デジタル大辞泉 「感覚」の意味・読み・例文・類語
かん‐かく【感覚】
2 美醜やよしあし、相違などを感じとる心の働き。センス。感受性。「日本人の
3 (他の名詞の下に付いて)
㋐それを感じ取る心の働き。「バランス
㋑そういう心構えや雰囲気。気分。「学生のアルバイト
[類語](1)知覚・官能・五感・体感・肉感・感触・感じ・感・センス・錯覚・幻覚/(2)感性・感受性・美感・美意識・神経・センス・センシビリティー・フィーリング
Sponserd by ![]()
翻訳|sensation
Sponserd by ![]()
一八世紀末、江戸の蘭学者による造語か。後に幕末、明治初期の英和辞典では、Sensation, Sense の翻訳語として用いられるようになる。
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
感覚器官に加えられる外的および内的刺激によって引き起こされる意識現象のこと。
仏教用語としては古くから眼識,耳識,鼻識,舌識,身識(これらを生じさせる五つの器官を五根と称する)などの語が用いられたが,それらを総称する感覚という言葉はsensationの訳語として《慶応再版英和対訳辞書》に初めて見える。日常語としては坪内逍遥《当世書生気質》などに定着した用法が見られ,また西田幾多郎《善の研究》では知覚と並んで哲学用語としての位置を与えられている。
哲学史上では,エンペドクレスが感覚は外物から流出した微粒子が感覚器官の小孔から入って生ずるとしたのが知られる。それに対しアリストテレスは〈感覚能力〉を〈栄養能力〉と〈思考能力〉の間にある魂の能力の一つととらえ,それを〈事物の形相をその質料を捨象して受容する能力〉と考えた。一般にギリシア哲学では,感覚と知覚との区別はいまだ分明ではない。感覚が認識論の中で主題的に考察されるようになったのは,近世以降のことである。デカルトが方法的懐疑の途上で,感覚に由来する知識を人を欺きやすいものとして真っ先に退けたように,大陸合理論においては一般に感覚の認識上の役割は著しく軽視されている。カントにおいては,感覚は対象によって触発されて表象能力に生じた結果を意味するが,〈直観のない概念は空虚であり,概念のない直観は盲目である〉の一句に見られるように,彼は感性的直観と概念的思考の双方を重視した。他方イギリス経験論においては,感覚はあらゆる認識の究極の源泉として尊重され,その思想は〈感覚の中にあらかじめないものは知性の中にはない〉という原則に要約されている。ロックによればわれわれの心は白紙(タブラ・ラサtabula rasa)のようなものであり,そこに感覚および内省の作用によってさまざまな観念がかき込まれる。ここで感覚とは,感覚器官が外界の可感的事物から触発されることを通じて心に伝えるさまざまな情報のことである。また感覚の要素的性格は,〈単純観念〉がいっさいの知識の材料であるとする考えの中に表現されている。ロックの思想はバークリーおよびD.ヒュームによって受け継がれ,さらに19世紀の後半マッハを中心とする〈感覚主義〉の主張中にその後継者を見いだす。マッハは伝統的な物心二元論を排し,物理的でも心理的でもない中性的な〈感覚要素〉が世界を構成する究極の単位であると考えた。その思想は論理実証主義によって展開され,〈感覚与件理論〉として英米圏の哲学に浸透した。〈感覚与件sense-datum〉の語はアメリカの哲学者J.ロイスに由来し,いっさいの解釈や判断を排した瞬時的な直接経験を意味する。代表的な論者にはB.A.W.ラッセルおよびG.E.ムーアがおり,そのテーゼは事物に関する命題はすべて感覚与件に関する命題に還元可能である,と要約される。マッハに始まるこれら現代経験論の思想は,要素心理学や連合心理学の知見,およびそれらの基礎にある恒常仮定(刺激と感覚との間の1対1対応を主張する)とも合致するため,19世紀後半から20世紀初頭にかけて大きな影響力をもった。
しかし20世紀に入ってドイツにゲシュタルト心理学が興り,ブントに代表される感覚に関する要素主義(原子論)を批判して,われわれの経験は要素的感覚の総和には還元できない有機的全体構造をもつことを明らかにした。メルロー・ポンティはゲシュタルト心理学を基礎に知覚の現象学的分析を行い,要素的経験ではなく〈地の上の図〉として一まとまりの意味を担った知覚こそがわれわれの経験の最も基本的な単位であることを提唱し,要素主義や連合主義を退けた。また後期のウィトゲンシュタインは,言語分析を通じて視覚経験の中にある〈として見るseeing as〉という解釈的契機を重視し,視覚経験を要素的感覚のモザイクとして説明する感覚与件理論の虚構性を批判した。このように現代哲学においては,合理論と経験論とを問わず,純粋な感覚なるものは分析のつごう上抽象された仮説的存在にすぎないとし,意味をもった知覚こそ経験の直接所与であると考える方向が有力である。いわば認識の構造を無意味な感覚と純粋の思考という両極から説明するのではなく,両者の接点である知覚の中に認識の豊饒(ほうじよう)な基盤を見いだそうとしているといえよう。日本では近年,中村雄二郎が個々の特殊感覚を統合する〈共通感覚〉の復権を説いて話題を呼んだ。
→意識 →感覚論 →知覚
執筆者:野家 啓一
われわれの体には,内部環境や外部環境の変化を検出するための装置がある。この装置を受容器という。受容器を備えて特別に分化した器官が感覚器官である。内・外環境の変化が十分大きいと,受容器は反応し,次いでそれに接続した求心神経繊維に活動電位が発生するが,これを神経インパルスあるいは単にインパルスという。求心繊維を通るインパルスは脊髄あるいは脳幹を上行し,大脳皮質の感覚野に到達する。普通,生理学的には,感覚は〈感覚野の興奮の結果生ずる,直接的・即時的意識経験〉と定義される。これらのいくつかの感覚が組み合わされ,ある程度過去の経験や記憶と照合され,行動的意味が加味されるとき知覚が成立する。さらに判断や推理が加わって刺激が具体的意味のあるものとして把握されるとき認知という。例えば,われわれが本に触れたとき,何かにさわったなと意識するのが感覚であり,その表面がすべすべしているとか,かたいとかいった性質を感じ分ける働きが知覚であり,さらにそれが,四角なもので,分厚く,手に持てるといった性質や過去の同種の経験と照合して本であると認知されるのである。受容器から出発して感覚野に至るインパルスの通る経路を感覚の伝導路という。受容器,伝導路および感覚野によって一つの感覚系が構成される。環境の中のいろいろな要因のうち,受容器に反応を引き起こすものを感覚刺激といい,特定の受容器に最も効率よく反応を引き起こす感覚刺激をその受容器の適当刺激adequate stimulusという。例えば眼(感覚器官)の光受容器は,電磁波のうち,400~700nmの波長帯域すなわち光にのみ反応する。このことから受容器は多数の可能な感覚刺激の中から特定のものを選び分けて,その情報をインパルス系列にコード化し,中枢神経系に送る一種のフィルターとして働くとも考えることができる。大脳皮質に達した神経インパルスは,ここで処理され,その情報内容が分析され,さらにいろいろな受容器からの情報と組み合わされて,総合的情報が形成され,それが感覚野の興奮に連なるのである。
受容器を適当刺激の種類により分類すると,表1のようになる。またシェリントンCharles Scott Sherrington(1857-1952)は,受容器と刺激の関係から受容器を外部受容器exteroceptor(体外からの刺激に反応する受容器)と内部受容器interoceptor(身体内部からの刺激に反応する)とに分けた(1926)。前者は,さらに遠隔受容器teleceptor(身体より遠く離れたところから発せられる刺激に反応するもの,視覚,聴覚,嗅覚の受容器)と接触受容器tangoceptor(味覚や皮膚粘膜にある受容器)に,後者は固有受容器proprioceptor(筋肉,腱関節,迷路などの身体の位置や,四肢の運動の受容器)と内臓受容器visceroceptor(内臓にある受容器)に分けた。このような受容器の相違に基づき感覚は種modalityに類別される。古くから五感といわれた視覚,聴覚,触覚,味覚,嗅覚のみならず,平衡感覚,温覚,冷覚,振動感覚,痛覚なども種である。さらに同じ感覚種内でも個々の受容器の特性の違いから起こる感覚の内容の違いを質qualityという(表1)。例えば視覚では,受容器として杆(状)体,錐(状)体の2種類がある。杆体の働きにより明・暗の感覚が,錐体の興奮により赤,黄,緑,青といった色づきの感覚が生ずる。これらを質というのである。表2に臨床的感覚の分類を示す。視覚や聴覚のように受容器から大脳皮質まで判然とした形態学的実体をもったものと,そうでないものという観点から,前者を特殊感覚,後者を体性-内臓感覚とするものである。
感覚の生理学的研究方法には,主観的方法と客観的方法とがある。主観的方法では刺激とそれによって引き起こされる被検者の感覚の大きさを被検者自身が評価するもので,精神物理学的方法ともいわれる。客観的方法は主として神経生理学的方法によるもので,例えば微小電極をしかるべき感覚系の特定の部位に刺入し,個々のニューロンのインパルス反応を記録することにより,感覚の神経機序を研究対象とする。最近では,行動科学的手法による感覚の研究も行われている。これはオペラント条件づけの方法を用いて,感覚刺激とそれによって引き起こされる行動の変化を観察,計測するものである。例えば視覚でよく知られている暗順応の時間経過をハトを使って行った実験が有名である。ハトに,刺激光を見たときにAのキーをつっつき,刺激光が見えないときBのキーをつっつくようオペラント条件づけの方法で学習させる。ハトを明るいところから暗いスキナー箱に入れ,目の刺激光を点灯する。ハトは刺激光が見えるのでAをつっつく。すると刺激光はしだいに暗くなっていき,ハトは見えなくなるまでAをつっつく。刺激光が見えなくなってはじめてハトはBをつっつき,見えるまでBをつっつき続ける。ハトはAとBのキーを操作することによって刺激閾(いき)を決定するわけである。このようにして時間的に刺激閾が低下する,いわゆる暗順応曲線がハト自身の行動によって描かれるのである。
受容器(具体的に細胞を指すときは受容器細胞または感覚細胞という)はそれ自身がニューロンであって,軸索が第一次求心繊維として働くものと,それ自身は上皮細胞に由来する非ニューロン性細胞で,これに感覚ニューロンがシナプス結合しているものとある。前者を一次感覚細胞(例,嗅細胞),後者を二次感覚細胞(例,内耳の有毛細胞)という。
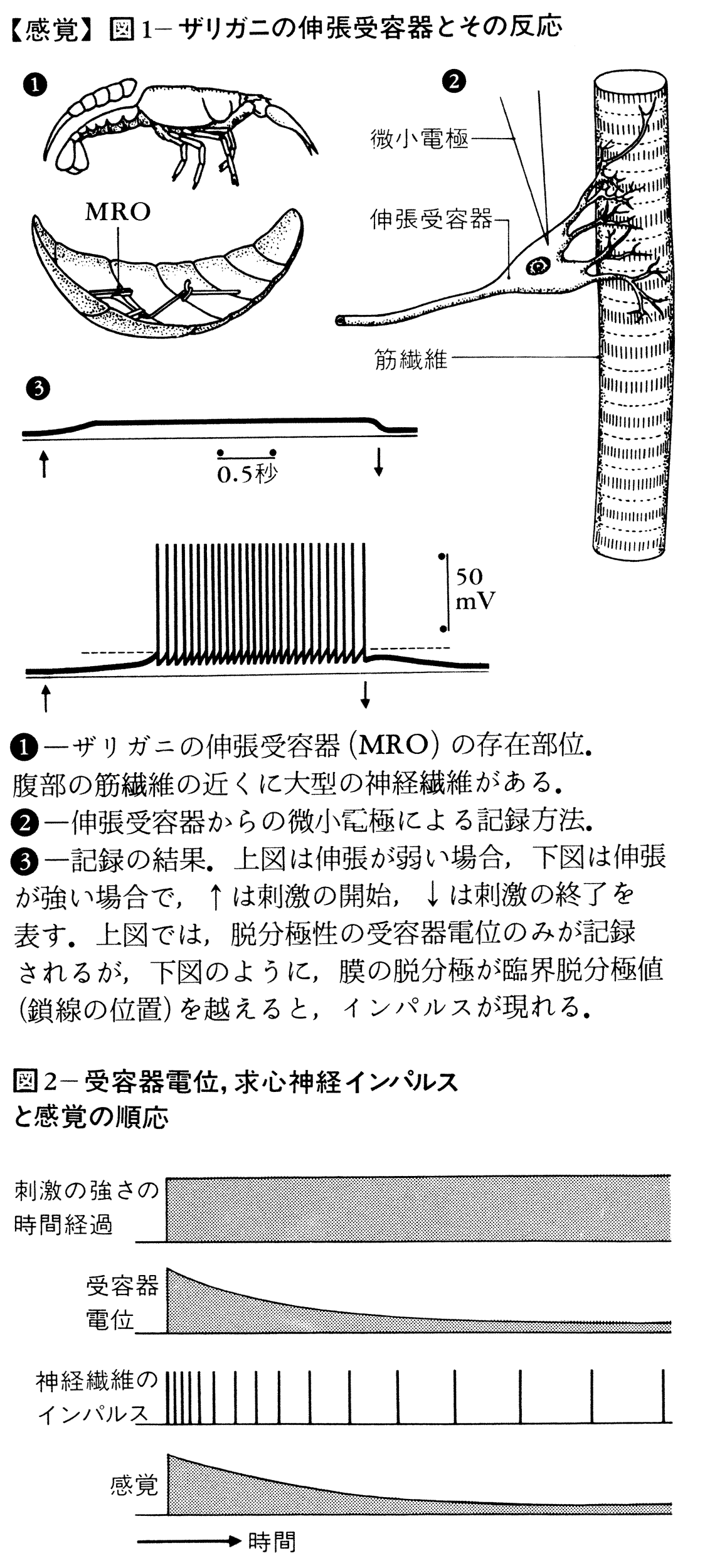
感覚の受容機構を甲殻類の伸張受容器を例にして簡単に説明しよう(図1)。この受容器細胞は大型の神経細胞で筋繊維の近くに存在する。細胞体からでる樹状突起dendriteが筋繊維の表面にくっついており,筋繊維が伸ばされると,樹状突起も引っ張られ変形を受ける。このとき細胞の膜電位は脱分極を示す。この脱分極の大きさは伸長が大きくなればなるほど大きくなるという性質をもつ(この性質をもつ反応を段階反応graded responseという)。脱分極がある一定の大きさを超えると,このニューロンの軸索に全か無かの法則によってインパルスが発生し,軸索を中枢に向かって伝わる。インパルスの頻度は受容器電位の振幅と直線関係をもつ。内耳の有毛細胞では,機械的刺激によって毛が屈曲するとき膜電位が変化するが,動毛側への屈曲で脱分極,不動毛側への屈曲で過分極が生ずる。脱分極性の受容器電位の場合には,有毛細胞からその振幅に相応した量の化学伝達物質がシナプス間隙(かんげき)に放出され,この伝達物質の作用を受けて求心繊維の終末が脱分極する。このシナプス後電位の大きさが十分大きいとき,求心繊維にインパルスが生ずる。一次感覚ニューロンでみた受容器電位は,直接インパルスを発生させる原因になるところから起動電位generator potentialともいわれる。一次求心繊維の放電頻度の時間経過をみると,一定の大きさの刺激を持続的に与えているにもかかわらず,しだいに低下してくる。この現象を順応adaptationという。これに相当する現象はすでに受容器電位(または起動電位)にも起こっていることが確かめられている(図2)。順応の速い受容器を速順応性quickly adapting(略してQA),遅いものを遅順応性slowly adapting(略してSA)という。感覚にみられる順応現象がすでに受容器で起こっていることを示すものである(もちろん,感覚の順応には受容器の順応のみでは説明できない部分がある)。
個々の感覚はいくつかの基本的特性(属性)によって規定される。質,強さ(大きさともいう),広がり(面積作用)および持続(作用時間)の四つが主要なものである。
(1)感覚の大きさ 一つの感覚系について,感覚刺激の強さを十分弱いところからしだいに増していくと,やっと感覚の生ずる強さに達する。感覚が生ずる最小の刺激の強さを,その感覚の刺激閾(絶対閾)という。またある強さIとI+⊿Iが識別できる最小の強さの差⊿Iを強さに関する識別閾という。この場合,⊿I/Iの比を相対刺激閾という。この比がそれぞれの感覚について,ある刺激の強さの範囲内でほぼ一定であることがE.H.ウェーバーによって見いだされた。この比をウェーバー比Weber ratioという。この比の値はだいたい次のようである。光の強さ1/62,手で持った重さ1/53,音の強さ1/11,塩の味1/5。絶対閾は,光覚で10⁻8μW,音の強さ10⁻10μW/cm2(このとき鼓膜を10⁻9cm足らず動かすにすぎない)などである。感覚の大きさと,刺激の強さの関係を示す式として,ウェーバー=フェヒナーの式とスティーブンスS.S.Stevensが提唱したスティーブンスのべき関数が知られている。感覚の大きさをR,刺激の強さをI,刺激閾をI0とすると,
R=K log I+C
(ウェーバー=フェヒナーの式)
R=K(I-I0)n
(スティーブンスのべき関数)
ともにKとCは定数である。スティーブンスのべき指数nの値は暗順応眼の点光源の明るさについては0.5,砂糖の甘味1.3,腕の冷覚1.0,圧覚1.1などである。中耳の手術の際に鼓索神経からインパルスを記録し,味覚刺激の濃度とインパルス頻度の関係を求めたところ,主観的計測で求められたのと同じnの値をもつべき関数が得られた。感覚神経から記録されるインパルスについては,〈刺激の強さが増すにつれてインパルス頻度が増し,また放電活動する繊維の数も増す〉ことが知られている。これをエードリアンの法則Adrian's lawという。
(2)感覚の空間的特性 感覚は大脳皮質感覚野の興奮に起因する現象であるが,このときわれわれは感覚刺激が外界の,あるいは身体の一定の場所に与えられたものと判断する。これを感覚の投射projectionという。感覚のこの性質によって刺激の位置および部位を定めることができる。この性質は,受容器の存在する受容面と感覚野との間に整然とした場所対場所の結合関係が存在するからである。このことを感覚野に部位再現topographic representation(皮膚感覚の場合には体部位再現somatotopy,視覚の場合には視野再現visuotopyまたは網膜部位再現retinotopy)があるという。ある強さの刺激が感覚を起こすためには,ある広さ以上の面積を刺激する必要がある。この面積を面積閾といい,ある面積以内では刺激の強さIと面積閾Aとの間にI×A=一定の関係が成り立つ(これをリッコーの法則Ricco's lawという)。同一種の刺激を二つの異なった2点に与えた場合,2点を分離して感ずることができる。しかし2点間の距離を小さくしていくと,ついには2点を2点として区別できなくなる。弁別しうる2点間の最小の距離を2点弁別閾または空間閾という。
(3)感覚の時間的特性 刺激が感覚を起こすのには,ある一定時間以上受容器に作用しなければいけない。この最小作用時間を時間閾という。例えば光の感覚では,光の強さIと時間閾Tとの間には,ある時間範囲内においてI×T=一定の関係が成り立つ。これは光化学反応におけるブンゼン=ロスコーの法則に相当するものである。閾上の感覚刺激を与えても,その強さに相当する大きさの感覚が生ずるまでには,ある時間の経過が必要である。すなわち感覚はしだいに増大(漸増という)する。また刺激を止めたときも,もとの状態に復帰するまで感覚は漸減する。刺激を止めた後に残る感覚が残感覚aftersensationで,その性質が初めの感覚と同じ場合,陽性残感覚,反対のとき陰性残感覚という。同じ刺激を反復して与えるとき,その周期が十分短いとき,個々の感覚は融合して,ある一定の大きさの連続した感覚となる。例えば点滅する光を見たとき,その点滅の周期が十分短いと,もはや点滅の感覚はなく,連続した一様な明るさの光として感じられる。この現象の起こる最小の点滅頻度を臨界融合頻度critical fusion frequency(略してCFF)という。
(4)感覚の感受性の変化 同じ刺激を続けて同じ受容器に与えているとき,感覚の大きさは順応によってしだいに低下していく。触覚は順応の速い感覚である。身体を動かさない限り,着衣の感覚が失われるのはこの性質による。このほか,感覚にみられる特殊な現象に対比contrastといわれる現象がある。例えば一定の明るさの灰白色の小さい紙面の感覚的明るさは,その紙を黒い大きな紙の上に置くときより明るく(白く)見えるし,もっと白い紙の上に置くときは暗く見える。この現象を同時または空間対比simultaneous or spatial contrastという。灰白色の紙が大きいときは,黒い紙と接する部分が中央の部分よりより白く見えるし,また白い紙と接する場合はより黒く見える。この現象を辺縁対比border contrastという。また,白い紙を見て次に黒い紙を見ると黒い紙はいっそう黒く見え,黒い紙を見て次に白い紙を見ると白い紙はいっそう白く見える。この現象は継時または時間対比successive or temporal contrastといわれる。
微小電極を感覚系のいろいろな部位に刺入して,ニューロンの活動を記録するという方法(微小電極法)の導入により,神経系が感覚情報を符号化(コード化)する機構についての研究がひじょうに進歩した。研究成果のなかで最も重要な発見は受容野ということである。例を視覚にとろう。1本の視神経繊維からインパルスを記録する。繊維により光で網膜を照射すると,インパルス頻度が増すものと,逆に減り,光を消したとき増すもの,および照射の開始と終了時に一過性に頻度を増すものがある。第1のような反応をON反応,次のものをOFF反応,最後のものをON-OFF反応という。照射面積を直径100μmくらいに小さくすると,網膜の特定の範囲を照射したときのみしか反応しない。この範囲はほぼ直径1mmくらいである。このように一個の感覚系ニューロンの放電に影響を与える末梢受容器の占める領域を,そのニューロンの受容野receptive fieldという。ネコやサルの視神経繊維(または網膜神経節細胞)の受容野は,ON領域とOFF領域が同心円状に配列した構造をしている。中心部がON領域でそれを取り巻く領域がOFF領域である受容野をON中心OFF周辺型,これと逆の配列をしているものをOFF中心ON周辺型という。一般に受容野の中心部と周辺部とは互いにその作用を打ち消し合うように働くため,受容野全体を覆う光刺激に対しては反応は弱く,中心部のみを照射するときは最も強い反応が得られる。このような中心部と周辺部の拮抗作用は網膜の神経網内に側抑制または周辺抑制の機構が存在することによるもので,辺縁対比の神経機構と考えられる。視覚系では脳幹の中継核である外側膝状体のニューロンの受容野も視神経繊維のものと本質的には同じものであるが,大脳皮質の第一次視覚野ではニューロンの受容野の性質は一変する。すなわち,視覚野ニューロンの受容野は一般に方形状で,長軸方向に伸びた細長いON領域とOFF領域から構成されている。したがって受容野全体を覆う光に対しては,皮質ニューロンはまったく反応しない。細長いON領域のみを覆う線状の光に対して最大の反応を示す。つまり,このような受容野をもつ皮質ニューロンは,受容野の軸の方位に一致し,受容野のON領域のみを覆うスリット状の光に選択的に反応するという特性をもっているということができる。このような方位選択性が皮質ニューロンに共通にみられる性質である。皮質ニューロンの受容野は,ON領域とOFF領域がはっきりわかるもの(単純型)ばかりでなく,これらの領域がはっきりしない複雑型,さらに受容野の両端に抑制帯がある超複雑型が区別される。いずれにしても皮質ニューロンは,自分の受容野の性質に従って,特定の条件に合う刺激を選択する性質をもっている(これを特徴抽出機能という)。視覚野が行ったこのような分析結果は,さらに高位の皮質中枢(連合野)に転送され,視覚情報の異なった側面についての分析と統合が異なった部位でなされている(分業体制)らしいことが,最近の研究により明らかになりつつある。サルの上側頭溝にある皮質ではヒトやサルの顔に特異的に反応するニューロンのあることが報告されており,また19野の一部では特定の色に選択的に反応するニューロンのあることが報告されている。他の感覚についても,皮質の感覚野では感覚刺激の特徴抽出を行うニューロンのあることが報告されている。オペラント条件づけの方法と微小電極法を駆使することにより,最近は感覚よりはむしろ知覚についての神経機構を解明すべく努力がなされている。
→神経系
執筆者:小川 哲朗
体の外部または内部から与えられた刺激を受容して興奮し,その興奮を中枢神経系側(求心側)に伝える器官を感覚器官という。一般に多数の受容器の集合よりなる。感覚器官は,適当刺激を選択したり,刺激を効率よく感覚細胞に伝えるのにつごうがよい構造をしていたり,そのための付属装置をもつ。例えば目のレンズや虹彩,耳の鼓膜や耳小骨などがこれに相当する。単純に見える昆虫の感覚子でも,クチクラ装置は,受容される刺激の種類によりひじょうに異なる。例えば嗅感覚子ではにおい分子がクチクラを通過するための嗅孔が数多くクチクラ壁に見られるが,味感覚子では味溶液は通常一つの味孔により感覚細胞の受容部と接触している。
感覚器の刺激受容部には,一般に感覚細胞と支持細胞が見られるが,ときには感覚細胞の興奮を求心側に伝えていく二次神経細胞や三次神経細胞が存在することもある。また,脊椎動物の味蕾(みらい)や嗅上皮のように,将来,感覚細胞に分化する基底細胞があることもある。
感覚器官は,感覚の種類によって視覚器,聴覚器,味覚器,嗅覚器,平衡器,圧覚器,触覚器,痛覚器,温覚器,冷覚器,自己受容器などと呼ばれることもあるが,感覚器官が受容できる適当刺激によって分類されることもある。適当刺激により分類すると光感覚器,機械感覚器,化学感覚器,温度感覚器,湿度感覚器,電気感覚器などに分類できるが,さらに細分された場合には,例えば振動感覚器などと呼ばれることもある。適当刺激による感覚器の分類は,とくに,ヒトには見られず動物に特有な感覚器,例えば電気感覚器や赤外線感覚器,あるいは水生無脊椎動物の化学感覚器などを扱うときにつごうがよい。動物には磁気感覚をもつものもあると報告されているが,磁気感覚器は見つかっていない。また,感覚器官には,検知する対象が体から離れた遠い所にある遠隔感覚器と体表に接して起こる事象に関する接触感覚器の区別もある。前者には視覚器,聴覚器,嗅覚器などが含まれ,後者には皮膚感覚器や味覚器が含まれる。
感覚器官の活動を知る指標として,感覚器官全体の電気的活動が用いられることがある。例えば網膜電図は目を光刺激したときに網膜に発生する電位変化を記録したもので,光刺激により最初に現れる電位変化は,脊椎動物では角膜側が負,無脊椎動物では正の波として現れ,感覚細胞の受容器電位の集合と考えられている。嗅粘膜をにおいで刺激したときに発生する電位を記録したものは嗅電図,昆虫の触角をにおいで刺激したときに発生する電位を記録したものは触角電図と呼び,においの有効性の検知などのために使われる。しかし,これらの電位変化は多くの種類の細胞の活動の集合であるので,感覚器官内の特定の細胞の活動を調べるためには微小電極法などの別の手段による観察が必要となる。
執筆者:立田 栄光
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
光や音のような生体外の事象とか、生体組織のゆがみや体内の化学物質といった生体内の事象によって直接おこされた意識過程を感覚という。これに対し、感覚を素材として、さらに記憶とか推理とかの高次の神経作用を加えて、刺激源、あるいは対象物に関する表象が構成されるとき、これを知覚perceptionといい、幾種類かの知覚から刺激源や対象物がなんであるかを知ることを認知cognitionという。たとえば、ある波長の電磁波が目を刺激し「青い」という単純な意識が生じたとき、これを感覚といい、これが木の葉の青さであって海の青さでないと具体的に判断されるとき、これを知覚という。さらに、同時に加えられたいくつかの感覚刺激によって、それは眼前の庭木の青さであると認めるとき、これを認知という。このように、いちおう知覚と認知とは、感覚を介して生ずる一段高次の精神現象とされているが、実際には、三者の間、とくに感覚と知覚との間に判然とした区別をつけることはむずかしい。
[市岡正道]
感覚は次の三つの働きによって生ずる。
(1)感覚受容器 感覚刺激を受け入れる受容器で、物理的あるいは化学的エネルギーの刺激を感覚神経の一連の活動電位、すなわち感覚信号に変換する器官である。つまり、受容器は一種のエネルギー変換器といえる。
(2)感覚神経 受容器に加えられたアナログ性(計量性)の感覚刺激は、受容器でデジタル化(計数化)され、神経インパルスに変換される。この感覚信号は、感覚神経系中をさまざまな修飾を受けながら求心性に伝達される。
(3)大脳皮質感覚領 感覚神経インパルスが感覚に変換されるところである。しかし、その変換のメカニズムは、現在のところ解明されていない。
[市岡正道]
形態学的な見地からの一つの分類法として、感覚は次のように分けられる。
〔Ⅰ〕特殊感覚……視覚、聴覚、味覚、嗅覚(きゅうかく)、平衡感覚
〔Ⅱ〕体性感覚
(1)皮膚感覚……動き受容感覚(触覚、圧覚、振動感覚)、温度感覚、痛覚
(2)深部感覚……深部圧覚、運動感覚、振動感覚、深部痛覚
〔Ⅲ〕内臓感覚……狭義の内臓感覚(一般感覚ともいう)、内臓痛覚
視覚や味覚のように、比較的判然とした構成と機能とをもった感覚を特殊感覚special sensationという。これに対し、内臓臓器のように、自律神経系の分布を受けている器官で生ずる感覚を内臓感覚visceral sensationといい、その他の感覚が体性感覚somatic sensationとなる。
体性感覚のなかの触覚、圧覚、振動感覚は、いずれも皮膚や皮下組織の変形、あるいはゆがみといった、いわば組織の動きが刺激となっておこるものである。これらの感覚は、単に順応(後述)の遅速が違うだけにすぎない同一次元の感覚であるため、一括して動き受容感覚とされる。この感覚の受容器は、皮膚ではマイスネルMeissner触覚小体様の神経終末であり、皮下ではファテル‐パチニVater-Pacini小体(被覆性神経終末)である。
深部感覚とは、皮膚と内臓との中間にある筋、腱(けん)、骨膜などからおこる感覚をいう。このうち運動感覚は、筋紡錘や腱紡錘中の張力受容器からの求心性情報によっておこる感覚で、四肢の相対的位置関係、四肢の運動、物体の重量や抵抗を感ずるものである。
[市岡正道]
感覚がもつ一般的な性質には次のようなものがある。
(1)特殊感覚エネルギーの法則 眼球を圧迫しても、視神経を電気刺激しても、目に光が作用したときと同じように光の感覚がおこる。このように、受容器から大脳皮質感覚領までの感覚系統において、そのどの部分に、どのような種類の刺激を作用させても、その感覚系統に固有な特定の感覚が生ずる。これを「特殊感覚エネルギーの法則」といい、ドイツの生理学者ミュラーJ. P. Müllerによって提唱された。ミュラーは、「各感覚系統はそれぞれ固有の活動を営む」と考えたのであった。
(2)適当刺激 受容器は各種の刺激で興奮するが、もっとも敏感に反応する特別な種類の刺激があり、これを適当刺激という。視細胞に対する光刺激などがその一例である。
(3)感覚の投射 身体に加えられた感覚刺激は、大脳皮質感覚領に興奮をおこし、感覚が生ずるわけであるが、その感覚は脳内のできごととして感じられるのではなく、外界の対象物によって生じたもの、あるいは刺激の加えられた身体部位に生じたものとして感じられる。これが感覚の投射であり、投射によって刺激の加わった部位を知ることを定位という。たとえば、左手第2指指背に物が触れたとすると、触覚は大脳皮質体性感覚領に生ずるが、実際には身体当該部位に物が触れたと感ずるなどである。
(4)刺激の強弱と時間 刺激が弱すぎれば感覚はおこらないが、弱い刺激でも反復して与えられると、ついには感覚が生ずることがある(潜伏加重)。また、感覚は刺激を与えても、すぐに生ずるものではなく、徐々におこってくるものであり(漸増)、刺激をやめても即時に感覚がなくなるものではなく、やはり、徐々に消えてゆくものである(漸減)。さらに、同一の刺激が持続的に与えられていると、感覚はしだいに鈍くなっていく(順応)。感覚のうちには、触覚、嗅覚のように順応の速いものと、痛覚や深部感覚のように順応の遅いものとがある。順応の遅い感覚は、刺激が加わっている間はほぼ同じ強さで感じられる。眼鏡や帽子の着用は時間の経過とともに意識しなくなるが、靴の中の小石による痛みはいつまでも続く。これは前者が順応の速い触覚によるものであり、後者が順応の遅い痛覚によるためである。
(5)ウェーバー‐フェヒナーの法則 二つの刺激を同時に、あるいは順次に与えたとき、認知できる両者の最小差を「弁別閾(べんべついき)」という。ドイツのE・H・ウェーバーは、一方の刺激の強さをR、弁別閾をΔRとすると、ΔR/Rは、Rが中等度の強さであれば、ほぼ一定であることをみいだした。これを「ウェーバーの法則」といい、ΔR/R=Cをウェーバー比(Cは定数)という。たとえば、100グラムと110グラムとが、また500グラムと550グラムとが弁別できたとすると、ΔR/Rは両者において10%となり、一定であるということになる。その後、ドイツのフェヒナーは、この法則から出発して、感覚の大きさEは刺激の強さRの対数に比例する(E=k1logR)ことを理論的に求めた(k1は定数)。これをウェーバー‐フェヒナーの法則という。この精神物理的な法則は、以前から、考え方の出発点において難点があると批判されてきたが、近年アメリカの心理学者S・S・スティーブンスは、感覚の大きさEと刺激の強さRとの間には、E=k2Rnの関係があることを実験的に求めた(k2, nは定数)。これを「べき関数の法則」という。
(6)感覚の対比 感覚は、同時に、あるいは順次に与えられた刺激によって、その性質や大きさが変わる。これを対比という。たとえば、同じ灰色の紙が、黒地の上では白く、白地の上では黒く見えるなどは同時的対比とよび、ある味を味わったあとに蒸留水を口にすると、別の味を呈するなどは継時的対比とよぶ。
[市岡正道]
動物が体の内外の環境変化(刺激)に対応しながら、自己の生活機能をもっとも有利な状態に維持していくためには、絶えず体の内外の刺激を検出していなければならない。このように体の内外の環境の変化を刺激として感知する機能を感覚といい、これにあずかる器官を感覚器とよぶ。動物は環境の変化、すなわち種々の刺激である物理的または化学的エネルギーをまずは体の一部で受容し、それに対して適切な反応をする。したがって動物の感覚は、受容器の種類による分類以外に、これらのエネルギーすなわち適当刺激の種類によっても分類される。刺激が化学物質である場合は化学感覚といわれ、味覚と嗅覚とがある。また、その刺激を受容する場所を化学受容器という。一方、物理刺激によるものは物理感覚とよばれ、それはさらに機械受容(音受容、振動受容、平衡受容、自己受容)、温度受容、光受容などに分類される。そして、これら各種の刺激をもっとも効果的に受容し、情報として中枢に伝えるためには、発達した受容器が必要となる。この受容器によって受容された刺激が感覚として脳で意識されるまでには、何段階かの神経過程を経なければならない。感覚が神経情報として脳内で処理され、さらに適応行動として発現するまでの過程を、感覚情報処理という。現在の生理学と行動生物学の面から、感覚情報として理解されている感覚について次に述べる。
[青木 清]
(1)味の感覚には酸・甘・苦・塩の四基本型があるが、これら四つの基本の味に特異的に応じる受容器はない。しかし、動物のなかでヒトが甘味にいちばん敏感なことなどが知られている。また、脳では味を認識できるが、ヒトが感じる味の複雑微妙な感覚のメカニズムについてはわかっていない。
(2)音の感覚のうちで、意味をもつ情報としては、ヒトの声によることばの認識と同種間動物のコミュニケーションとがある。これらは、聴覚求心経路を経て聴覚領野に情報が送られることによって生ずる。
(3)においの感覚は、鼻腔(びくう)粘膜のにおい受容細胞で受け取られたのち、大脳の前頭葉嗅領野まで送られて生ずる。しかし、においのどの要素がどのように送られるかは、まだわかっていない。
(4)視覚情報による感覚は、視細胞、双極細胞、水平細胞、アマクリン細胞などといった網膜内で複雑に絡み合った細胞群で連絡されたのち、視神経節細胞を経て視神経交差後に外側膝状体(しつじょうたい)に行く。しかし両眼視を行わない動物では、中脳視蓋(しがい)が視覚感覚の最高位中枢となっている。哺乳(ほにゅう)類以上では、大脳皮質の視覚領の17、18、19野が視覚の最高位中枢となっている。
(5)触覚、温度覚、痛覚などの皮膚感覚は、後索核を経て視床腹側基底核群に入り、大脳皮質体性感覚野に情報として伝わる。この感覚野が最高中枢である。
[青木 清]
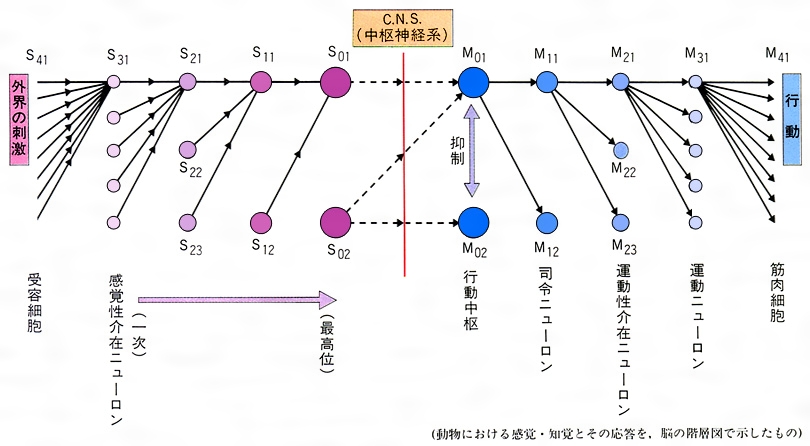
各種感覚がどこで生ずるかを述べてきたが、感覚における最高中枢の機構がよく調べられているのは、アメリカの生理学者ヒューベルとT・N・ウィーゼルが研究した視覚に関してである。彼らは大脳皮質のニューロンレベルの研究のなかで、視覚ニューロンには、視界の特定の位置を検出する二つの異なるニューロン層があることを発見した。その二つとは、単純型検出ニューロン、およびさらに上位のニューロンで、後者は、どの位置にあっても同方向の黒線であれば反応する複雑型検出ニューロンである。この2種のニューロンの研究により彼らは、感覚としての階層性の存在を示し、さらに視覚を例として感覚が知覚され行動として発現する中枢の階層性モデルを示すことができた。この感覚情報処理機構を階層性モデルで示すとのようになる。
この図に示されるSは感覚系をさしており、Mは運動系をさしている。S41は視覚、聴覚、嗅覚、味覚などの感覚受容細胞層を示す。視覚を例にすると、網膜から大脳皮質の視覚領野がS41、S31、S21、S11、S01としだいに上位の中枢へとあがることを示している。S22、S23は一連の感覚情報系への制御として働くニューロンを示す。またS01は、たとえばカエルがハエを視界中に収めていて、ハエが動きだしたという感覚をおこすニューロンである。このS01の活性化が摂食のための最高中枢M01を活性化し、さらにそのM01がM11を刺激してカエルはハエの方向に向き照準を定めることになる。これを定位するといって、動物が感覚し知覚したことを意味する。
[青木 清]
従来から、動物の感覚器の精度が異なれば、それに伴いそれぞれの動物にとっての世界は異なり、環境とは主体的なものであるという環境世界説があった。両者の研究の結果、さらに、ほとんど同じ感覚器をもっていても、その後の情報処理において刺激を検出するニューロンが異なることで、環境世界が異なるということがわかったわけである。したがって、動物において本能行動としてみられる感覚としての種特異性の多くは、感覚系の種特異的検出ニューロンの存在、およびその働きに由来するといえる。
このような感覚の例には次のようなものがある。まずミツバチのダンスであるが、これは「垂直方向を太陽方向に読み替える」ことの表現である。すなわち、視覚刺激と体表の感覚毛の機械的刺激との両方を読むニューロンが中枢にあって、明るい方向と重力の方向を一つのニューロンが検出している結果であると考えられる。またフクロウが音を聞くのは、音源の方向を高い精度で検出する能力をもつニューロンの働きによる。しかしそれとは別に、大脳皮質のほかの領野には視覚と聴覚とに応答するニューロンが存在していて、ここでは一つのニューロンが同じ方向からくる光と音との両方に応答することになる。これらの例から、二つ以上の異種感覚系からの情報を受け取る感覚認識ニューロンの存在が知られる。
[青木 清]
植物は一般に刺激の受容・伝達・認識という、動物生理学的な過程を経る特別の器官をもたない(とくに認識のための器官を欠く)ので、厳密な意味での感覚はない。しかし、植物には刺激を受容する部分があり、感受された刺激は伝達され、特定の部分で反応がみられる。したがって、植物における感覚とは、外界の刺激の受容と同義に扱ってよいであろう。以下、こうした意味からの植物の感覚について述べる。
[勝見允行]
植物は重力に反応して姿勢を変える。その代表的な例が屈地性である。こうした重力刺激の受容部位は根や茎の場合、その先端である。根では一般に先端数ミリメートルのところまで、マカラスムギの子葉鞘(しようしょう)では先端約10ミリメートルのところまでが感受部位である。重力刺激に対する感度は、植物の種類、部分、発育段階、外界の諸条件によって異なる。反応を得るために必要な最少刺激時間(刺激閾時)は、根では20分前後、茎では5~10分が普通である。ゼンマイの成熟葉はとくに速く30秒である。重力刺激の受容機構はまだ明らかではないが、スタトリスstatolith説(平衡石(へいこうせき)説ともいう)による説明がある。重力刺激感受部位の細胞内にはスタトリスとよばれる、動きやすく、重いアミロプラスト(デンプン粒)があり、細胞の位置が変わると、アミロプラストは重力方向に動き、細胞内のいちばん低い場所に集まる。その場所での細胞膜との接触刺激で重力を感知するという考えである。現在のところ、この説を支持する事実、否定する事実の両方があって、いずれかは決めがたい。
[勝見允行]
植物は、屈光性、傾光性というように、光に対して反応を示す。子葉鞘や茎の屈光現象の場合、光を感受する部位は先端部である。マカラスムギの子葉鞘では先端50マイクロメートルがもっとも鋭敏である。傾光性や横日(おうじつ)性の場合には葉が光感受部位となる。
屈光現象をおこすのに必要な光の強さは、植物によっても、観察条件によっても異なるが、一定の大きさの屈曲をおこす光の刺激は、光度ではなく光量によって決められる。
光刺激は光受容物質によって感受される。マカラスムギの子葉鞘の場合、青色光がもっとも有効であり、作用スペクトルから、カロチンまたはフラビン系統の物質が関与するのではないかと考えられている。ヒザリオ(緑藻類)の一種の細胞に含まれる平らな葉緑体では、その面が光と平行になるように移動する。この場合、光受容物質はフィトクロムである。また、オジギソウやネムノキなどの傾光性運動でもフィトクロムが関係している。
[勝見允行]
ある種の植物は屈触性、傾触性、傾震性のように接触、あるいは震動、風圧などの刺激に反応する。食虫植物の捕虫葉には接触刺激を受容する特別な器官がある。ムジナモやハエジゴクの捕虫葉の内側には感覚毛があって、刺激を与えると基部で折れ曲がり、その部分にある特別な細胞が変形する。傾性における刺激の受容体については、まだ明らかにされていない。感覚毛の変化や、オジギソウの傾震運動は、刺激受容後にきわめて短時間で活動電位が伝わるので、受容体は活動電位の発生に関係したものとも考えられる。
[勝見允行]
チューリップ、サフラン、クロッカスなどの花は温度差に敏感に反応して開閉運動を行う。サフランの花は0.2℃の温度上昇で開き始める。
花粉管やカビの菌糸は化学的刺激に反応し屈化性を示すし、食虫植物の捕虫葉は傾化性を示す。
[勝見允行]
『問田直幹他編『新生理学 上巻』第5版(1982・医学書院)』▽『W・F・ギャノング著、岡田泰伸他訳『医科生理学展望』原書20版(2002・丸善)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
出典 最新 心理学事典最新 心理学事典について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
字通「感」の項目を見る。
出典 平凡社「普及版 字通」普及版 字通について 情報
Sponserd by ![]()
Sponserd by ![]()
…いっさいの認識は感覚のみに由来すると主張するか,それとも感覚がいっさいの認識の必要,かつ十分な条件であると主張する哲学的立場。sensualisme(感覚論)という用語は19世紀初頭以来,フランスで使われており,フランスの《アカデミー辞典》には,1878年版から採録されている。…
…晩年はボジャンシーの近くの田園に隠棲した。哲学的主著には,《人間認識の起源に関する試論》(1746)と《感覚論》(1754)がある。前者ではロックの学説を継承し,人間の認識の起源として感覚と反省の二つを認める立場をとった。…
…perceptionという語は,〈完全に〉〈すっかり〉などの意を示す接頭辞perと,〈つかむ〉を意味するラテン語capereとからなる語であり(ドイツのWahrnehmungは,〈注意〉の意を有するwahr――英語のawareなどに残っている――と,〈取る,解する〉を意味するnehmenとからなっている),たいていは五感によって〈気づく〉〈わかる〉ことを意味する。哲学や心理学でも,感覚を介する外的対象の把握が普通に〈知覚〉と呼ばれている。したがって,それは,純粋に知的な思考や推理とは区別されるが,また単なる感覚とも区別されるのが普通である。…
…人間に固有の思考力,認識力は一般に〈知性intellect〉ないし〈理性〉と呼ばれ,古来,規則に従って分析し論証する〈悟性understanding〉,原理・始元を直覚・洞察して総観し統括する〈理性reason〉の二面を含むとされる。本能,感覚,記憶,想像,意志とは区別され,また啓示や信仰に対置されてきた。 理性という訳語は,事物の本性を示す仏教用語〈理性(りしよう)〉および〈道理〉とともに,1881年(明治14)の《哲学字彙》でreasonに当てられた(1870年西周(にしあまね)はreasonを人間に備わる〈性の智〉,86年中江兆民はフランス語のraisonを〈良智〉と訳した)。…
※「感覚」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
福岡県福岡市博多区の櫛田神社の夏祭り。壮麗な山笠で知られる。今日,山笠には飾り山笠と舁き山笠(かきやまがさ)の 2種類がある。明治時代に電線が架設されて以降,物語場面の人形などを飾りつけた高さ 15m...