進され、土壌も豊かになり始めた。脊椎(せきつい)動物もこのころ上陸した。
前裸子植物のなかから、種子で繁殖する種子植物が現れた。最古の種子植物は、3億7000万年前のエルキンシアElkinsiaとモレスネティアMoresnetiaである。現在、種子植物には種子が裸である裸子植物と、雌しべの子房内に種子ができる被子植物とがあるが、最初の種子植物は裸子植物であった。初期の裸子植物は、種子がシダのような葉にそれぞれ離れて単独に生じ、針葉樹類の球果(まつかさ)のようにまとまった生殖器官を形成しなかった。このような段階の裸子植物を、シダ種子類という。シダ種子類は石炭紀に多様化し、中生代以降の裸子植物の祖先となった。代表的なものに、メドゥロサMedullosa、リギノプテリスLyginopterisがある。
温暖な石炭紀には裸子植物よりもシダ植物が繁栄し、膨大な量の石炭が形成された。小葉類の鱗木(りんぼく)類、シギラリアSigillaria(封印木(ふういんぼく))、トクサ類のカラミテスCalamites(ロボク)がよく知られている。石炭紀後半からペルム紀になると寒冷化と乾燥が進み、シダ植物が衰退するかわりに、種子植物が台頭を始めた。
[西田治文]
裸子植物は中生代に全盛となり、現生の4群(イチョウ、ソテツ、球果、グネツム類)もペルム紀から三畳紀にかけて出現したとされる。さらに、カイトニアCaytonia、キカデオイデアCycadeoidea(ベネチテス)類など、多くの絶滅群が中生代末期まで相次いで出現した。そのなかで共通してみられたことは生殖器官の特殊化であり、分類群ごとに独特の形態と構造をもった生殖器官が進化した。とくに、種子がさまざまな方法で包まれ始めた。しかし、のちの被子植物のように、種子のもとである胚珠が雌しべに包まれた結果、花粉が胚珠に直接到達できなくなったものはなかった。
被子植物の祖先と起源の地は、現在も特定されていない。花粉化石からは、約1億4000万年前までに被子植物が赤道に近い低緯度地域に出現したことがわかっている。被子植物の大形化石でもっとも古いものは、中国産白亜紀前期のアルカエフルクトゥスArchaefructusである。
被子植物は、白亜紀の間に徐々に分布と量を拡大し、北アメリカの白亜紀末には植生の半分近くか、所によりそれ以上を占めていたことがわかっている。現在のような熱帯雨林は、温暖であった新生代古第三紀始新世ごろに成立した。さらに漸新世から中新世にかけておこった地球規模の寒冷化により、中緯度地域に草原が発達するなど、現在の植生の基本がつくられた。このような寒冷化の過程と気温低下の度合いは、樹木の葉の化石の形態解析からも推定されている()。
[西田治文]
化石の採集に先だって、化石の産状や化石を包含する地層の性状について、詳細な観察記録を残すことを怠ってはならない。とくに化石の産状のスケッチや写真撮影も必要である。化石の採集には、母岩の固さや化石の埋没状態によって、たがねやドリルなどの補助用具を用いて、化石の破損を避けなければならない。破損した場合には、接着剤を用いて現地で修復しておくか、破損部位を明確にして、持ち帰ってからの復原に必要な表示を行ったり、記録を残しておくように心がける。
母岩から剖出できないような化石や微化石を採集する場合は、母岩を採集して持ち帰ってから室内で化石の抽出処理を行う。化石を運搬する場合には、破損を防ぐため綿や新聞紙などで包んだり、標本を石膏(せっこう)で固めたり、材木を組んだ箱のなかに吊るすなどの手法が用いられる。とくに、肉眼では識別できない花粉、胞子、有孔虫、コノドントなどの微化石を母岩から抽出する場合には、化石の存在が期待できる岩片を岩石截断(せつだん)機や回転粉砕機などによって細かく砕いたり、希塩酸やフッ酸などにつけて母岩を軟化させた後に、特殊な試薬を用いた比重選別法などによって、微化石を母岩から分離抽出する。巨視化石の抽出のためには、万力などの岩石截断機や、ビブラトゥールとよぶ小形の電気ドリルを用いるとよい。また、破損しやすい巨視化石の抽出は、衝撃を与えることのないよう、砂袋の上で小形のたがねや、のみ、錐(きり)などを用いる。
化石の産地から標本を運搬する際は、産地や産状についての記録と照合できるような記号を標本につけたのち、運搬の途中で破損することのないように、前述のように緩衝物を用いるなどの配慮が必要である。
持ち帰った化石の整理には、以上述べたような母岩からの剖出のほかに、化石の洗浄、破損部位の接着、軟弱標本の硬化などの処置が必要である。母岩から剖出できない微化石や、巨視化石の内部組織を研究するためには、母岩の表面を研磨して反射顕微鏡で調べたり、母岩の薄片について岩石顕微鏡で観察することも行われる。化石の洗浄には、化石の硬度に応じてワイヤブラシ、針、各種の刷毛(はけ)などが使われる。弱塩酸や酢酸などを用いて化石の表面を洗浄したり、化石の微細構造の起伏を浮き出させたものを合成樹脂のフィルムなどに写し取ってレプリカ標本をつくることもある。また、石膏やプラスチック、モデリング・コンパウンドなどで化石の型をつくることもある。このほか、化学分析、アミノ酸分析、X線回折などのために化石の粉末を用いる。化石標本の保存のためには、標本採集地、採集年月日、地層名などの必要事項を記入したラベルをかならず添付しておくことがたいせつである。
[大森昌衛]
『益富壽之助・浜田隆士著『原色化石図鑑』(1966・保育社)』▽『浅野清著『古生物学入門』(1968・朝倉書店)』▽『大森昌衛編著『地史・古生物』(1969・共立出版)』▽『A・L・マックアレスター著、大森昌衛訳『地球生物学入門――生命の歴史』(1970・共立出版)』▽『鹿間時夫著『増訂版 日本化石図譜』(1970・朝倉書店)』▽『D・L・アイッカー著、大森昌衛訳『地史学入門――地質時計』(1971・共立出版)』▽『浅野清著『古環境論――古生物学の立場より』(1971・朝倉書店)』▽『井尻正二著『古生物学汎論』全3巻(1972・築地書館)』▽『大森昌衛監修『地層と化石』(1977・東海大学出版会)』▽『森下晶編『日本標準化石図譜』(1977・朝倉書店)』▽『B・コックスほか著、大森昌衛訳『古生物の世界――生命の発生から人類の誕生まで』(1980・文理)』▽『西村三郎著『地球の海と生命』(1981・海鳴社)』▽『マーチン・J・S・ルドウィック著、大森昌衛・高安克己訳『化石の意味――古生物学史挿話』(1981・海鳴社)』▽『藤山家徳ほか編『学生版 日本古生物図鑑』(1982・北隆館)』▽『秋山雅彦ほか著『双書地球の歴史』全7巻(1984~1985・共立出版)』▽『L・B・ホールステッド著、亀井節夫訳『太古の世界を探る――化石、地層、生痕、生命の起源と進化』(1985・東京書籍)』▽『日本古生物学会編『化石の科学』(1987・朝倉書店)』▽『デビッド・ランバート編、長谷川善和・真鍋真訳『図説化石の百科――343種の謎を解く』(1988・平凡社)』▽『日本化石集編集委員会編『日本化石集』68集・別1集(1988~1989・築地書館)』▽『日本古生物学会編『古生物学事典』(1991・朝倉書店)』▽『S・M・スタンレー著、長谷川善和・清水長訳『生物と大絶滅』(1991・東京化学同人)』▽『井尻正二・秋山雅彦編著『化石の世界』(1992・大月書店)』▽『リチャード・G・ブロムリー著、大森昌衛監訳『生痕化石――生痕の生物学と化石の成因』(1993・東海大学出版会)』▽『地学団体研究会編『地学教育講座6 化石と生物進化』(1995・東海大学出版会)』▽『間島隆一・池谷仙之著『古生物学入門』(1996・朝倉書店)』▽『速水格・森啓ほか編『古生物の科学シリーズ』全3巻(1998~2001・朝倉書店)』▽『大森昌衛著『進化の大爆発――動物のルーツを探る』(2000・新日本出版社)』▽『化石研究会編『化石の研究法――採集から最新の解析法まで』(2000・共立出版)』▽『福田芳生著『生痕化石の世界――古生物の行動を探る』(2000・川島書店)』▽『ジョヴァンニ・ピンナ著、小畠郁生監訳、二上政夫訳『図解 世界の化石大百科』(2000・河出書房新社)』▽『リチャード・フォーティ著、渡辺政隆訳『生命40億年全史』(2003・草思社)』▽『井尻正二著『化石』(岩波新書)』▽『J・ウィリアム・ショップ著、阿部克巳訳『失われた化石記録』(講談社現代新書)』▽『浅間一男著『生物はなぜ進化したか』(講談社ブルーバックス)』▽『B・クルテン著、小原秀雄・浦本昌紀訳『哺乳類の時代』(1976・平凡社)』▽『浅野清ほか編『新版 古生物学 第1、2、4巻』(1978・朝倉書店)』▽『井尻正二監修『古生物学各論 第2~4巻』(1981・築地書館)』▽『E・H・コルバート著、田隅本生監訳『脊椎動物の進化』(1994・築地書館)』▽『福田芳生著『古生態図集・海の無脊椎動物』(1996・川島書店)』▽『岩槻邦男・馬渡峻輔監修、白山義久編『バイオディバーシティ・シリーズ5 無脊椎動物の多様性と系統(節足動物を除く)』(2000・裳華房)』▽『スティーヴン・ジェイ・グールド著『ワンダフル・ライフ――バージェス頁岩と生物進化の物語』(ハヤカワ文庫)』▽『西田誠著『たねの生いたち』(1972・岩波書店)』▽『浅野清ほか編『新版 古生物学 第3巻』(1978・朝倉書店)』▽『井尻正二監修『古生物学各論 第1巻』(1981・築地書館)』▽『遠藤隆次著『植物化石図譜』(1986・朝倉書店)』▽『L・E・グラハム著、渡辺信・堀輝三訳『陸上植物の起源――緑藻から緑色植物へ』(1996・内田老鶴圃)』▽『岩槻邦男・馬渡峻輔監修、加藤雅啓編『バイオディバーシティ・シリーズ2 植物の多様性と系統』(1997・裳華房)』▽『西田治文著『植物のたどってきた道』(1998・日本放送出版協会)』▽『岩槻邦男・加藤雅啓編『多様性の植物学2 植物の系統』(2000・東京大学出版会)』▽『浅間一男・木村達明著『植物の進化』(講談社ブルーバックス)』▽『西田誠著『陸上植物の起源と進化』(岩波新書)』
化石の産状
軟体動物の系統分類
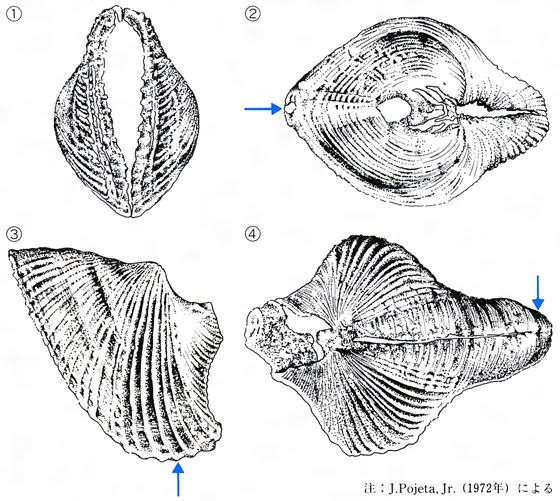
吻殻類の化石

毛顎動物(アミスキウィア)
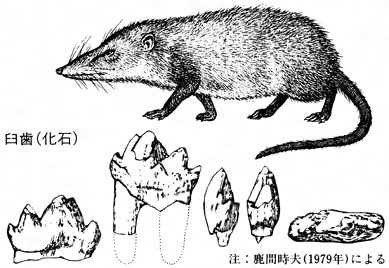
モルガニュコドン(梁歯類)の復原図と臼…

エンドテリウムの復原図と臼歯

シュードフズリナ(紡錘虫)

シリンゴポーラ(床板サンゴ類)

イノセラムス・シュミッティ(二枚貝)

ベレムナイト・ハスタータ(箭石)

ゴウドリセラス・デンセプリカタム(アン…

オドントケファルス・アエゲリア(三葉虫…

プテリゴツス・マクロフタルムス(ウミサ…

エンクリヌス・リリフォルミス(ウミユリ…

ボトリオレピス・カナデンシス(板皮類)

ダペディウム・フォリドツム(硬骨魚)

ホメオサウルス・マキシミリアニ(爬虫類…

アーケオプテリックス・リトグラフィカ(…

初期の陸上植物〔図A〕
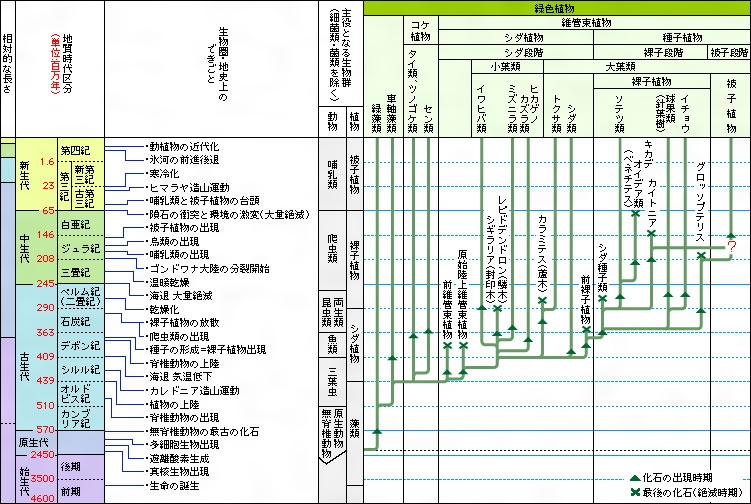
陸上植物の系統〔図B〕

レプトフロエウム・ロンビクム(ヒカゲノ…

カラミテス(トクサ類)

ギンゴイテス(イチョウ類)

ポドザミテス(球果類)

オトザミテス(ベネチテス類)

キカデオイデア・エゾアナ(ベネチテス類…
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
地質時代の生物(古生物)の遺体や生活の痕跡が地層中に埋没・保存されたもの。文字どおりの意味では〈石に化したもの〉であるが,化石は必ずしも石化していない。英語のfossilはラテン語で〈掘り出された〉を意味するfossilisに由来している。19世紀初期までは,鉱物の結晶のようなものまで化石に含められていたが,今日では比喩的に用いられる以外は,自然状態で地層中に埋没している古生物の遺物に限定されている。
古生物の遺体の化石(体化石)は保存状態によって3種類に分けられる。すなわち,体部の構造や組織がそのまま全部ないしは一部残存しているもの,他の物質によって置換されたもの,および岩石・鉱物中に痕跡のみを残したものである。体部がそのまま残存している場合,まれに軟組織もそっくり化石として残ることがある。その種の化石には,乾燥した砂漠や洞穴中で脱水しミイラ化した爬虫類や哺乳類,北シベリアの凍土中に冷凍されたマンモスやケサイ(毛犀),ポーランドのオイルサンドに埋もれたサイのような大型動物をはじめ,樹脂の化石であるコハク中に閉じ込められた昆虫やクモ類,岩塩の結晶中のバクテリアのような微小なものまで,多くはないがさまざまな例がある。しかし大多数の化石は古生物の死後多かれ少なかれ変化したものである。動物の骨格・歯・貝殻あるいは植物の樹幹のような,機械的破壊や化学的分解作用に対する抵抗力に富む硬質部のみが残っているのが普通である。植物の遺体は樹幹や葉のほか,球果・種子・花粉など繁殖器官も,セルロースなど分解しにくい化学組成であるため残りやすい。生物の硬組織を構成する無機成分は,主としてアラレ石および方解石(CaCO3),リン灰石(Ca5[PO4]3[F,Cl,OH])ないしオパール(SiO2・nH2O)であるが,まれに天青石(SrSO4),ストロンチアナイト(SrCO3),磁鉄鉱(Fe3O4),ファーテライト(CaCO3)のものがある。炭酸塩鉱物の場合は溶解しやすく,方解石・ドロマイト・石英などによって置換される。オパールで構成されている場合は石英に変わり,また方解石で置換されることもまれではない。古生物遺体の置換は前述のような鉱物によるもののほか,岩石中の酸化鉄に由来する鉄化合物によるものがあり,黄鉄鉱化・褐鉄鉱化した貝殻や骨格の化石が少なくない。古生物の体部を構成する物質がこのように置換されることもなく,完全に消失してしまって,その型のみが岩石・鉱物中に残されることがある。この残された痕跡を印象化石という。古生物の体部の外表面の型は外形雌型(めがた),内表面のものは内形雌型である。岩石中に残されたこれらの型の間に堆積物が侵入したり,地下水が浸透して鉱物が沈殿し,元来の体部とそっくり同じ形の型が形成される。このような実物の表面と凹凸の一致した型を雄型(おがた)と呼ぶ。雄型は研究のため人工的によく作られる。内形雄型は二枚貝などの場合,対になった貝殻の間に粘土を入れてはさめば容易に得られるが,自然状態でも各種の岩石や鉱物により形成されている。これを核ないし石核(ドイツ語ではSteinkern)という。
古生物の生活活動の痕跡が岩石や鉱物の形で残存しているものは生痕化石と呼ばれる。生痕化石はいろいろの基準によって分類される。例えば成因的には,(1)海藻やバクテリアなどが堆積物の粒子を方解石やケイ酸で固結させて作り上げた層状堆積構造(ストロマトライト),採餌行動によりかくはんされた状態がそのままとどめられた堆積構造,軟らかい堆積物の表面に残された足跡・はい跡・巣穴・採餌行動の跡など生物起源の堆積構造,(2)固結した堆積物に対して穴をあけた跡,かじったり,かんだり,ひっかいたりした生物による侵食構造,(3)その他の生活活動を示す糞(ふん)・卵・卵囊・胃内容物などに分類される。
形態が小さく,顕微鏡下ではじめて識別されるような化石を微化石microfossilという。この中には,有孔虫・放散虫・貝形虫・ケイ藻のような微小な殻や骨格をもつ動植物,複雑な構造をもつ大型動物の骨格の一部分(例えばウニのとげや骨片など)や,植物の花粉・胞子などの化石が含まれる。さらに超微化石などと呼ばれているのは,500倍ないし1000倍の高倍率の光学顕微鏡下でようやく識別できる種類の古生物の遺骸であり,代表的なものには石灰質ナンノプランクトン(コッコリス)がある。これらの殻の詳細な構造は電子顕微鏡によって究明される。さらに,岩石中に含まれる生物起源の有機化石物などは化学化石と呼ばれる分子レベルの化石である。そのほか,擬化石と呼ばれるものがある。地層の生成過程やその後の熱・圧力・地下水などによる続成作用によって,古生物とはかかわりなく化石のような見かけの形態を生じたもので,字義どおり化石ではない。
化石は古生物として過去の生物圏の一員であると同時に,現在は岩石圏を構成する物質の一つである。この間の転化の全過程を対象とする古生物学の分野を化石生成論taphonomyという。ここでは古生物の死因,遺体の軟組織の分解,生息地外の場への埋没,埋没に先立つ硬組織の機械的破壊,選択的溶解,堆積物中における配置様式,遺体により生じた堆積物表面の痕跡,化石の続成過程,石核形成過程,化石の変形,続成過程における選択的溶解,分子の再配列・再結晶,地層中での団塊形成,化石の交代作用(一つの鉱物から他の化学組成をもつ鉱物に漸進的に置換される作用),硬組織の保存などが扱われる。これら諸過程のうち,化石の続成過程以後のものは化石続成論,それ以前のものは化石産出論の対象として二分される。以上のような化石化の過程に関与する要因は多数あるが,成因的にはなかでも,その古生物が繁栄して個体数が多かったという生活時の条件,死後遺体が堆積物中に埋没するまでに受けた諸作用に耐えうる強い硬組織や構造を備えていたという埋没時までの条件,そして埋没後引き続いての続成作用・変質作用・地殻変動などによる溶解や分解を免れたなどの保存条件が重視されている。
化石化の過程で起こる古生物の原組織の変化では,溶脱・蒸留・付鉱化・鉱化(石化)などが多く見られる。溶脱は遺体を構成する物質が溶解して消失する作用であって,キチン質の表皮や石灰質の硬組織の場合によく起こる。蒸留はキチン質の外骨格のようなものの分解過程で,大部分の窒素・酸素・水素が蒸発し炭素のみが薄膜となり残存する作用である。筆石類・昆虫類・植物などに多い。付鉱化は多孔質透水性の貝殻や骨格に炭酸カルシウムやケイ酸のような無機物質が付加する作用である。多くの場合,地下水が孔隙(こうげき)に浸透して物質が沈着し,引き続いてそれらが晶出して硬組織に加わり,強化し,硬組織を破壊から守ることになる。鉱化ないし石化は,地下水が完全に硬組織を溶解し,代りに他の物質を沈殿する作用であって,この場合は古生物の原組織が残されない。こうした置換でできた化石を元来の硬組織の仮像(かぞう)pseudomorphという。
地層中に見られる化石の状態は,古生物の生態(古生態)や化石・地層の成因を探る重要な手がかりとなる。地層のうちで化石を多産する部分を一般に化石層と呼んでいるが,化石の産状には散在型と密集型とがある。密集部の産状を記述するには,形態や成因を考慮して種々の用語が用いられている。密集して層をなし水平方向にかなり連続性をもつものを狭義の化石層という。これに対して,地層内で化石を多産する部分が含まない部分をはさみ断続的に横に並ぶのは化石床(かせきしよう)と呼ばれる。化石床の場合は,周囲の無化石の部分と共通の層理面をもっていて,両者が同時的に形成されたことを示すというその成因が重視されている。また形態や化石内容に基づいたものにバイオハームbiohermとバイオストロームbiostromeがある。バイオハームは塚状,ドーム状,レンズ状または礁状の岩塊で,ほとんどサンゴ,ウミユリ,有孔虫,軟体動物,層孔虫のような固着性生物群の石灰質の遺骸で構成されており,周囲を違った岩質の堆積岩で取り囲まれているものを指す。一方,バイオストロームは,明りょうな成層構造をもち広く広がっている毛布状ないし薄いレンズの岩体であって,大部分貝殻,ウミユリ,サンゴのような固着性生物の遺骸で構成されているけれども,前者のように周囲からとくに盛り上がった形状は示さない。
散在型の産状を示すものには,泥質の堆積岩中に生活時の体位を保ったままで埋もれている二枚貝のような例が少なくない。また樹幹や樹根化石が直立したまま地層に埋もれている化石林もその一例であるが,このような生活地と埋没地(堆積地)とが一致しているものを現地性ないしは自生的化石という。生痕化石はその多くが現地性であるため,古環境を知るのに重視されている。このような古環境を指示する化石を示相化石という。これと反対に,遺骸が水や大気によって生活地外へ運搬され堆積したものを異地性ないしは他生的化石という。水流により掃き寄せられ,多くの破片とともに密集状態で産出する貝化石層はその好例である。異地性化石の場合,遺骸が生活地より堆積地までの間,長距離を運搬されても,現地性化石と同時期のもの(同時的異地性化石)とみなされる。それに対して,いったん化石となったものが,その後の地殻変動などにより地層中より洗い出され,また再び別の地層中に埋没されたもの(異時的異地性化石)は誘導化石derived fossilあるいは二次化石,再堆積化石などと呼ばれる。
化石の採集にあたって必要なのは,採集用の道具,採集地に関する知識,および効果的な採集手段についての経験である。初心者にとって最も手っとり早いのは,各地において学校・博物館などを中心に組織されている地学同好会などに参加し,野外巡検を行うことであろう。古生物学・地質学の入門書もかなり助けになる。
採集用具として最も基本的なものはハンマーであるが,このほか化石を岩石から取り出すためのたがね,化石産地や地質図を記入するための地形図,観察事項を記録する野帳(フィールドノートあるいは方眼紙をつづったノートブック),筆記用具,マジックインク,消しゴム,ものさし,ルーペ,古新聞紙などの包装用紙,採集袋(ポリエチレンやビニル製など),およびリュックサックなどが最低限必要である。次にどこで化石を探すべきか,過去に採集されたことのある地点については地域地質に関する巡検案内書,地質図,あるいは学術論文などの出版物からかなり知識を得ることができる。また,それぞれの産地付近の人たちから多くの具体的情報を得られることが多い。しかし,さらに新産地を系統的に探すには堆積岩の地質調査法に関する基礎的知識が必要である。近年は土地の開発,道路の新設など土木工事が活発に行われ,新産地が増加する一方で,都市周辺の古くからの産地が消失していることが多く,さらに自然保護のため有名産地が天然記念物指定を受けて採集禁止になっていることも少なくないので,事前調査が望ましい。
化石を含むような露頭に向かった場合,まず露頭基部に風化してたまっている岩くずの中に落ちている化石の有無を調べ,また風化面を綿密に観察して化石の痕跡を探すのが能率的である。その後,はじめてハンマーを用いて露頭の岩石をたたき割り化石を探せばよい。採集にあたり地形図に採集地点を入れ,野帳に露頭のスケッチと化石の産状や地質に関する観察事項を記入することが必要で,できれば露頭の遠景・近景の写真撮影をしておく。産地や産出地層についての表示のない化石は,科学的研究にとり無価値に等しい。大型の無脊椎動物化石の場合,泥質岩や固結の進んでいない砂岩に含まれているものはそのまま取り出せることが多く,紙に包み,採集地点名を記し,採集袋におさめて持ち帰る。小型のものはさらにパッキング材で包むなど注意を要する。硬い岩石中から取り出すにはハンマーとたがね,つるはしなどが用いられる。殻が溶脱して外形雌型のみ残っている場合には,型取り材料で雄型模型を現地で作るなり,あるいはそのまま持ち帰ればよい。微化石の場合,野外においてできるだけルーペで存在を確認してから,それを含む岩石を採集することが望ましいが,判定にはかなり経験を必要とする。石灰質の殻の化石の場合は殻の溶脱が少なくないので,とくに新鮮な岩石の採集に努めねばならない。植物化石は砂岩・泥質岩・夾炭(きようたん)層・細粒凝灰岩などに伴われることが多い。葉の化石などは地層の層理面に沿ってよく見いだされ,また炭化物になっていることもある。採集にはハンマー,たがね,つるはし,スコップ,ナイフなどを用い,堆積岩を割って化石を出すなり,大きく堆積岩を切り出して持ち帰る。脊椎動物の化石では,歯や骨の部分化石などがよく産出するが,しかし1個体分の骨格が密集して産することもまれではない。そのため一部を発見してもそれでとどめず,周辺の十分な調査を行う必要がある。発掘にあたっては,専門研究者の指導の下に計画的に進めなければならない。
採集して持ち帰った化石は整理後,化石の種類に応じて処理をし,不必要な母岩を取り除き,破損部を補修し,標本として種の同定を行えるようにする。このような準備段階を整形ないし剖出という。大型化石の整形には,各種のたがね,小型ハンマー,きり,金のこ,ピンセット,はけなどのほか,万力,超音波洗浄装置,歯科用デンタルマシンのような小型の機器が用いられる。化石の内部構造を明らかにするには,化石の一部を破壊・除去し,また切断して薄片を作成するほか,軟X線による透視なども行う。雌型や雄型しか残っていない化石の場合は,同定のため原形を復元する必要があり,模型を作成する。これには型取り材料として,油粘土,モデリングコンパウンド,セッコウなどが使用される。微化石の場合は,岩石やまだ固結の進んでいない堆積物より化石を分離するか,化石を含む岩石の薄片を作るか,あるいはその岩石の研磨面を作る。分離には単純な水洗以外に各種の薬品による処理が行われるが,化石や岩石の種類により異なった方法を用いる。分離した化石は,微化石用スライドに接着するか,スライドグラスに封入し標本とする。以上のような各種の化石標本の観察・記録・計測は,直接肉眼による以外に,各種の光学顕微鏡・電子顕微鏡や軟X線発生装置による観察・写真撮影により行われる。このような化石の研究法については,種々の参考書が市販されている。
化石を同定(または鑑定)して,その生物分類上の位置を決める,すなわち種名を決めるには,広い知識と多くの文献が必要である。一応の見当をつけるには,動植物や化石の図鑑類が役だつ。しかし,さらに突き詰めるには,その化石の時代や分類上のおおよその位置をつかんだうえで,専門研究書や学術論文にあたって,記載内容をよく調べなければならない。そのうえで種名を決定するには,その種のみならず近縁種を含めた大きい分類群(科・属など)についての知識のほか,分類学上の基礎知識ももち,かつ国際動(植)物命名規約にも通じている必要がある。古生物の分類には,できるかぎり完全かつ多数の標本に基づく形態の計測が原則として必要で,その他数量分類学・発生学・生化学的手法も研究に用いられる。以上のような一般的手続については,入手の容易な市販の手引書などに解説されており,また各地の自然史関係の博物館や大学などの地学系教育・研究機関で助言が得られよう。
化石の研究の中で困難な作業の一つは古生物の復元である。復元には,分解して地層中に散乱する化石骨から古生物の全体像を組み立てたり,類似の生物が現存しない〈謎の化石〉から古生物の姿をよみがえらせたり,さらには化石群から古生物群集の生態を明らかにすることまで含まれる。例えば,大型動物化石の場合,発掘作業が復元の初段階であって,現場周辺の地質調査を行って産出層準を確認したうえで,地層中の化石の埋没位置・姿勢などを詳細に記録しつつ発掘を進め,不要な母岩をできるだけ除去し,くずれやすい部分の一時的補強を行って掘り上げる。各部分は番号つきで持ち帰られるが,実験室では最初に道具や薬品により整形・補強を行う。また必要に応じ,各部の複製標本を作る。その後,骨格の再構成を行うが,化石化過程での変形を修正し,失われた部分を補うには,現生動物の骨格や既存の化石の完全に保存されている部分を比較参照しなければならない。最後に,発掘時の産状を念頭におき,部分を計測しつつ分類・整理して,配列し,保持用の支柱や心材を用いて組み立てて固定する。完全な復元のためには,さらに肉付け標本を作るが,これには筋肉のつき方など現生動物を参考に彫塑用材料・樹脂材などを用いて行う。国外では恐竜類をはじめ多数の例があるが,日本でも北海道広尾郡忠類村産のナウマンゾウはその一例である。また骨格復元標本としては,デスモスチルスやオオツノシカが知られている。古生態の復元例としては,大規模発掘によって知られる長野県の野尻湖底から発見されたナウマンゾウが著名である。
地球上に生命が誕生してから30億年以上も経過し,その間に地球表層部に蓄積された化石は莫大な数に達する。これらの化石が記載・同定され,古生物の分類体系が編纂(へんさん)された結果,過去から現在にまたがる動・植物界の門・綱・目・科・属・種などの分類上の階級的位置が定められ,系統進化の筋道が明らかにされ,種の分化・進化速度・絶滅の原因などが追究されてきた。最近ではコンピューターによる統計処理により記録の定量的解析も進められている。かつて地球上に生息した古生物の種数は,研究者により推定値に幅があるが,1億ないし5億とされている。ちなみに現生種の推定数は110万ないし450万である。また,種の平均寿命は50万年から1500万年とされている。先カンブリア時代の最古の化石とみなされているバクテリアや藻類状の原核生物の記録は,34億年前にさかのぼることができる。その後約20億年前に真核生物が出現し,約7億ないし5.9億年前ころになると多細胞の動物の生痕化石がオーストラリア,アフリカ,ヨーロッパ,シベリアの各地から報告されている。これらはまったく殻を欠いていたか,あるいは少なくとも体部の鉱物化が行われなかったのである。約5.9億年前に始まる古生代初頭のカンブリア紀には,無脊椎動物のかなりの数の門が出現したが,それらはリン酸カルシウムに富んだキチン質や方解石の殻を備えていた。今日知られている化石種の99%はこの時代以後のものである。このことは,化石の記録が保存されやすい殻や骨格をもった生物の出現で多分に左右されていることを示している。しかし,また一面では,カンブリア紀の古動物群の多様化は,海進により食物が豊富で環境的に安定した浅海域が地球上に広がったことに関連するであろう。以後,オルドビス紀における最初の脊椎動物である魚類,シルル紀における陸上維管束植物,石炭紀における爬虫類などの出現があって,約2.48億年前に始まった中生代に移り,さらに6500万年前に新生代に入って動植物はいよいよ多様化し発展しつつ現在に至っている。この間に多くのグループが盛衰を繰り返したが,昆虫類は石炭紀以来,鳥類はジュラ紀以来,被子植物は中期白亜紀以来,哺乳類は新生代以来というように,それぞれ緩急の相違はあるがしだいに隆盛化し,現在絶頂期に達している。これに反して,腕足類,頭足類,ウミユリ類などは古生代や中生代に最盛期を過ごして今や衰退しつつある。また三葉虫類,筆石類,アンモナイト類,恐竜類のように絶滅しているものも少なくない(絶滅生物)。
化石の記録はまた,地層の年代を決定し,あるいは離れた地域間の地層の時間的対応関係を明らかにする対比の手段として重要視される。生物進化や環境変化の結果,個々の化石ないし化石群の産出は地質時代の特定の時間および空間に限られる。累重する地層中での化石の層位的分布を究め,化石内容に基づいて地層を化石帯biozoneに区分し,対比するのが生層位学(化石層位学あるいは生層序学ともいう)である。化石帯はいろいろの基準により定義されるが,化石の組合せや特定化石の層位的産出範囲(古生物の生存期間)に基づくものが多い。このように,それが含まれている地層を認定し,年代を決定するのに用いられる化石を示準化石という。
化石化した古生物とそれが生息していた環境との関係を扱う古生物学の分野が古生態学である。そこでは,地層中の化石や化石群の形態的特徴・産状・量的組成・種構成などによって,古生物の生活様式や生息場所が明らかにされる一方,また堆積岩自体も検討され化石を含む地層が堆積したときの古環境が復元される。化石の形態からは古生物の生理・生態に関して多くの情報がもたらされる。生物の形態には生活様式や環境に対する適応があるため,化石種と近縁の現生生物の体制・構造などとの比較によって,古生物の実態が明らかにされる。また現生生物との比較が不可能な場合でも,化石の形態から直接にそれらのもつ機能的意義が探られる。さらに殻や骨格の形成に関与する物理化学的過程も追究され,海生生物の石灰質の殻の酸素同位体比から形成時の水温が測定されたり,化石骨に残る膠原(こうげん)質のアミノ酸分析によって化石哺乳類の体温測定が行われている。一方,化石の埋没している地層も重要な情報源である。地球表層部に広がる多様な環境では,物理・化学・生物・地理さらに地質的条件により,それぞれに特有の堆積岩が形成される。それらの組成や構造に,堆積時の大気や水の状態はもちろん,気候・地形条件が反映している。堆積岩中の希元素類も有用で,ホウ素は塩分量を測る相対的尺度として用いられる。堆積物の粒度組成は直接にはその場の運動エネルギーを反映しているが,また底生生物にとり栄養や生活様式と深い関係があって,現地性・異地性化石を問わず古生態の解明のため重要である。
化石群の構成や産状は,自然状態における古生物群集を知る直接の手がかりである。しかし,化石の群集は生活している生物群集と同一ではない。両者の成因や起源上の相違を明らかにするのに種々の用語が用いられる。例えば遺骸群集thanotocoenosisであるが,これには死後運搬・集積された遺骸の集まりを意味する場合と,単一の生物群集の遺骸に異地性の遺骸が混合した群集を指す場合がある。古生物の死後の過程を明らかにするには,現生生物の死後の諸過程に関する知識が不可欠であり,このためドイツで古生態学の1分野として現在古生物学が体系化された。
微化石の場合,本質的には大型化石と異ならないが,産状の観察が容易でない反面,少量の岩石試料中に多数含まれているので,群集の定量的解析を行いやすい。そのため油田開発や海洋底探査には,堆積岩の試錐試料による年代決定や古環境解析にもっぱら微化石が用いられる。底生動物の微化石による海底環境の解析と,海洋プランクトンの微化石による水温・塩分・酸素および炭素の安定同位体比など海水の諸要素の測定が行われ,全地球的な古海洋変動・古気候変動の解明が進んでいる。1.8万年前の世界の気候図を復元したアメリカの気候長期研究・図化・予測計画(CLIMAP。1976)の基礎資料は,これら微化石の多変量解析から得られた。また,深海掘削計画(DSDP)で海洋プレートの年代を決定し,古海洋の変遷を明らかにするのに微化石の研究が貢献している。
化石は地球表層部の堆積岩の構成員の一つとして大きな役割を果たしている。現在の熱帯・亜熱帯海に分布するサンゴ礁が礁性生物の遺骸で構成され,また深海底では浮遊性有孔虫の遺骸が降り積もって軟泥を形成しているように,古生物の遺骸がそのまま堆積して岩石化した石灰岩,ドロマイト,チョーク,ケイ藻土,あるいは古生物遺骸から変化した石炭・石油・天然ガス,海鳥の糞が堆積・固化してできるグアノを起源とするリン鉱床,鉄バクテリアにより生成された鉄鉱床など,人間生活にとって有用な地下資源となっているものが少なくない。かつては化石は正倉院薬物中に見られる竜歯・竜骨・竜角のたぐい(実は哺乳類の化石)のように,洋の東西を問わず薬物として大いに珍重されていた。また,コハクのほか,放散虫の遺体で形成されることの多い碧玉(へきぎよく)など,昔から装飾や宝石に使われている化石もある。
執筆者:高柳 洋吉
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
fossil ,organic remains
過去の生物の遺体または遺跡が地層中に埋没・保存されたもの。化石という語は石化(petrification)に由来するが,石化は化石の必要条件ではない。化石の多くは生物の遺体がそのまま,あるいは鉱物質に置換されて,または印象として残ったもので,これらは遺体化石(remain, Überrest, Krperfossil)と総称される。古生物の遺跡(生活の産物)が残ったものが生痕化石である。古生物に由来するが,変質して形態的・構造的特徴を残していないもの(例;石油・石炭・リン鉱・褐鉄鉱など)は,ふつう化石から除外する。また化石(fossil)という語は,古生物とは無関係な無機的事象に対して,「過去の」を意味する言葉として転用される場合(例;化石景観,漣痕化石,地磁気の化石など)があるが正しい用法ではない。
執筆者:絈野 義夫
出典 平凡社「最新 地学事典」最新 地学事典について 情報
Sponserd by ![]()
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
…古生物を研究する科学。生物を扱う点では生物学の一分野であるが,化石を直接対象として地質時代の生物現象を研究する点では地球の歴史科学である。化石に人間が関心を抱き始めた時代を特定することはできないが,少なくともクロマニョン人の遺跡から貝化石で作った首飾が出土していることで,その古さがわかる。…
…堆積岩は量としては地殻表層の20%以下を占めるにすぎないが,表面積としては実に75%以上に達し,しかも地層が地球の歴史に関する三次元的枠組みを与えることが明らかになったことから,重要なものとして扱われている。
[地層研究史]
地層には化石が含まれているが,その意味については長い間正確には理解されなかった。ギリシア時代にアリストテレスは化石を観察し,それらは海の生物の遺骸であり,かつて陸地は海に覆われていたと考えた。…
※「化石」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
夏の暑さに体が慣れること。数日から数十日間で起こる短期暑熱順化と、数年または数世代にかけて起こる長期暑熱順化とがある。→寒冷順化[補説]近年では、冷房設備の普及にともない短期暑熱順化が起こりにくくなっ...