高等な植物(維管束植物)のからだは根,茎,葉の三つの基本的な器官から成り立っている。茎と根が軸性の器官であるのに対して,葉は茎に側生する器官で,平面的な広がりをもつ。コケ植物や藻類でも,植物体のうち平面的な構造を葉ということがあるが,維管束植物の葉と相同の器官ではない。コケや藻類の植物体を葉状体ともいうが,コケなどの葉は,形態学的に厳密にいえば葉的器官というべきものである。
形状
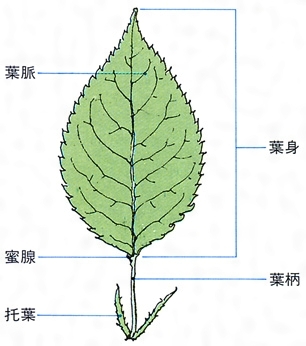
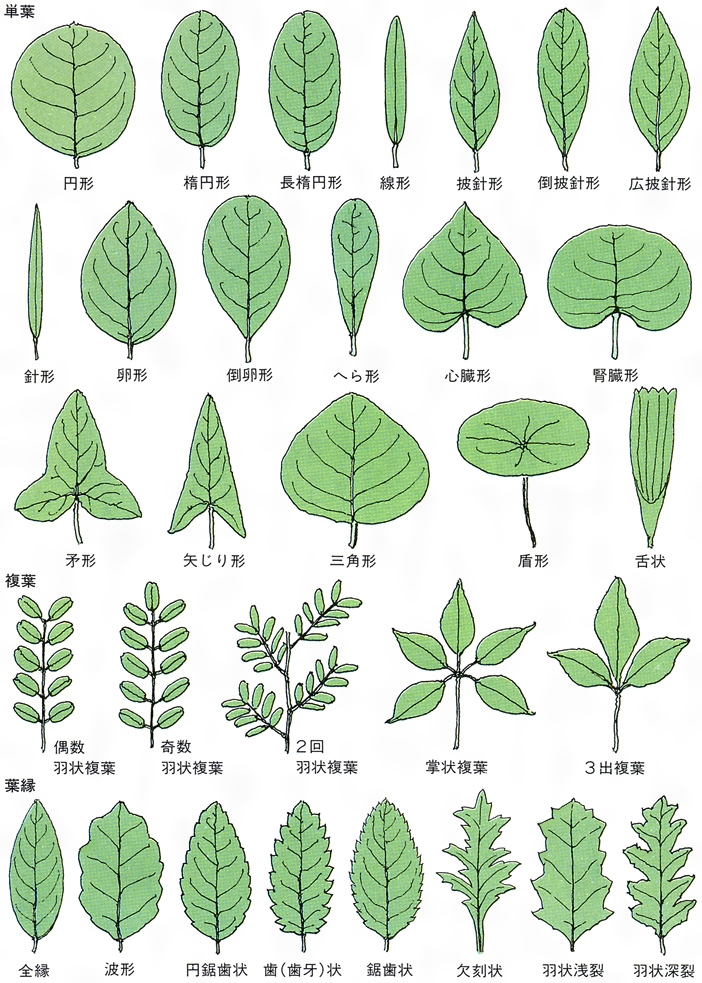
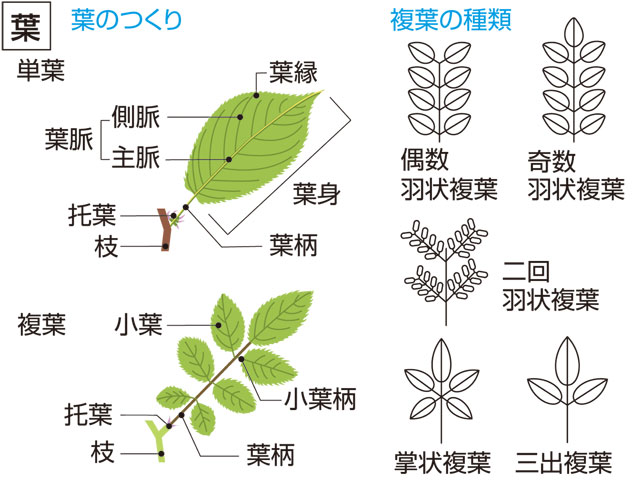
葉にはさまざまの形のものがある。大きな葉としてはバショウやヤシの葉の例があげられるが,ウラジロやカニクサのように葉が無限に展開していくものでは,数十mにも伸びる。小さい葉の例としてはウキクサの仲間の例や,コケシノブ科には0.5mmに達しないものもある。葉は葉柄と葉身からできており,托葉をもつものもある。葉柄petiole(stipe)は発達しないものもあるが,一見茎のようにしっかりしているものもある。毛や鱗片などの付属物をつけている場合も多い。葉身lamina(blade)の構成にも変化があり,単葉と複葉に二大別できる。単葉は葉身が単一の面でできたもので,辺縁は全縁のものから深く切れ込むものまでさまざまで,ほとんど中肋近くまで切れ込むものでは複葉との差がはっきりしない。複葉は,葉身が軸とそれから分岐した複数の面とでできているもので,構成にはいろいろのものがある。羽状複葉が最も多い例で,中軸とそれから羽状に分岐した羽軸を基本とする。エンドウのように1回羽状に分岐する複葉や,ナンテンのように2~3回羽状に分岐するものがあり,シダ植物の葉のように4~5回羽状分岐する複葉もある。掌状に分岐する例としてはシュロやクジャクシダなどがある(掌状複葉)。コシダ類の葉は叉(さ)状に分岐する軸と,そのうちのいくつかの部分の欠落と,羽状に分岐する小葉との組合せで,さまざまな型の構成を種や変種の差によって示している。被子植物の葉には葉柄の基部近くに托葉stipuleをつけるものがある。
葉から変態してつくられた相同器官には,サボテンのとげ,エンドウなどの巻きひげ,ウツボカズラの漏斗状の補虫器官などいろいろの例があげられるが,最も顕著なものとしては,花被片をはじめとする花を構成する要素への変態があげられる。
→花
構造
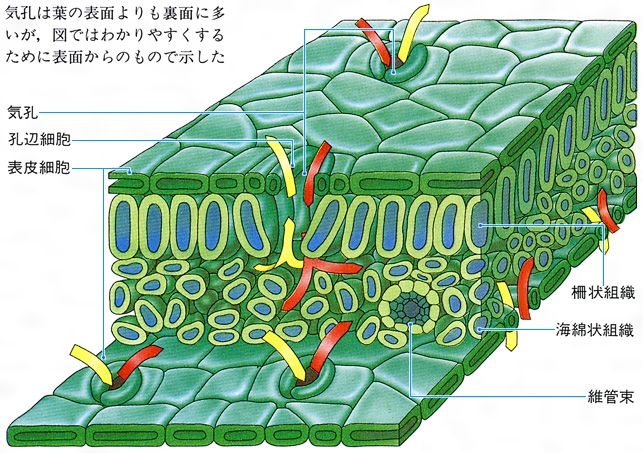
被子植物の葉は,外側に1層(まれに多層になることもある。ムラサキツユクサなど)の表皮があり,おもに裏面に気孔がみられる。気孔は2個の孔辺細胞の間にできる穴であるが,孔辺細胞の周辺に助細胞など,特殊な細胞がつくられることもある。表皮細胞には葉緑体が含まれていないが,孔辺細胞には葉緑体がつくられる。シダ植物の葉も表皮の構造はよく似ているが,すべての細胞に葉緑体がある点で異なっている。表皮にはクチクラ層が発達しており,水分が過度に蒸散しないようになっている。表皮組織の下には長方形の細胞が密に整列した柵状組織があり,その内側には細胞間隙(かんげき)の発達した海綿状組織があって,それらの組織で光合成が行われる。これらの基本組織をひっくるめて葉肉と呼ぶことがある。シダ植物などの葉では,柵状組織と海綿状組織の差ははっきりしない。
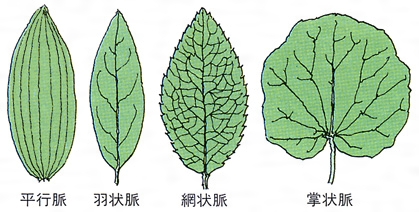
茎から葉が側生するとき,茎の維管束が枝分れする。茎から離れて葉柄へ入っていく維管束を葉跡leaf traceという。茎の維管束から葉跡が分出するとき,茎の維管束にできる間隙を葉隙leaf gapというが,原生中心柱の場合,葉隙はつくられない。葉跡は1本だけの場合もあるが,一つの葉柄に複数の葉跡が入るのがふつうで,その数は種によって一定していることと,不定のことがある。シダ植物などで,葉跡の数や構造が分類群のよい指標になることもある。葉跡は葉柄の維管束で,さらに枝分れする。複葉では中軸の維管束が分枝して羽軸に入り,枝分れして小葉や裂片の主脈となる。単葉の場合,葉跡はそのまま葉の中肋の維管束となり,それから枝分れした維管束が葉脈veinとなる。葉身にいろいろの模様をつくる葉脈は,葉へ入ってきた維管束とそれをとり巻く特殊な組織とでつくられる構造である。葉脈が葉身につくる模様を脈理venationという。脈理は植物群によって一定で,シダ植物では遊離脈といろいろの段階に発達した網状脈があり,いずれの場合も脈の末端は開放二叉分岐を基本としている。双子葉植物ではほとんどのもので複雑な網状脈をもっており,単子葉植物でも細脈は網目をつくるが,主脈は平行に走るので平行脈といわれる。ヒカゲノカズラ類の葉では脈は単生する。
形成
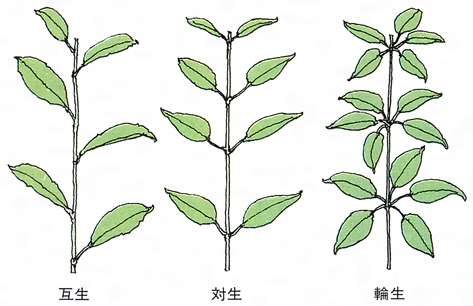
葉は茎に側生するが,茎頂付近で茎の生長点よりやや下がった位置に葉原基leaf primordiumができる。葉原基は少し大きくなると上下の二つの部分がはっきりしてきて,下部の葉基からは葉鞘(ようしよう)と托葉,上部の上葉から葉身と葉柄がつくられる。葉鞘 leaf sheathは単子葉植物によくみられるもので,幼葉や生長点を保護している。双子葉植物には葉鞘は珍しいが,キンポウゲやナンテンなどには発達している。托葉は変化に富んでおり,まったく無いものもある。レンリソウ属の1種では,本葉が巻きひげになり,托葉が栄養器官になっている。単子葉植物には托葉をもつものは少なく,シダ植物には葉鞘も托葉も発達しない。葉身は上葉で原型がつくられ,辺縁生長がくり返されて完成され,葉が展開するときには,すでにできて畳み込まれていたものが,個々の細胞の生長につれて伸びてきて葉の形となる。なお苗条(シュート)の上での葉の配列のしかたを葉序phyllotaxisという。
進化
葉には二つの種類があるというのが今日ほぼ定説となっている見方である。ヒカゲノカズラやミズニラの葉は小葉microphyllで,これはもともと茎の表面に突起状のものがつくられ,それが発達して維管束の分枝を受け入れるようになり,側生器官として完成されたものである。小葉は原則として葉脈が単生であり,葉跡は葉隙をつくらないといわれるが,例外がないわけではない。大部分の維管束植物の葉は,これに対して大葉megaphyllといわれる。大葉はもとは一つの苗条からつくられたものであるが,苗条から大葉への進化はテロム説telome theoryで説明される。原始陸上植物は1回ごとに分裂の軸が90度回転して,マツバランのような立体的な二叉分岐をした苗条をもっていたが,この1単位のテロム枝が,扁平化,単軸化,融合,単純化などの進化をくり返して葉となった。だから,1枚の大葉は1単位の苗条であり,脈はもともとの枝のなごりである。もちろん,葉脈は葉が完成されてからいろいろの模様をつくるように進化してきたものだから,網状にからみ合った苗条があったことはない。原初の葉はすべて遊離脈をもっており,石炭紀になって網状脈をもった裸子植物が進化してくる。
大葉と小葉はその系統発生の過程が異なっているので,機能などはほとんど同じであるにもかかわらず,相同な器官とはいいがたい。すなわち,一方は苗条からできた葉であり,他方は茎の皮層などを主とした葉だからである。これら2種類の葉は,それぞれ異なった進化の道をたどってきた二つの植物群に別々に見られるもので,維管束植物には二つの系統群があるというのが現在では有力な説になっている。しかし,テロム説にしても,進化過程のすべてが実証的に示されたとはいえず,植物の進化にはまだ解明されていない問題も多いので,維管束植物に二つの系統群を認めるというのも仮説の域をでない。
コケ植物のうちにも,たとえばスギゴケにみるように,維管束植物とよく似た葉的器官がみられ,記載用語では実際に葉といわれている。しかし,この葉は細胞層1層の構成で,中肋には維管束はみられない。個体発生でみれば,茎の先端にある三角錐形の頂端細胞が3方向に細胞を切り出し,そのうち1個の列が発達して茎を,他の2個が葉と呼ばれる構造をつくる。維管束植物の大葉,小葉のいずれも,形態形成の方法,系統発生において,まったく異なったものである。藻類の場合も,扁平な側生器官で光合成の機能の高いものを葉と呼ぶことがあるが,形態的に維管束植物の葉と相同のものではない。
役割
植物が陸上へ進出してさまざまに進化したのは,陸上で生産の効率を高めたことに負う部分が大きいが,葉は光合成の主役を演じる構造であり,それだけに陸上における植物の多様化と機能上の進化に果たした葉の意義は大きいものがある。陸上の生活環境が多様であるのに対応させて,葉のかたちがさまざまに分化しているほか,常緑性と落葉性の樹木の差は葉の越冬法(または乾期に対する耐え方)の差によっているし,多年生草本のうち,夏緑性と常緑性の差も基本的には樹木の場合と同様である。海浜や乾燥地帯で多肉の葉が多かったり,渓流沿いで流線型の葉が進化してきたりするのも,環境に対する適応が,生活機能の最も高い葉に現れている例である。森林の階層構造も,個々の種の樹形も,光合成の効率を高めるように進化してきたものであり,葉の位置や構造が重要な役割を果たしている。それにもかかわらず,日常用語では,重要なことが〈根幹〉であり,〈枝葉末節〉の問題はとり上げるに足らぬこととされている。これは植物体を静的にみて,葉は中心から最も外れた存在であると解釈したもので,植物からみれば心外な表現かもしれない。
植物と人間とのかかわり合いのうち,葉が生活と触れ合う部分は大きく,衣食住のすべてにかかわっている。植物の季節変化は花や実と同様葉が表現しており,若菜摘み,紅葉狩り,落葉たきをはじめ,柏餅,草餅,早苗,ささ舟と,葉に関するものをあげればきりがない。最近では都会で緑の乏しい生活環境が多くなったために,観葉植物のブームといわれ,人間の潜在的な緑への欲求が,葉との触れ合いを求めることではからずも表に出てきているともいえる。
執筆者:岩槻 邦男


 〈ヨウ〉
〈ヨウ〉 〈ショウ〉梵語の音訳字。「
〈ショウ〉梵語の音訳字。「 〈は(ば)〉「葉陰・葉巻/青葉・枝葉・草葉・言葉」
〈は(ば)〉「葉陰・葉巻/青葉・枝葉・草葉・言葉」 [名]木の葉の
[名]木の葉の [接尾]助数詞。
[接尾]助数詞。




 (よう)。
(よう)。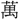
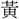
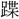 dyapは声近く、
dyapは声近く、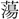
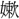 葉・敗葉・貝葉・病葉・風葉・楓葉・歩葉・末葉・万葉・木葉・門葉・来葉・落葉・乱葉・蘭葉・柳葉・緑葉・累葉
葉・敗葉・貝葉・病葉・風葉・楓葉・歩葉・末葉・万葉・木葉・門葉・来葉・落葉・乱葉・蘭葉・柳葉・緑葉・累葉