デジタル大辞泉 「光合成」の意味・読み・例文・類語
こう‐ごうせい〔クワウガフセイ〕【光合成】
[類語]反応・化学反応・連鎖反応・化学変化・化成・化合・合成・腐食・漂白・分解・加水分解・感光・解毒・核融合・核分裂・核爆発
Sponserd by ![]()
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
光(ひかり)合成ともいう。植物が光のエネルギーを利用して二酸化炭素CO2と水H2Oから有機化合物を合成する過程。その反応は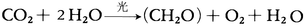 炭酸固定の代表的な例で,より一般的には,光のエネルギーを利用してCO2を還元する過程をいう。ファン・ニールvan NielはCO2+2H2A─→(CH2O)+2A+H2Oを光合成の一般式として提唱している(1929)。光合成細菌(緑色硫黄細菌,紅色硫黄細菌などの硫黄細菌,紅色無硫黄細菌)は,水素供与体として水ではなくH2S,H2S2O3,H2,有機化合物などを用いる。反応はCO2+2H2S─→(CH2O)+2S+H2O,CO2+2H2─→(CH2O)+H2O,この場合はO2の発生はない。細菌型光合成を基礎として進化の結果,地球上に豊富に存在する水を水素供与体とする緑色植物型の光合成が約10億年前に出現し,地球上にO2の出現をもたらした。
炭酸固定の代表的な例で,より一般的には,光のエネルギーを利用してCO2を還元する過程をいう。ファン・ニールvan NielはCO2+2H2A─→(CH2O)+2A+H2Oを光合成の一般式として提唱している(1929)。光合成細菌(緑色硫黄細菌,紅色硫黄細菌などの硫黄細菌,紅色無硫黄細菌)は,水素供与体として水ではなくH2S,H2S2O3,H2,有機化合物などを用いる。反応はCO2+2H2S─→(CH2O)+2S+H2O,CO2+2H2─→(CH2O)+H2O,この場合はO2の発生はない。細菌型光合成を基礎として進化の結果,地球上に豊富に存在する水を水素供与体とする緑色植物型の光合成が約10億年前に出現し,地球上にO2の出現をもたらした。
1個の光量子によって,固定されるCO2または発生するO2の分子数を光合成の量子収量quantum yield of photosynthesisといい,この逆数を光合成の要求量子数quantum requirement of photosynthesisという。要求量子数として,かつてO.H.ワールブルクは4という値(4光量子説)を主張したが,現在では8~10という値が一般に受け入れられている。波長678nmの光(赤色光)を用いた場合,1モル光量子のエネルギーは41kcalであるから,6CO2+6H2O+672kcal─→C6H12O6+6O2よりCO21モルの還元に112kcalの自由エネルギーが必要となり,要求量子数8ではエネルギー効率は34%である。自然状態では,植物によって吸収された太陽光(赤外域を除く)のうち,光合成によって化学エネルギーとして蓄えられるのは1%程度で,1年間に地球上で光合成により固定される炭素量は約2×1011tと推定され,陸上植物による固定量と水中の藻類による固定量の比は約1:9といわれる。
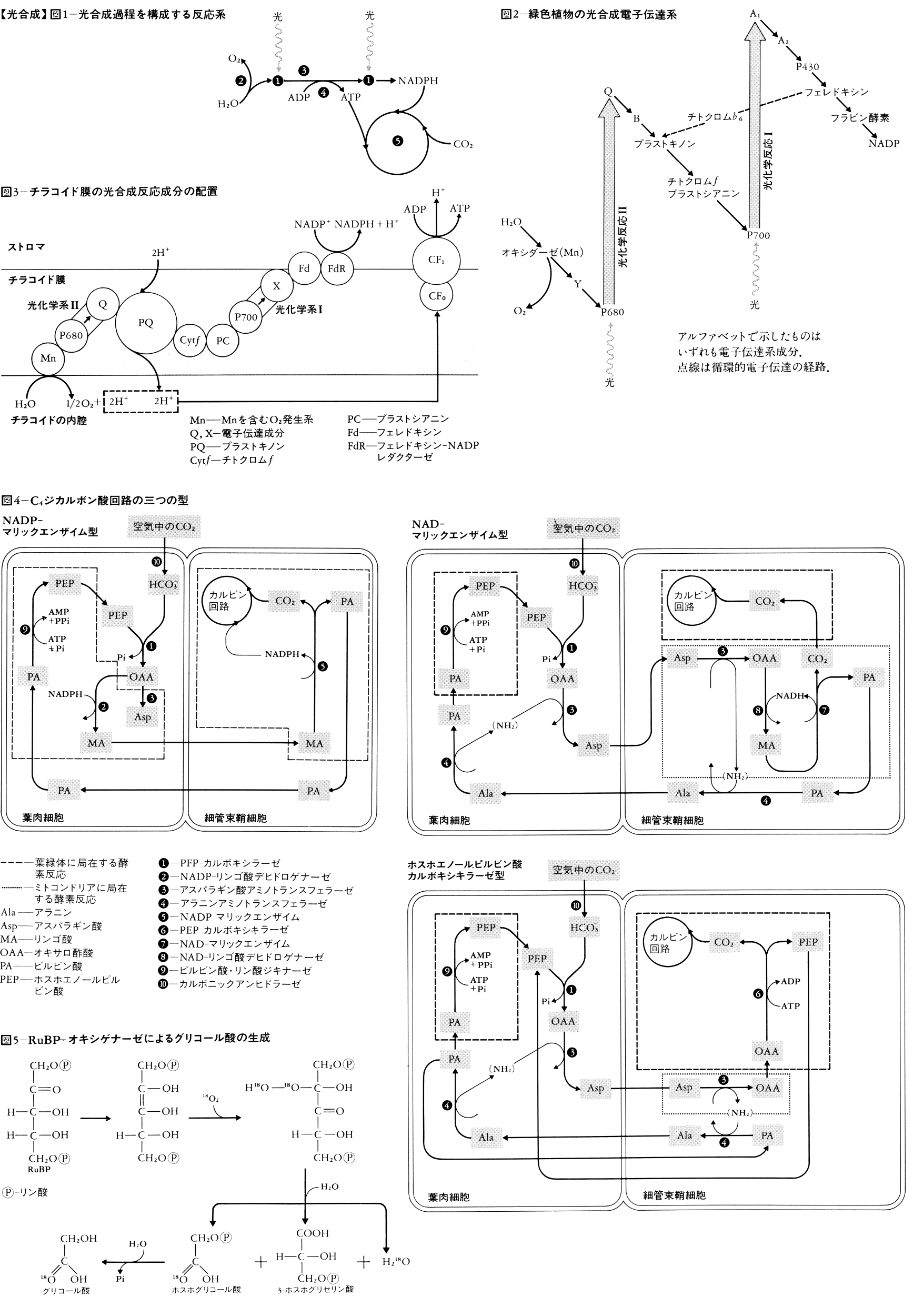
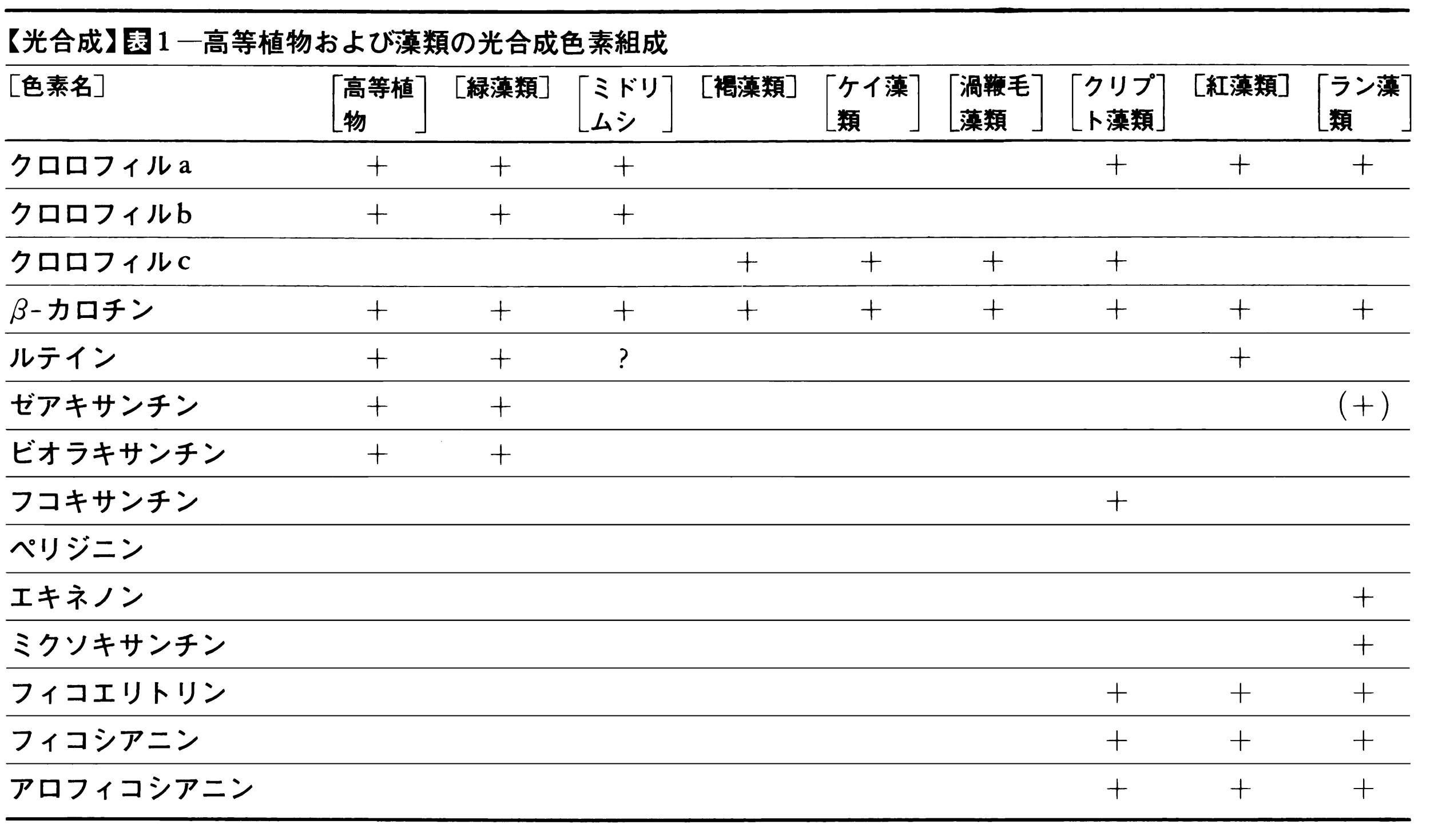
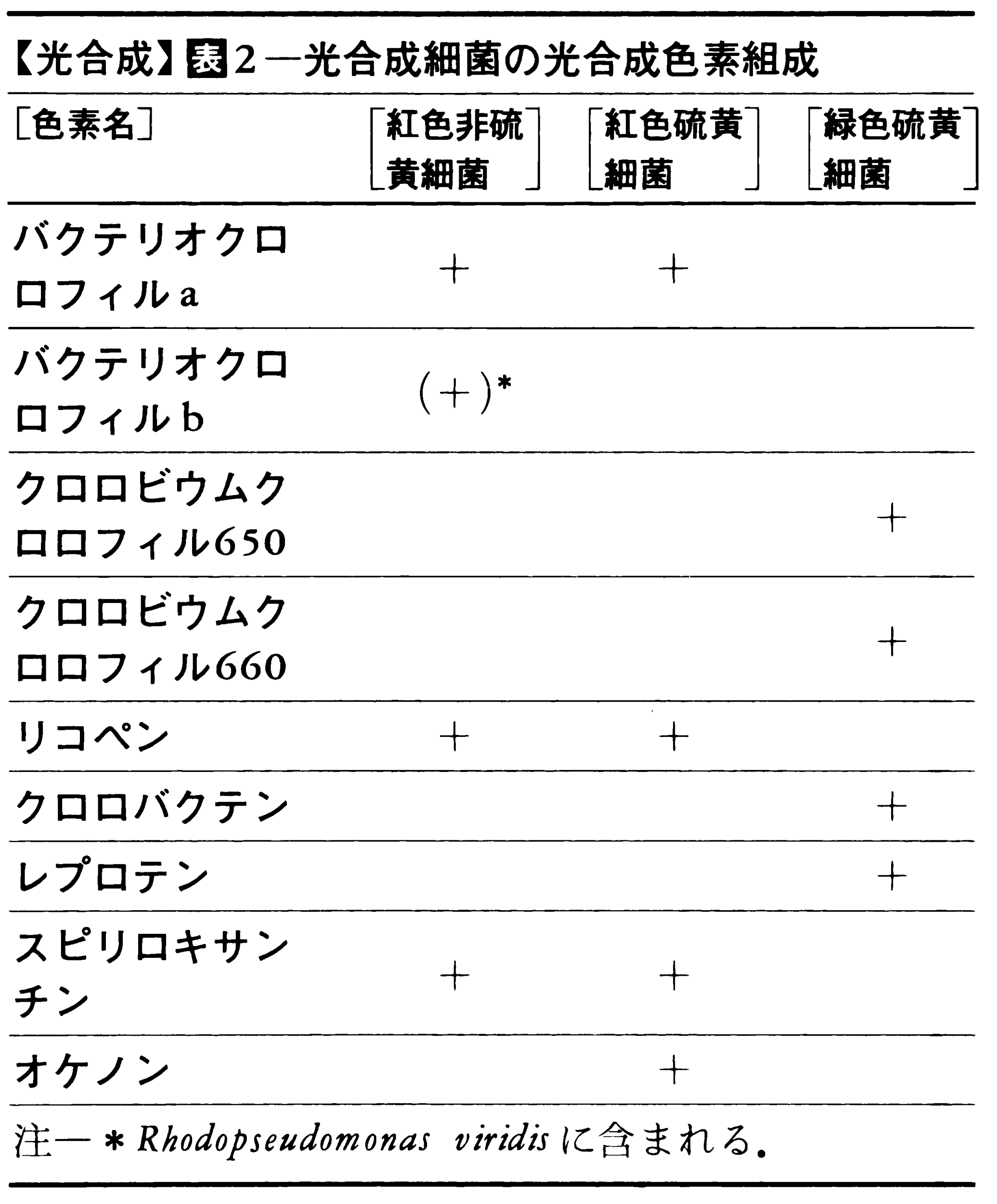
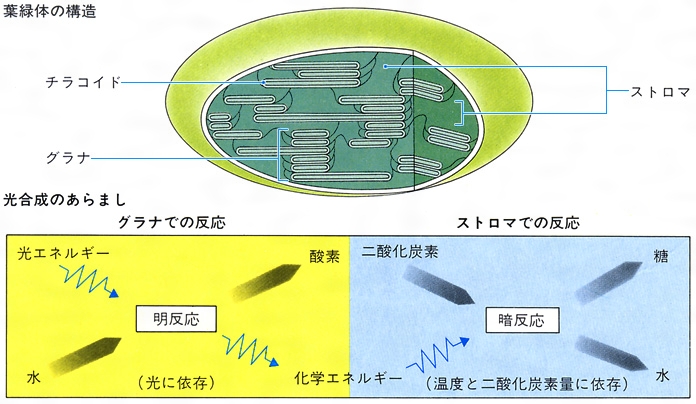
緑色植物の光合成は(1)初期光化学反応,(2)O2発生反応,(3)電子伝達反応,(4)光リン酸化反応,(5)CO2固定反応から成り立っている(図1)。(1)~(4)が古典的には明反応light reactionと呼ばれていた系で,葉緑体のチラコイド膜に局在し,(5)はストロマに局在する。自然界には多くの光合成色素(同化色素)が存在するが,その組成は光合成生物の系統,類縁によって定まっている(表1,表2)。
光合成は同化色素が光エネルギーを吸収することによって始まる。光量子によって励起された色素分子は,そのエネルギーを最終的に反応中心と呼ばれる特別なクロロフィル(葉緑素)a(またはバクテリオクロロフィルa)分子に伝達し,そこで光化学反応が行われる。これらの同化色素分子は,集合して機能的な単位体を作っていると考えられている。クロロフィル約300分子に対し,反応中心のクロロフィル分子が1個あると推定されており,このような1組を光合成単位photosynthetic unitという。光合成単位としては,主としてクロロフィルaに吸収される光によって反応が行われる光化学系Ⅰ(PS Ⅰ)に属するものと,主として補助色素に吸収される光によって反応が行われる光化学系Ⅱ(PS Ⅱ)に属するものとの2種がある。PS ⅠおよびPS Ⅱの反応中心のクロロフィルaは,光により酸化されるとき,吸光度減少の起こる波長の値を付して,それぞれP700,P680と呼ばれる。クロロフィルはタンパク質と結合し,クロロフィル-タンパク質複合体として存在し,これが一定の様式で集合し,それぞれPSⅠ,PS Ⅱの光合成単位を構成している。緑色植物ではP700またはP680を結合したクロロフィル-タンパク質複合体,反応中心をもたずクロロフィルaとbを1:1に含む集光性クロロフィルa/b-タンパク質複合体がよく知られている。
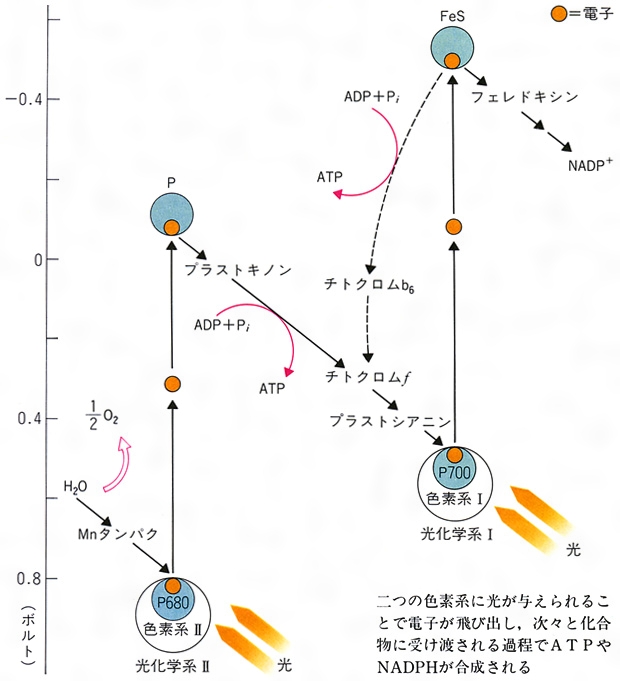
PSⅡの反応中心が励起されると,最初の電子受容体Qを還元し,他方,水を酸化して電子を受けとるとともにO2を発生する(図2)。Qの電子は電子伝達系を経てP700へ渡される。P700も光を受けると酸化され,電子はA1を経て最終的にNADPに渡される。この反応経路の模式図はZのような形をしているのでZスキームと呼ばれる。1957年エマーソンR.Emerson(1903-59)は,クロロフィルaの赤色域の吸収極大より長波長側の光(700nm)しか与えないと,緑色植物や藻類の光合成の量子収量が低下する(赤色低下red drop)が,ここへより短波長の光(緑色植物なら650nm,紅藻なら575nm)を同時に与えると,光合成速度は両者を単独で与えたときの和より大きくなることを見いだした(エマーソン効果)。光合成細菌ではこの現象はみられない。エマーソンの発見がきっかけとなって,光合成の初期過程には二つの光化学系があるという,現在のZスキームに至る研究が発展した。光合成単位はZスキームの他の成分とともに,一定の配列でチラコイド膜に組み込まれていると考えられている(図3)。ただし,PSⅡの分布には偏りがあり,チラコイド膜どうしが重なっている部分にあるといわれている。
発生するO2は水に由来するので,反応機構を考慮に入れた場合には光合成の全反応は6CO2+12H2O─→C6H12O6+6O2+6H2OまたNADP還元までの部分反応は12NADP⁺+12H2O─→12NADPH+12H⁺+6O2と書く。O2発生機構としては,2分子の水の酸化が4段階を経て行われ,これにマンガンMnが関与していることが知られている。Zスキームの電子伝達と共役してADP(アデノシン二リン酸)とオルトリン酸からATP(アデノシン三リン酸)が合成される(光リン酸化という)。Zスキームに沿った一方向的な電子伝達と共役するものを非循環的光リン酸化,循環的電子伝達経路(クロロフィルから出た電子が電子伝達系の途中から再びクロロフィルへ返ってくる)と共役するものを循環的光リン酸化という。前者では反応産物の比O2:ATP:NADPHは1:2:2である。循環・非循環両形式の比率の調節機構があるものと考えられている。光リン酸化には,チラコイド膜の外表面(ストロマ側)に結合したCF1と,チラコイド膜内に結合したCF0と呼ばれるタンパク質が関与する。電子伝達の結果生成したATPとNADPHは炭酸固定系の反応に用いられる。RuBPC(リブロース-1,5-二リン酸カルボキシラーゼ),NADP-グリセロアルデヒド-3-リン酸デヒドロゲナーゼ,リブロース-5-リン酸キナーゼなど,炭酸固定系の主要な酵素の活性は光によって制御される。制御機構は2種に大別される。すなわち,光があたり光化学系が働いた結果起こる,(1)チラコイドへのH⁺の取込み,チラコイドからのマグネシウムイオンMg2⁺の放出,葉緑体へのカルシウムイオンCa2⁺の取込みによるストロマのイオン環境の変化,(2)チラコイドの電子伝達系の還元により酵素活性調節に関与する物質の還元,によって制御される。
光合成によるCO2固定速度を光合成速度と呼ぶが,一般にその測定は単位時間当りのCO2の吸収量またはO2の発生量によってなされる。このような測定値は植物が呼吸によって生じたCO2,消費したO2を差し引いたものであるから,〈純(あるいは見かけ上の)光合成速度〉と呼んで〈真の(あるいは総)光合成速度〉から区別する。
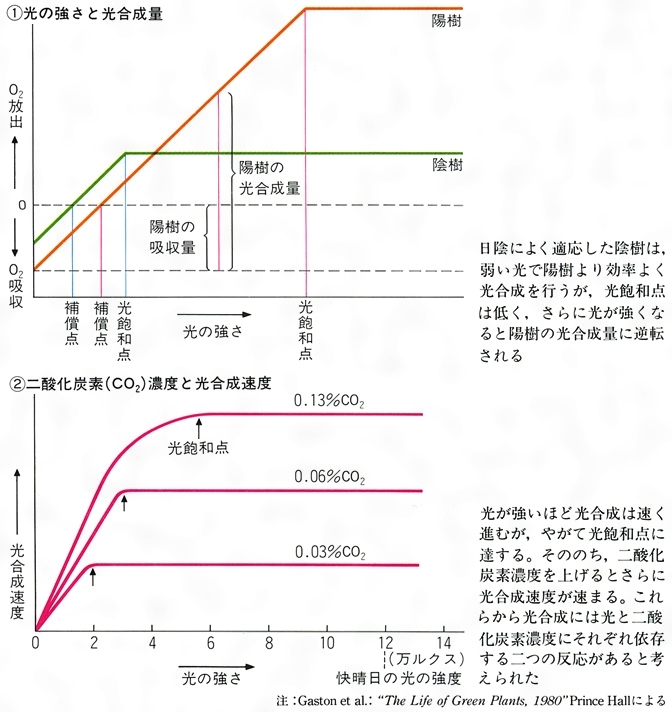
光合成速度を左右する最も大きな環境要因は光で,光の強さと光合成速度の間には双曲線的な関係がみられる。まず,光の量が増加して,真の光合成速度が呼吸速度に等しくなると,純光合成速度が0になる。このときの光の強さを〈光補償点〉と呼ぶ。光の増加とともに光合成速度は速くなるが,ある程度以上光が強くなると,光合成速度は飽和状態に達し,もはやその速度は光の量とは無関係になる。このときの光の強さを〈光飽和点〉,光合成速度を〈飽和光合成速度〉と呼ぶ。
〈光補償点〉〈光飽和点〉〈飽和光合成速度〉は,植物の種によってきまっているが,一般に陽樹のほうが陰樹よりすべての値が高いのがふつうである。
光合成速度を左右するもう一つの大きな要因は大気中のCO2濃度で,とくに光飽和に達して以後の光合成速度増加にとって重要である。CO2濃度も光と同じように光合成速度との間に双曲線的な関係をもち,〈CO2補償点〉つまり純光合成速度が0になるCO2濃度が存在する。
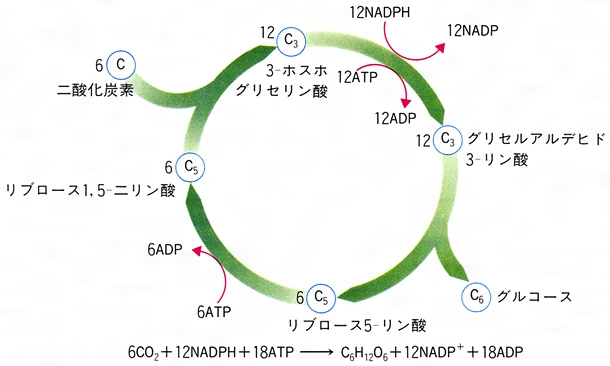
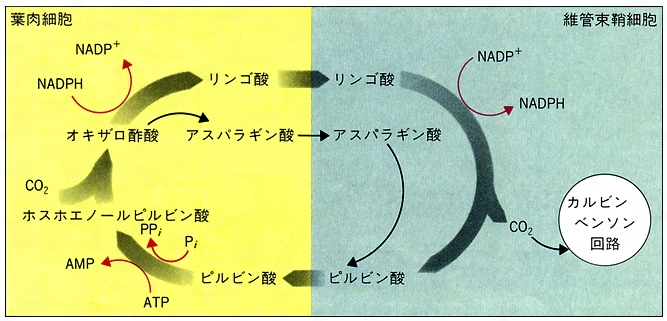
光合成の炭酸固定系にはカルビン回路とC4ジカルボン酸回路(C4回路)の2種類がある。前者をもつ植物(C3植物)はRuBPCによってCO2固定を行う。C4回路をもつ植物(C4植物)では,まず葉肉細胞でHCO3⁻を基質としてホスホエノールピルビン酸カルボキシラーゼ(PEPC)でCO2を固定し,オキサロ酢酸を生成し,これからリンゴ酸またはアスパラギン酸を生じ,これが維管束鞘(いかんそくしよう)細胞へ送られたのち,そこでこれらの化合物からCO2を放出し,これをカルビン回路で再固定する。この反応経路のうち,カルビン回路より前の過程を狭義のC4回路という。C4回路(広義)はカルビン回路に対して,効率のよいCO2供給系(狭義のC4回路)が付け加えられたものである。C4植物は維管束鞘細胞に多数の葉緑体をもつ。C4植物は,葉肉細胞で作られたCO2固定産物から維管束鞘細胞でCO2を放出させる反応の種類によって三つのタイプ,すなわち(1)NADP-マリックエンザイム型,(2)NAD-マリックエンザイム型,(3)ホスホエノールピルビン酸カルボキシラーゼ型に分類される(図4)。C4植物としては,イネ科,カヤツリグサ科などに属するもののほか,双子葉植物を含む20科1100種が知られており,熱帯原産のものが多い。CO2補償点がきわめて低く(5ppm以下),強光,高温,低CO2濃度中での光合成速度はC3植物より高い。ベンケイソウ科の植物では,夜間に気孔が開いてCO2を吸収し,これがリンゴ酸として蓄えられ,昼間は気孔が閉じてリンゴ酸が減少し,デンプンを形成するという日変化を示すことが古くから知られており,この代謝形式は,ベンケイソウ型有機酸代謝crassulacean acid metabolism(CAM)と呼ばれていた。C4植物の研究の進展の結果,C4植物では葉肉細胞と維管束鞘細胞との協調で2種のCO2固定反応が空間的に分けて行われているのに対し,CAM植物ではこの2種の反応が夜と昼というように時間的に分けて行われる,すなわち夜間PEPCによるCO2固定が行われ,昼間はこのCO2がカルビン回路で再固定されることが明らかにされた。CAM植物は乾燥に対する適応として特異的な代謝系をもつに至ったものと考えられる。現在,ベンケイソウ科のほかサボテン科,アナナス科などの112属300種余り(多肉のものが多い)がCAM植物として知られている。
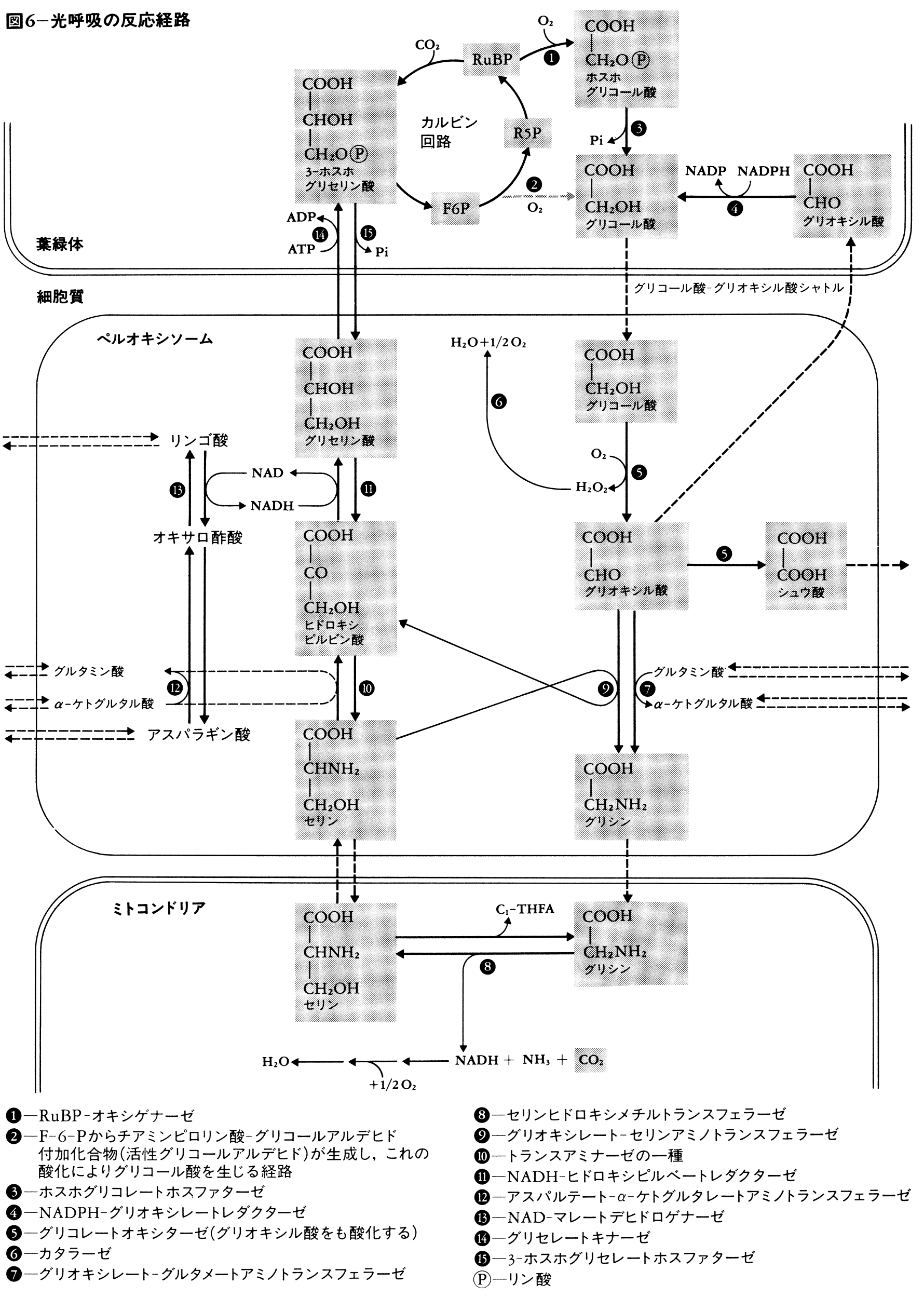
光のもとでカルビン回路に由来するグリコール酸が,グリコール酸経路により酸化分解される過程を光呼吸photorespirationという。RuBPCはオキシゲナーゼ活性もそなえており(リブロース-1,5-二リン酸カルボキシラーゼ/オキシゲナーゼRuBisCOと呼ばれる),ホスホグリコール酸を生じる(図5)。これが呼吸基質となりマイクロボディ(パーオキシソーム)とミトコンドリアの関与によって,図6の経路を経て酸化される。光呼吸は,過剰の還元力を消費することに役だっているのではないかとも考えられているが,生理学的意味はまだ完全には明らかではない。C3植物では,強光,高温,高O2濃度,低CO2濃度という条件で,光呼吸は増大し見かけの光合成は低下するが,C4植物ではCO2の回収が速いためこのような低下はみられない。
→葉緑素
執筆者:辻 英夫
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
緑色植物や光合成細菌が太陽光エネルギーを用いて、二酸化炭素から糖類などの有機物を合成すること。炭酸同化(炭酸固定)の一種である。地球上の生物は、生命の維持のためにエネルギーを必要とするが、化学合成細菌を除くすべての生物は、直接あるいは間接に太陽光エネルギーに依存している。緑色植物や光合成細菌は、直接太陽光エネルギーを利用できるが、動物などの従属栄養生物は、植物を食べるか、または草食動物を食べてエネルギーを得ているので、間接的に太陽光エネルギーを利用していることになる。われわれが燃料として使っている石油、石炭、天然ガスも何百万年も前の動植物の分解物であるから、これらの燃料に蓄えられたエネルギーは、太陽光エネルギーの変形したものといえる。このように、地球上の生命にとって光合成による太陽光エネルギーの捕捉(ほそく)は不可欠であり、もし植物がこのような働きをしなかったなら、地球上の生命は、とっくに消滅していたに違いない。毎年地球の表面に降り注ぐ太陽光のうち、光合成に利用できる波長域のものは半分で、しかもその40%は海や砂漠で反射されるので、光合成生物が利用している光のエネルギーはさらに少ない。それでも光合成生物が1年間に光合成によって生産するエネルギー量は、糖に換算して1000億トンに達するといわれる。これは地球上で利用できる太陽光エネルギーの0.2%であり、さらにこのうちの0.5%以下が人類の食糧として消費されているにすぎない。なお、光合成によるエネルギー生産量の40%は海面近くにいる植物プランクトンによって行われている。地球上の人口の増加とともに、食糧エネルギー問題が深刻となっているが、近年、その解決のための光合成の利用が脚光を浴びている。
[吉田精一]
植物の光合成は、種々の環境要因、とくに光の強さ、温度、二酸化炭素濃度によって左右される。酸素と二酸化炭素の出入りに関しては、光合成と呼吸は逆の関係にある。暗所では植物は呼吸だけを行うので、酸素の吸収だけがみられるが、弱い光を照射してすこしずつ光の強さを増すと、光合成による酸素の放出と呼吸による酸素吸収が等しくなり、見かけ上ガス交換がみられなくなる。このときの光の強さを「光(こう)補償点」という。補償点よりさらに光を強くすると、酸素放出は直線的に増加するが、ある程度の光の強さに達すると光合成速度は頭打ちになる。このとき、二酸化炭素濃度または温度を上げると、光合成速度はまた上昇する。この事実からイギリスの植物生理学者ブラックマンFrederick Frost Blackman(1866―1947)は、光合成の反応は、光に依存した光化学反応(明反応)と、二酸化炭素濃度や温度によって影響される化学反応(暗反応)の二つからなることを示唆した。つまり明反応は、光のエネルギーが化学エネルギーに変えられる過程であり、この過程で酸素が放出される。一方、暗反応は、明反応でつくられた化学エネルギーを用いて二酸化炭素から糖が合成される過程といえる。
[吉田精一]
植物細胞から単離した葉緑体には、光のもとで二酸化炭素からの糖の合成、酸素の発生など、光合成のすべての反応を行う能力が備わっている。高等植物の葉緑体は、直径4~10マイクロメートル、厚さ1マイクロメートルの円盤状で、二重膜で包まれ、その内側は層状のラメラ(層板ともいう)、あるいはチラコイド構造(扁平(へんぺい)な袋状)の膜系がある。この膜系が積み重なったものをグラナという。この部分に明反応に関係するすべての色素と反応系が存在する。一方、チラコイドの周りにある無色の部分はストロマとよばれ、ここに二酸化炭素から糖を合成する暗反応の酵素群が含まれる。
[吉田精一]
植物の葉緑体にはクロロフィル類のほか、カロチノイドやフィコビリンなどの色素が存在している。植物にいろいろの波長の光を照射して、その波長での光合成速度を調べた作用スペクトルをみると、クロロフィルaの最大吸収を示す420ナノメートルと660ナノメートルの波長域だけでなく、他の波長のところでも光合成がかなり効率よく行われていることがわかる。このことは、それらの波長域の光を吸収する色素が光合成に関与していることを示している。事実、ほかのクロロフィル類、カロチノイド、フィコビリンなどの色素がその吸収した光のエネルギーをクロロフィルaに受け渡ししていることが明らかとなり、これらの色素を「補助色素」とよんでいる。アメリカの植物生理学者エマーソンRobert Emerson(1903―1959)は藻類の光合成作用スペクトルを研究し、波長が680ナノメートル以上になると光合成効率が著しく低下する、いわゆる「赤色低下」現象をみいだした。このとき、短波長側の光を同時に照射すると、それぞれの光を単独に与えたときの光合成量の和より大きくなった。このことから、植物の光合成には二つの光化学反応が関与していることが明らかとなり、それぞれ光化学系Ⅰ、光化学系Ⅱとよんでいる。
光化学系Ⅰは、主としてクロロフィルaを色素系として含み、これに光が当たるとクロロフィルが励起されて電子がはじき出され、この電子がフェレドキシン(電子運搬体の作用をもつ小分子のタンパク質)を還元し、最終的にNADPH(還元型のニコチン酸アミドアデニンジヌクレオチドリン酸)が生成され、電子を失った状態になる。一方、クロロフィルbやカロチノイドなどの補助色素を多く含む光化学系Ⅱに光が当たると、同様に電子がはじき出され、この電子はチトクロム類を含む一連の電子伝達系を通って光化学系Ⅰの電子の「穴」に送られ、この過程でATP(アデノシン三リン酸)が生産される。電子を失った光化学系Ⅱは、水の光分解によって電子を得ると同時に酸素を放出する。このような二つの光化学系の共役によるATPの生成を「非循環的光リン酸化」という。また、光化学系Ⅰと光化学系Ⅱによる電子伝達系は、「Z経路」とよばれ、この経路によってATPとNADPHが等量生成される。しかし、暗反応で二酸化炭素から糖を合成するには、1分子のNADPH当り1.5分子のATPが必要であり、この経路だけではATPが足りない。この不足を補うため、光化学系Ⅰのみを使う循環的光リン酸化反応というのがある。これは、光化学系Ⅰに光が当たって放出された電子は、別の電子伝達系を通ってふたたび光化学系Ⅰに戻され、その過程でATPが生産されるという過程である。この過程では酸素の発生はみられない。葉緑体のチラコイドで行われるこれらの明反応によって生成したATPとNADPHは、ストロマ(葉緑体の中でラメラを包んでいる無色の基質)に送られて暗反応に用いられる。
ストロマには二酸化炭素から糖を合成する経路が存在する。この経路は糖の分解経路である「ペントースリン酸回路」の逆の経路であるので、「還元的ペントースリン酸回路」、あるいは発見者の名をとって「カルビン‐ベンソン回路」とよばれる。この経路では、初めにペントース(五炭糖)のリブロース二リン酸に二酸化炭素が結合して2分子のホスホグリセリン酸が生成し、これからNADPHによる還元とATPによるリン酸化を経て糖が生成し、その経路の回転によってリブロース二リン酸が再生されてくる回路となっている。この回路の回転によって6分子の二酸化炭素から12分子のNADPHと18分子のATPを用いて1分子の糖が合成される。
最近になって、植物が光合成を盛んに行っているとき、同時に活発に呼吸していることが明らかとなった。このような光のもとでの呼吸は、普通の呼吸とはまったく異なる仕組みで行われることがわかり、「光呼吸(こうこきゅう)」または「光呼吸(ひかりこきゅう)」とよばれる。これは、カルビン‐ベンソン回路の炭酸同化を行う酵素が、酸素分圧の高いときにはリブロース二リン酸を分解してホスホグリコール酸を生成するためで、グリコール酸経路とよばれる。この経路による光呼吸の反応は、葉緑体、ペルオキシゾーム、ミトコンドリアの細胞内顆粒(かりゅう)の共同作業で行われる。光呼吸活性が高いと、必然的に光合成効率は低下する。光合成効率の悪い植物では、光呼吸は純光合成の50%に達することもある。光呼吸は、光が強く、大気中の酸素分圧が高く、二酸化炭素分圧が低いほど高くなる。
サトウキビ、トウモロコシなどの熱帯性草本では光呼吸が非常に低い。これら植物では光合成の際の二酸化炭素固定の初期産物がリンゴ酸、アスパラギン酸などのC4ジカルボン酸であるためであり、このような植物をC4植物とよび、光合成の初期産物が糖であるC3植物とは区別される。C4植物には、はっきりとわかる2種類の葉緑体の型がある。その一つは葉肉細胞に葉緑体が存在する型であり、もう一つは維管束鞘(しょう)細胞に局在する型である。C3植物では、葉緑体はほとんど葉肉細胞にのみ存在する。C4植物には炭酸同化経路としてC4ジカルボン酸回路があり、気孔から入った二酸化炭素は、まず葉肉細胞でホスホエノールピルビン酸カルボキシラーゼによって固定されてオキサロ酢酸となり、これからリンゴ酸やアスパラギン酸のC4ジカルボン酸が生成する。これが維管束鞘細胞に送られて、そこで脱炭酸されて二酸化炭素を再生し、これがこの細胞に局在するカルビン‐ベンソン回路によって固定されて、糖を合成する。C4植物にこのような複合経路が存在する理由は、熱帯のような二酸化炭素濃度の低い環境では、カルビン‐ベンソン回路の炭酸同化酵素よりも二酸化炭素に対する親和性の高いホスホエノールピルビン酸カルボキシラーゼでまず炭酸同化を行うためである。
密閉した容器内に植物を置いて、十分に光を照射したとき、最終的に一定になった容器内の二酸化炭素濃度を二酸化炭素補償点という。温帯性植物のC3植物ではこの値は50ppm以上であるが、C4植物ではきわめて低いことが特徴で、光呼吸の小さいこととともに太陽光エネルギーの利用効率が高いことがわかる。
ベンケイソウ、サボテンなどの多肉植物はC4植物と同様の炭酸同化経路をもっている。これらの植物の葉は夜間にリンゴ酸を蓄積するため酸度が高くなり、日中はリンゴ酸が消失して酸度が低くなる。このような昼夜の有機酸の変動をCAM(キャム)(ベンケイソウ型有機酸代謝)といい、これを顕著に行う植物をCAM植物とよんでいる。CAM植物は乾燥地域に生育するため、日中は蒸散を防ぐために気孔がほとんど閉じており、光合成のための二酸化炭素が吸収されない。そのため夜間に気孔が開いて、そのとき吸収した二酸化炭素をリンゴ酸の形で蓄積し、日中にこのリンゴ酸の脱炭酸によって生成した二酸化炭素を光合成に利用しているわけである。
[吉田精一]
光合成細菌の細胞内にはクロマトホアとよばれる粒子があり、ここで光合成が行われる。細菌の光合成では、酸素の発生がない、水素や硫化水素が電子供与体となる、遠赤外光も光合成に利用できるなど、いくつかの点で植物の光合成と違っている。明反応では光化学系は一つしかないと考えられているが、植物と同様、光リン酸化反応を行い、ATPを生成するとともにNADPHも生成する。暗反応もカルビン‐ベンソン回路によって炭酸同化を行っている。最近、一部の光合成細菌ではクエン酸回路の逆の経路で炭酸同化を行っていることが知られ、この経路を還元的カルボン酸回路とよんでいる。
[吉田精一]
『D・O・ホール、K・K・ラオ著、金井龍二訳『光合成』(1980・朝倉書店)』▽『藤茂宏著『光合成』(1982・東京大学出版会)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
高等植物,藻類,光合成細菌が行っている太陽光エネルギーによる炭酸同化の生物過程をいい,生物における無機物から有機物を合成する独立栄養の一形式である.高等植物および藻類では,光エネルギーを利用して二酸化炭素が水によって還元され,糖が合成されると同時に酸素が発生するが,
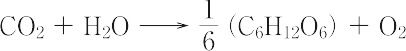
光合成細菌では,水のかわりに硫化水素などが酸化されるので酸素は発生しない.光合成では,まず光がクロロフィル,カロテノイドなどの色素類によって吸収され,そのエネルギーによって,シトクロム,プラストキノン,プラストシアニン,フェレドキシンなどの電子伝達系を通じて,水分子が酸化されてニコチンアミドアデニンジヌクレオチドリン酸(NADP)が還元されると同時に,光リン酸化が起こってアデノシン5′-三リン酸(ATP)が合成される.生成されたNADPHの還元力とATPのエネルギーは,ついでカルビンサイクルまたは C4-ジカルボン酸サイクルなどの光合成サイクルによりCO2を糖,さらにはデンプンへと還元する.地球上では光合成により炭素として年間5×1010 t,有機物として年間12.5×1010 t の物質が生合成されている.これは陸上では(2.2~3.2)×1010 t の炭素,海洋では(2.2~2.8)×1010 t の炭素に相当する.
出典 森北出版「化学辞典(第2版)」化学辞典 第2版について 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
Sponserd by ![]()
…光(ひかり)合成ともいう。植物が光のエネルギーを利用して二酸化炭素CO2と水H2Oから有機化合物を合成する過程。…
※「光合成」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
半夏ともいう。七十二候の一つで,本来は夏至後 10日目から小暑の前日までをいったが,現行暦では太陽の黄経が 100°に達する日 (7月1日か2日) を半夏生とし,雑節の一つとして記載している。この頃半...