デジタル大辞泉 「植物」の意味・読み・例文・類語
しょく‐ぶつ【植物】
[類語]
うえ‐もの〔うゑ‐〕【植(え)物】
2 畑に植える野菜類。
3 連歌・俳諧の題材となる事物の分類の一つで、草木・苔などの植物。
4 近世、軍略上、城郭内に植えた樹木。
Sponserd by ![]()
翻訳|plant
Sponserd by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponserd by ![]()
生物界を動物と植物に二大別するのは,常識の範囲では当然のように思えるが,厳密な区別をしようとするとさまざまな問題がでてくる。かつては生物の世界を動物界と植物界に二大別するのが常識だったが,菌類を第三の界と認識すると,それに対応するのは狭義の動物(後生動物),狭義の植物(陸上植物)ということになり,原生動物や多くの藻類などは原生生物という名でひとまとめにされ,また,これら真核生物に比して,細菌類やラン藻類は原核性で,原核生物と別の群にまとめることができる。そこで,生物の世界を,動物界,植物界,菌界,原生生物界,原核生物界(モネラ界)の5界に大別する5界説が提議され(R.H. ホイッタカー,1969)広く使われているが,これは生物の世界をわかりやすく整理したもので,いわゆる人為分類の体系である。
本項でいう植物は動物と対応させる広義のものであるが,藻類,菌類についてはそれぞれの項で分類に言及されているので,本項の分類の記述では,主として狭義の陸上植物について詳述する。
動物と植物の区別はヨーロッパで古くから確立しており,アリストテレスは生物界を植物phytōn,動物zōon,人間anthrōposに三大別した。この当時は,大型の後生動物と,陸上植物や一部の大型藻類,菌類だけしか知られていなかったので,動くか動かないかで動植物を区別することも可能であった。しかし,生物には顕微鏡的な微小なものもあることが知られるようになった今日では,動物か植物かという区別だけでは割り切れない生物も多くみつかっており,現に,動物分類表にも植物分類表にも挙げられるものが少なくない。それでも一般に,大昔からの動物,植物の区分が便宜的に通用しているのは,この,どちらかといえば人為的な区別に便利な点が多いからであろう。日本では動物,植物の概念は近世までなく,植物の代りに,草,竹,木,花などのことばが使われていた。
動物と植物の差を簡単に定義することは難しい。動物は従属栄養で,自然界の物質循環の消費者,分解者の地位を占めるが,植物のうちでも,菌類は葉緑素をもたずに従属栄養の生活をしており,基本的には分解者である。被子植物のうちにも,ギンリョウソウやツチトリモチのように,二次的に腐生あるいは寄生生活をするようになったものがあり,すべてが生産者であるとはいいきれない。ミドリムシはふつうの環境では葉緑体をもっていて独立栄養の生活をしているが,暗所で培養すると葉緑体をつくらず,養分を与えられると従属栄養の生活を続ける。さらに,同じ独立栄養でも,葉緑素をもっていて炭酸同化をする光独立栄養のほか,一部の細菌類のような化学同化を行うものの化学的独立栄養などは,見かけ上の類似はあっても,系統的に同質のものとみなすのは困難である。
細胞壁の有無は動植物の差の一つであるが,単細胞藻類には細胞壁をもたないものが多い。また,同じ細胞壁でも,陸上植物や緑藻類ではセルロースなどを基質としたものであるが,褐藻類や紅藻類ではフコイジン,ラミナリン,寒天,キチン質などでできており,簡単に同質と断じることはできない。さらに,生活環のある時期,とくに生殖細胞には細胞壁はない。動くか動かないかも動植物の差とはいえず,動物でも,海綿動物や腔腸動物などには固着生活をするものが多く,植物とされていても,クラミドモナスやミドリムシは運動性をもっている。植物の運動の例に,膨圧の差による運動を行うオジギソウが挙げられるが,これは筋肉や鞭毛による運動とは異質のものである。
いろいろの形質を指標としても,植物を容易に定義することができないのは上に述べてきたとおりであるが,このことも,生物が単元的な起源をもつものか,いくつかの群が起源を異にするものか,まだ結論の出せない理由の一つである。だから,系統分類学の観点から,生物界の大分けをどう定義すればよいかは,客観的に示すことができない。
ここでは,菌類や,原生動物の植物性鞭毛虫類に入れられることのある単細胞藻類も含めて,広義の植物について以下に記したい。ウイルスやリケッチアも植物分類表の付録につけられることもあるが,それぞれの項目に任せることにして,ここでは一応除外しておく。
ミリミクロンの単位の大きさのウイルスを除外しても,植物の大きさの変異には目をみはるものがあり,小さいものはミクロン単位の細菌類から,大きいものはジャイアント・セコイアのように高さは100mを超え,径が数mに達するものまである。1世代の長さにしても,数十分で分裂を繰り返して新しい個体をつくる細菌の例から,屋久島の縄文杉のように6000年の樹齢を誇るものまである。形の複雑多様なことについては,いちいち触れることもできないほどであり,それぞれ生活場所の多様さと複雑な相関関係を示している。植物がこのように多様に分化してきたのは,いうまでもなく,40億年に近い進化の歴史を通じて,さまざまの環境に適応してきたからである。
植物の起源や系統についてはよくわかっていないのだから,系統分類の体系はできていない。しかし,これまでの研究で,光合成色素,光合成の最終産物,細胞壁の成分,鞭毛の性質などを手がかりにすると光独立栄養の植物の大綱分類ができることがわかっており,
菌類もそれに対応させることによって植物の分類系に組み込むことができる。
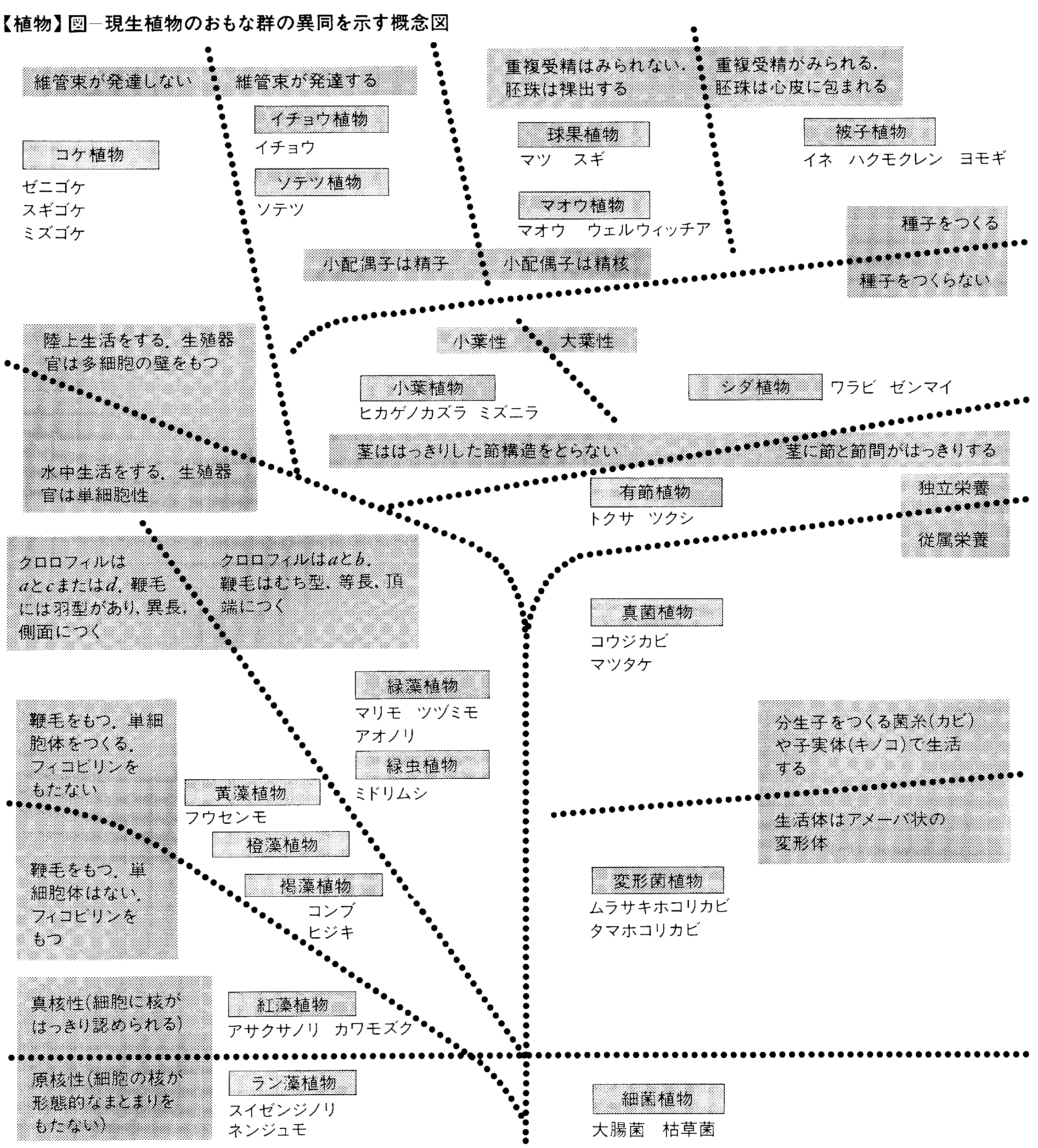
以下に19の門からなる植物の分類系について,門ごとに短い解説をつけるが,この分類系には,藻類,菌類,シダ植物,裸子植物,種子植物などという門は出てこない。これらのグルーピングは系統の違いをもとにしたものではなくて,むしろ進化の段階を示す人為的なまとまりといえよう。しかし,門の名まえではなくても,この人為的な群の名まえや範囲が便利なことがあるので,現在でもしばしば使われることがある。
(1)細菌植物門 単細胞の菌類で,簡単な群体をつくることもある。核膜に包まれた核はなく,DNAは細胞内に散在している。有性生殖は知られておらず,増殖はもっぱら二分裂による。細胞が球形,棒状,らせん形などになり,鞭毛をもつものがある。この鞭毛は真核生物のもつものとは構造は異なっている。生物の死体など,有機物の分解に重要な役割を果たしているものと,病原菌など生物に有害なものなどがある。化学合成を行うものや,光合成バクテリアのように,独立栄養のものもある。約150属1500種。
(2)ラン(藍)藻植物門 単細胞体か,それが寄り集まった群体で,淡水または海水中に生活しており,地衣を構成する藻ともなる。細胞内では核や色素体がまとまった構造はとらず,核酸が中心部に,葉緑素が周辺部に散在している。有性生殖は行わず,細胞分裂で増殖する。ラン藻素,紅藻素などのフィコビリンをもち,同化産物はデキストリン。約160属1400種。細菌とラン藻では核膜で包まれた構造の整った核はつくられない。このようなものを原核生物と呼び,構造のはっきりした核をもっている真核生物と区別することがある。
原生生物界には原生動物のほか卵菌類,サカゲツボカビ類,変形菌類など菌界に属さない広義の菌も含まれるが,かつて植物とされていたもののうちでは,藻類が原生生物界に含められる。藻類の分類は藻類の項に記されているが,主要な門について述べると,
(3)紅藻植物門 多細胞体ではあるが,体制は基本的には糸状体であり,それがもつれあって寒天質に包みこまれ,外見は複雑な構造をもつようにみえる。生活史は複雑であるが,生殖細胞にも鞭毛はない。ラン藻素,紅藻素などのフィコビリンをもち,同化産物は紅藻デンプン。海産,まれに淡水産。約500属4000種。
(4)黄藻植物門 単細胞体,群体,糸状体,多核の囊状体などで,細胞壁は2片が重なり合うことが多い。運動性のある細胞の鞭毛は1本か,不等長の2~3本,うち1本は羽型である。鞭毛で運動するものは原生動物鞭毛虫類に分類されることもある。クロロフィル(葉緑素)はaとcで,同化産物は油かロイコシン。海産,淡水産のほか,陸上の湿地にも生じる。約300属1万種あり,その多くはケイ藻類である。
(5)橙藻植物門 炎藻植物ともいう。単細胞体,群体,糸状体で,運動性のある細胞には不等長2本の鞭毛があり,うち1本は羽型。運動性のあるものは原生動物鞭毛虫類に分類されることもある。クロロフィルはaとc,同化産物は油,デンプンなど。有性生殖はまだ確かめられていない。海産プランクトンとして重要で,淡水にも生育する。約150属1000種。
(6)褐藻植物門 多細胞の大型藻類が多く,キサントフィルを多量に含んでおり,褐色を呈する。クロロフィルはaとcで,同化産物はフコーサンなど,ラミナリン,マンニット,油脂の形で貯蔵される。生殖細胞のうち,運動性をもつものでは不等長の2本の鞭毛をもち,うち1本は羽型。淡水産の数種を除いて海産で,寒い地域に多い。約240属1500種。黄藻,橙藻,褐藻の3群は,鞭毛の構成やクロロフィルaとcをもつ性質などで共通しており,藻類のうちではまとまった一群である。
(7)緑虫植物門 ミドリムシ植物ともいう。鞭毛をもった単細胞体で,細胞壁はない。クロロフィルはaとb,同化産物はパラミロンと油脂,有性生殖は知られておらず,増殖はもっぱら二分裂による。淡水産。原生動物鞭毛虫類に分類されることもある。約25属400種。
(8)緑藻植物門 単細胞体から群体,糸状体,葉状体,多核体など,体制は多様である。単細胞体や群体で,鞭毛をもって運動するものは,原生動物鞭毛虫類とされることもある。運動性をもつ細胞の鞭毛は等長の2本または4本,まれに多数,いずれも尾型。クロロフィルはaとbで,同化産物はデンプン。淡水産,海産,湿った陸上にも生活し,土壌藻や気生藻もある。約500属5000種。
以下に述べる陸上植物は,すべてクロロフィルaとbをもち,同化産物としてデンプンをつくり,鞭毛がある場合には等長で尾型のものをもつ点などで共通しており,その性質は緑藻のものと同じであることから,陸上植物は緑藻から進化したものと信じられている。
(9)コケ植物門 陸上,まれに水中に生活し,樹幹,岩上に着生することが多い。生活史の主相は配偶体で,胞子体はその上に寄生的に付着する。精子の鞭毛は2本。器官,組織の分化は乏しい。約900属2万5000種。蘚類と苔類の起源が別であるとみなして,別の門に分類する意見もある。
(10)小葉植物門 現生のものは陸生で小型の草本。生活史の主相は胞子体で,根,茎,葉の分化していないものもある。配偶体は独立の生活をしているが目だたない。精子の鞭毛は2本。茎は原生中心柱をもち,葉のあるものでは葉脈が単生するのがふつうである。根,茎,葉が分化していないマツバランの類を独立の門とする考えもある。古生代から中生代にかけて栄えた群で,現生は5~7属約1000種。維管束植物は単元的に進化してきたと考えられていたが,葉の系統発生などに指標されるように,小葉植物は大葉性の植物と別の系統に属することがわかってきた。植物が陸上に進出してきたのは約4億年前のシルル紀のことである。
(11)有節植物門 生活史の主相は胞子体で,節と節間のはっきりした茎をもつ。節に輪生する葉は楔葉(せつよう)とよばれる。精子の鞭毛は多数。配偶体は小さくて目だたず,胞子は同型胞子性。古生代に繁栄したが,現生は1属約25種。
(12)シダ類植物門 生活史の主相は胞子体で,根,茎,葉が分化する。葉は大葉,茎の中心柱は原生中心柱,管状中心柱,網状中心柱など。配偶体は小型で扁平,ハート型の前葉体になるものが多く,独立栄養を営む。精子は多数。現生種は約320属1万種。小葉植物,有節植物,シダ類植物を合わせてシダ植物という。無種子維管束植物というまとまりで認識されたもので,系統的な単位ではない。
以下の5門((13)~(17))がいわゆる種子植物である。配偶体が極端に退化して胞子体の胚珠の組織に寄生する。被子植物以外の4門が裸子植物であるが,これが多系的であることから,五つの群を独立の門とすることが多い。
(13)ソテツ植物門 茎は太く,軟木質。胞子囊は葉緑につき,配偶体は胚珠内で発達する。精子には鞭毛がらせん状に配列する。子葉は2枚。古生代から中生代にかけて繁栄した群で,現生は9~10属約100種。
(14)イチョウ植物門 茎はよく分枝する高木となり,材は硬木質。胞子囊は短枝の葉腋(ようえき)につき,配偶体は胚珠内で発達する。精子には鞭毛がらせん状に配列する。胚乳(雌性配偶体)には葉緑体がある。古生代から中生代にかけて繁栄した群で,現生はイチョウ1種。
(15)球果植物門 茎がよく発達した木本で,真正中心柱をもち,材は硬木質。葉は針状のことが多く,胞子囊は軸に密集してつき,胞子囊穂をつくる。鞭毛をもつ生殖細胞はない。子葉は2枚から数枚。古生代から中生代にかけて繁栄し,現生は約40属600種。
(16)マオウ植物門 茎の二次組織に道管が発達する。胞子囊穂は分枝し,胞子囊托は花被に包まれる。胚囊(雌性配偶体)は退化し,花粉管受精を行う。子葉は2枚,系統関係のよくわからない3属よりなる。
(17)被子植物門 草本,木本など生活型は多様である。維管束には道管があり,葉では脈が連結する。配偶体は著しく単純化し,胚囊(雌性配偶体)は7核,花粉管(雄性配偶体)は3核となる。2個の精核が卵細胞と中心核に受精する重複受精がみられ,胚と胚乳がつくられる。子葉は1~2枚。ジュラ紀末に出現し,現在地球上で最も優勢な植物群で,約1万2000属20万種以上に多様化している。
(18)変形菌植物門 光合成色素をもたず,従属栄養の生活をする。固形物をとりこんで細胞内消化を行うものが多い。栄養生活相と生殖相は区別され,アメーバ集合体と胞子の形成が特徴的である。不等長の2本の鞭毛をもつものもある。原生動物偽足虫類に分類されることもある。約100属500種。
(19)真菌植物門 カビやキノコの類で,寄生や腐生などの従属栄養の生活をしている。生活史は複雑なものが多く,鞭毛も1~2本,羽型と尾型がある。有性生殖の知られていないものをひっくるめて不完全菌類という。子囊菌類と担子菌類はよくまとまった群であるが,接合菌,藻菌,鞭毛菌などといわれるものの分類については不明の点が多い。約5000属4万5000種,菌類の起源については,藻類が二次的に従属栄養になったとする説と藻類とは起源を異にするという説とがある。真菌類は内容が多様な群であるので,いくつかの門に細分すべきだという意見もある。
このほか地衣類は,藻類と菌類の共生体であるが,よくまとまった共生の植物体をもっているので,単一の分類群として扱われることがある。約400属2万種。
地球上に生命が発生したのは三十数億年前とされるが,それから長い間細菌やラン藻のような原核生物の時代が続いた。真核生物が進化してくるのはおよそ15億年前と推定されているが,真核生物が単系的なものかどうかはまだ確かめられていない。細胞の共生によって真核生物が出現したという説もある。
ラン藻,紅藻,ケイ藻,褐藻,ミドリムシ藻,緑藻などが藻類である。クロロフィルはaだけでフィコビリンをもち,鞭毛をもって動く細胞をつくらないラン藻,紅藻と,クロロフィルはaとcで,鞭毛には羽型のものがあるケイ藻,褐藻と,クロロフィルはaとbで,鞭毛は複数あっても等長ですべて尾型のミドリムシ藻,緑藻と,藻類には三つの群があるが,これが系統的な差かどうかについては明らかでない点も多い。
コケ植物とシダ植物の造卵器は壁が多細胞の頸卵器であるが,種子植物にはそれが単純化した構造があり,その中で胚の形成がみられる。そのことから,陸上植物を有胚植物ということもある。また,陸上植物を茎葉植物とよんで藻類,菌類の葉状植物と対比させることもあるが,苔類なども葉状植物といって定義がはっきりしない。陸上植物が緑藻類から約4億年前に進化してきたことはほぼ確かなようである。
地衣類は藻類と菌類の共生体であるが,菌類の部分によって分類されるので,菌類といっしょに扱われる。細菌が変形菌,真菌とどのような系統関係をもつかは問題である。菌類は生物界における第三の界を構成するという考えは,真菌は細菌から進化してきたものであるという前提に立っている。菌類が紅藻や黄藻から進化してきたという意見が正しいとすれば,菌類の進化の歴史は15億年以内だということになる。
菌類以外の藻類,陸上植物をひっくるめて有色植物という。菌類を第三の界とした場合の植物がこれに相当し,二次的にクロロフィルを失ったものを別にすれば,独立栄養の生活を営むものである。
後生動物はひじょうに多様な生物群であるが,個体発生の過程で,胞胚から囊胚をつくり,胚葉を分化して組織や器官を形成するので,基本的な構造で共通している。それに対して,植物では,同じ多細胞体でも,柔組織をつくるもの,偽柔組織をもつもの,多核体のものなど,異なる系統群で体制が平行的に進化しており,形に共通のまとまりがみられない。維管束植物は比較的まとまった群であるが,それでも,葉についてみればすべてが相同な器官とは断言できない。系統発生をみれば,小葉植物の葉が茎の表面の隆起によってできたものであるのに対して,それ以外の葉はテロム枝(初期の陸上植物にみられる二叉(にさ)分枝をする枝系)の変形でできた可能性が強いからである。いわんや,コケ植物の葉とよばれているものは,配偶体上に分化したものであり,藻類には陸上植物の葉と相同の器官は認められない。それでも,植物の形を記述する場合には,記載用語を使うのが便利なので,系統発生上の形態の相同性はあまり気にしないで,例えば,葉的器官とよぶべきような葉と相似の構造をも,すべて葉と記載するような簡便法がとられることが多い。
植物の細胞は細胞壁をもつ点で動物のものと区別されるが,ミドリムシ類のように細胞壁をもたない細胞からだけなる植物門もある。しかし,精子などのように鞭毛をもって運動する植物細胞には細胞壁がないのがふつうであり,運動性のある単細胞体はすべて動物であると定義することも難しい。植物細胞のもう一つの特徴は葉緑素をもっていることであるが,この性質も菌類にはあてはまらない。ギンリョウソウやツチトリモチのように,腐生生活や寄生生活をし従属栄養となったものでは葉緑体をもたないが,これはむしろ二次的にそうなったということが推定される。それに対して,菌類の起源については,藻類が従属栄養化したものであるという説と,有色植物とは別に起源してきたものであるという説とがあって,まだはっきりとはわかっていない。また,葉緑体をもつ細胞のほとんどは基本組織系のもので,孔辺細胞以外の種子植物の表皮組織系のように,葉緑体をもたないものも多い。また,根のように,光のあたらない部分の細胞にも葉緑体はつくられない。
植物の組織が,分裂組織と永存組織に大別できることも動物とは違った特徴である。分裂組織の位置や働き方によって植物の体制はさまざまな形をとる。生長の様式には,頂端生長,辺縁生長,節間生長,頂毛生長などいろいろあるが,維管束植物では,茎頂と根端における一次生長と,形成層による二次生長がはっきりしている。生長点の細胞(群)は,植物体が全体としてどれだけ年をとっても,つねに胚的な性質を失わず,新しい組織を形成する。屋久島の縄文杉は樹齢6000年といわれるが,この個体でも,生長点のところは胚と同じように若々しい活動を続けている。また,永存組織となっているものでも,カルスのように,再び分裂能を取り戻すこともあり,植物体は個体全体が老齢化するというよりは,組織ごとの自立性の強いものである。
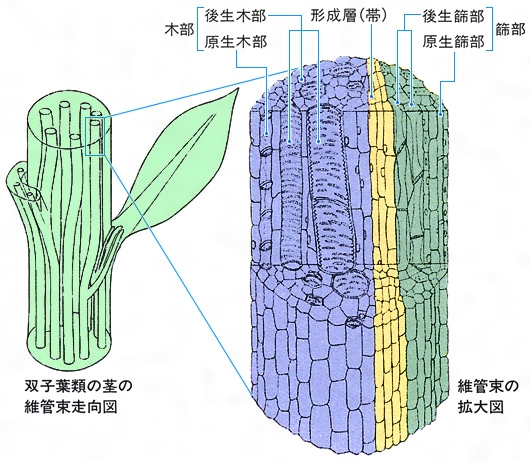
維管束植物では,一次生長によって,表皮組織系,基本組織系,維管束系が分化し,さらに多様な組織がつくられてくるが,藻類や菌類では組織の分化は乏しい。大型藻類やキノコ類では,形や機能のよく似た細胞の集りを組織とよんではいるが,他の組織との間の差が明確でない場合も多い。
植物の器官としては根,茎,葉,花が挙げられるが,これらの器官が分化しているのは維管束植物だけで,花については被子植物でよくまとまった構造となっている。藻類や菌類にも,根,茎,葉に相当する見かけ上の構造の分化がみられることがあり,生殖細胞が特殊な構造となることもあるが,器官とよべるほどまとまった構造をとることはない。
植物には核相が単相の細胞でつくられる配偶体の世代と,複相の細胞でつくられる胞子体の世代があり,配偶体につくられる生殖細胞の配偶子は合体して接合子となり,胞子体では減数分裂の結果核相が単相の胞子をつくる。このため,配偶体の世代と胞子体の世代が交互に現れる世代交代がみられる。後生動物では生活環は単純で,成体に卵と精子がつくられ,受精が行われると胚発生を経て成体となる。多様に分化した動物のうちには,この生活環にいろいろの例外を生じることもあるが,基本的な道筋は一つであるといえる。植物では,上に述べた二つの世代が一つの生活環をつくっており,その点で後生動物と異なっている。二つの世代が規則正しい世代交代をみせている植物もあるが,それぞれの世代にさまざまの例外や変形を生じたり,一方が省略されて,単相の植物体だけになったもの(アオミドロなど)や,逆に後生動物と同じように複相の植物体だけになったもの(ヒジキ,ミルなど)もある。植物の多様化の過程には,生活史を多様化させることによって獲得されてきた性質も数多い。
維管束植物では世代交代は規則的である。コケ植物では,ふつうに目にとまる植物体は配偶体で,卵と精子をつくる。受精卵は配偶体上で生長し,胞子体が配偶体に寄生するような姿となる。胞子体は短い1本の棒であるが,その先端に胞子囊を1個つくり,胞子を散布する。シダ植物段階のものでは,ふつうに目につくのは胞子体で,胞子をつくって散布する。胞子は発芽して配偶体となるが,扁平で緑色の前葉体とよばれる構造をとるものと,塊状になって地下性で菌根をもつものとがある。配偶体で受精を行った卵と精子が配偶子に相当する。受精卵は胚発生を行って胞子体をつくるが,若い胞子体は配偶体上に生じ,やがて独立できる大きさになると,親であった配偶体を押しつぶしてどんどん生長していく。種子は若い胞子体が種皮に包み込まれて休眠している状態のものであるが,種子をつくる植物体では,胞子葉に生じた胚珠中で胚囊母細胞(胞子母細胞)が減数分裂を行って胚囊細胞(大胞子)をつくり,それは細胞分裂(発芽)して胚囊(雌性配偶体)となる。一方,小胞子葉にあたるおしべには花粉囊(小胞子囊)中で花粉母細胞(胞子母細胞)が減数分裂をして花粉(小胞子)をつくるが,花粉はすでにその構造のまま細胞分裂を行っており,単細胞の生殖細胞と配偶体との中間的な性質ももっている。花粉は柱頭で発芽して花粉管(雄性配偶体)となり,精子または精核(雄性配偶子)をつくる。受精が行われると,受精卵は発生を行って若い胞子体をつくり,それが休眠して種子となる。すなわち,種子植物の体は胞子体であり,配偶体は花のめしべの組織にみえるほど極端に単純化し,その配偶体上に次世代の胞子体が形成されることになる。配偶体が単純化しているが,胞子体の世代と配偶体の世代は,核相もそれぞれに対応して,ひじょうにはっきりしており,規則正しく交互に現れている。
事実上,植物は地球上のあらゆる場所に住んでいる。熱帯降雨林のように,光が強く湿度も高いところが植物の生活には最適らしく,そういう場所では,種数も個体数も多く,植物社会が最もよく発達している。日本列島も黒潮暖流の影響で緯度のわりに気温が高く,雨量が多いうえ,地形が複雑であることなどもあって,植物の多様度はきわめて高い。生育している植物の種数も多く,植物群落もよく発達している。維管束植物が繁茂すれば,コケ,地衣なども多様となり,植物社会の構造と機能はますます複雑になってくる。植物相の多様さは,土壌菌や藻類にも及んでくることになるが,これらについては日本でもまだ十分研究されているとはいい難い。
地球上で植物社会の発達の悪いところといえば砂漠や極地である。しかし,そういう場所でも植物がまったく生育していないのは限られた部分だけで,それぞれに特殊化した植物の生存が確認されている。南極大陸には,コケや地衣をはじめ,藻類,菌類の生育も確かめられている。海の生物がどれだけ深いところまで生きているか,洞穴などを含めて地中にどのように生物が生活しているかなどについては,まだまだ未知な部分もあるが,動物の死骸などの有機物のある洞穴には菌類が必ずといってもよいほど生育しており,これらのことから,およそ生育のための条件がわずかでも備わっておれば,植物は地球上のあらゆるところに生活場所を広げているといえる。
植物が分布圏を拡大するのには,分類群によってさまざまなやり方が確立している。種子についてみても,空中を飛散するもの,鳥や獣に付着して運ばれるもの,はじけたり舞い下りたりして移動するもの,潮に流されて運ばれるものなど,さまざまである。胞子は小さい場合が多く,空に舞い上がると気流に乗って相当の距離を運ばれるようである。小型の藻類の場合,土といっしょに固められて鳥の脚などに付着して運ばれるということも知られている。このように,植物によって生育場所を広げるやり方はいろいろあるけれども,生育可能だからといって無制限に広がることはない。実際に地球上に生育している植物の分布には生態的特異性とともに地域的な特異性がみられる。地形その他の障害によって,分布域の拡大が阻害されているために,分布の地域特異性が生じるのであるが,これを手がかりに地史的な植物相の発展のあとを推定することもできる。
現生の植物の地球上における分布のおおよそについては,ほとんどの場合,種子植物の分布に従って区系地理的な区分が設定される。胞子の飛散によって分布域の拡大を行っている植物群の分布は,種子植物の場合と必ずしも一致するとはかぎらないが,種子植物以外の植物群の分布をおもな資料として,地球上における植物の分布について詳細な比較がなされることはむしろ珍しい。
動物地理区に対比させて,地球上の植物の分布区系を全北植物区系界,旧熱帯植物区系界,新熱帯植物区系界,オーストラリア植物区系界,ケープ植物区系界,南極植物区系界に六大別しようとする意見が古くからあったが,最近では,グッドR.Goodの区分を基本とする37の植物区系を設定して,地球上の植物分布を整理しようという傾向が強い。これによると,日本列島の大部分は中国からヒマラヤにかけての地域と共通で,日華植物区系区Sino-Japanese regionとなり,北は南サハリンから南千島までを含むことになる。吐噶喇(とから)列島以南の南西諸島は,台湾やインドシナ,タイ,ミャンマーなどと東南アジア区系区Continental South East Asian regionとなり,小笠原諸島については日華区系に入るのかメラネシア・ミクロネシア区系に入るのか,明快にどちらかは決めかねるところであるが,グッドはメラネシア・ミクロネシア区系区Melanesia and Micronesia regionに入れている。
日本列島における植物の分布についてもいくつかの特徴がみられるが,分布域の類型には定説はない。多雪地帯の日本海側と太平洋側の分化,ソハヤキ要素,マキネシアなどは日本の植物区系を理解するために考えられた区分である。また,南千島とオホーツク地区との境界に宮部線,フォッサマグナ地区に牧野線,吐噶喇列島のところに渡瀬線,小笠原の南に細川線などが提唱されており,それぞれ分布域の境界とされている。
植物は地球上のあらゆるところに生活場所を占めているが,それぞれの環境条件に合わせて,形態も機能もひじょうに多様化させている。高山植物が矮生(わいせい)化したり,海浜植物が多肉化したりというのはよく知られた現象である。また,昆虫による受粉を行う種子植物では,花の色や香りに特徴があり,形態的にも訪花昆虫とともに進化してきたような例がある。そのような外見上に現れる性質だけでなく,植物のもっている性質には環境とのかかわりの深いものが多い。
植物の働きにも植物固有のいろいろのものがあるが,人間の生活にとってかかわりの深いのが光合成であろう。菌類以外の植物は葉緑体をもっていて炭酸同化の能力をもち,地球上の物質循環の基礎生産者としての役割を果たすとともに,地球上の生物にとって不可欠の遊離酸素の産生にもあずかっている。物質生産については,細菌類の一部にいわゆる化学合成を行うものがあるが,生物界で最も普遍的で重要なのは光合成であり,現在の生物界の繁栄は植物がクロロフィルをもつようになったことと無関係ではない。クロロフィルには構造の異なるaからeまでのタイプのものが知られているが,これらの差は一般に植物の系統の差と平行にもたらされたものと信じられている。しかし,光合成におけるC3回路とC4回路が,系統の分化とは独立にいろいろの群で同時に分化してきたように,クロロフィルの構造の差も,もっと機能的な差に関連するものなのかもしれない。
植物は現に地球上に生活していて,活発に進化しつつある。さらに,人間の文化が始まって以来,多くの栽培植物が育種されてきたし,森林の開発,農地化も行われてきた。今後さらに遺伝子組換え技術の発達によって,自然界で進化してきた植物とは違った新しい人工的な植物もつくり出されるに違いない。地球上で創造され,その世界で進化発展してきた植物が,同じような進化の歴史の末に高等な知能をもつようになってきた人間と,どのように共存していくかは,進化の歴史の新しいページに書き加えられることであろう。それを,よりよい未来の礎とするのは,人類に与えられた最重要の課題である。
→有用植物
執筆者:岩槻 邦男
世界各地でそれぞれの民族がさまざまのしかたで植物を神格化したり,シンボル化したりしている。しかし,そこにはおのずから一定の法則が働いている。すなわち,人間と植物との交渉のしかたが変わるにつれて,当然のことながら,人間の植物イメージも変わっていく。人間と植物の付合いは次のような3段階で変化してきたと考えられる。その第1は,人間がまだ栽培植物をもたず,もっぱら野生植物だけと付き合っていた段階である。第2は栽培植物の発見以後の段階,つまり農業段階である。そして第3は農業生産に余裕ができ,それゆえ,有用植物以外の植物つまり観賞用の植物の開発が行われるようになった段階である。
J.J.ルソーは《人間不平等起源論》で〈人間はオークの下でどんぐりを腹いっぱい食べ,食物を提供してくれたその同じ木の下を自分のねぐらとしていた。しかし小麦の出現とともに人類は堕落した〉と述べて原始生活を賛美したが,逆にボルテールは〈小麦を知ってしまっているわれわれを,再びどんぐりの時代へと連れもどすな〉ということわざを使って,時代に逆行する試みをいましめた(《哲学辞典》〈小麦〉の項)。また二宮尊徳は〈豊あしのふか野が原を田となして食をもとめて喰ふ楽しさ〉という歌を詠んで,葦原(あしはら)から稲田への移行をよしとした。このように,農業時代のシンボルである麦や稲は人間にとって有用な存在ではあるが,しかし,野生植物にくらべるとひ弱い存在である。とくに日本における稲は冷害に弱いし,干害にも弱い。それに対し,野生の木や草ははるかに強い。そこで農業段階に入った人々は,そうした野生の木や草を村里へ運びこみ,農耕作物がそれにあやかるように祈った。そうした野生の木は,古代ギリシアの祭りではオリーブであったりゲッケイジュであった。また,ゲルマン人たちの五月祭で森から切り出される木は,シラカバの枝であったりサンザシの花枝であったりし,それらはメー・ポールつまり〈五月柱〉と呼ばれた。そしてこのことは日本でもまったく同様であり,水口祭(みなくちまつり)では山からサカキや松,ヤマブキの花やツツジの花が切り出され,苗代田の水口に立てられた。
人間が森林や原野を開墾しつつあった時点においては,野生の木,そしてそこに宿る神々や精霊は,人間に対して敵意をもつ恐ろしい存在であった。しかし,いったん開墾が終わると,もはや周囲の森や山の木々や神々は敵であることをやめ,農作物の生長に力を貸す存在となったのである。しかしながら,農民はそれだけでは満足せず,森や山から樹霊を招来するばかりでなく,みずからの耕している大地の中に新しい神を設定する。例えば古代ギリシア人はヒュアキンティアHyakinthiaなる祭りを行ったが,この祭りは,アポロンの誤って投げた円盤があたって死んだ美少年ヒュアキントスを記念する春祭である。ところで,このヒュアキントスは実はヒアシンス,あるいは類似の植物を人格化,神格化したものであると考えられる。そして,この地中海地方の荒野に生えるヒアシンスは,多年生の球根植物であり,初夏に紫色の花を咲かせ,そののち地上部分を枯死させて生き延び,翌年そのおなじ球根からまた花を咲かせるのである。こうした植物学上の事実からみても明らかなように,ヒュアキンティアは,冬を逃れ,夏を迎えることを喜ぶ祭りであり,それは同時に死を逃れ復活を喜ぶ祭りであった。ヒュアキントスはそうした死と復活に対する農耕民の願望を体現し,実演してみせた。
このようにヒュアキントスは,農作物をはぐくむ大地に根ざす植物を神格化したものであるが,やはりなんといっても野生の植物である。ヒュアキントスの死と復活は農耕民が望むところであるが,そうした願望は次の時代では野生の植物でなくて,栽培植物をシンボルとして使うことによって象徴化される。そして,その栽培植物とは,西方の世界でいえば小麦だったのである。実際イエス・キリストは〈1粒の麦は地におちて死ななければ1粒のままである。しかし死ねば多くの実を結ぶ〉といった意味のことを説いている。また近世ヨーロッパの寓意画には,一方では農夫が麦まきを行っており,他方では死者が墓からむくむくと起き出てくるといったものがよく見かけられるが,これは麦粒の死と発芽というイメージと,人間の死と復活の思想とを二重映しにしたものということができる。そうした死-復活のキリスト教思想と平行するように,東洋では〈大死一番,絶後再蘇〉といった禅のスローガンがあるが,これもまた〈枯木花を生ず〉といったイメージで説明されているのである。
人間は農業生産が進んで生活にゆとりが生まれてくると,植物を美的観賞の対象として見るようになる。例えば,ヨーロッパでは初めヒナゲシは小麦畑に生える雑草として憎まれていたが,19世紀になってJ.ラスキンがその美しさを鼓吹した結果,今ではもっぱらその美しさだけがめでられるようになっている。日本でも同様で,いけばなや四季の花を盛りこんだ歳時記,花暦などは,農業との直接の結びつきからは切り離された伝統の中にあるといってよい。花をそれ自体として楽しむ態度はもっぱら農事に携わらない都会人によって培われたということも,あわせて指摘しておきたい。
→木 →草 →花
執筆者:山下 正男
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponserd by ![]()
地球上の生物は動物、植物、菌類に大別されるが、植物とは、細胞壁を有し、独立栄養で光合成を行うことができる生物をいう。動物は細胞壁をもたず、従属栄養で栄養源は植物に依存しているし、菌類は細胞壁をもつが、もっぱら動植物に寄生し、生活のすべてをこれらの生物に依存している。このようにみると、植物は生物の栄養源として重要な役割を果たしているということができる。
[金井弘夫]
植物の特徴は、細胞膜の外に細胞壁を生じることのほかに、無機物からエネルギー源となる有機物を合成する能力をもつことである。細胞壁は体制の簡単な植物体の保護に役だち、乾燥から守り、とくに陸上植物では植物体を支えるために働いている。陸上植物の細胞壁はおもに多糖類のセルロースであるが、藻類ではマンナン、グルカン、アルギニン酸、寒天質などの細胞壁をもち、セルロースを欠くものもある。光合成においては、植物細胞の原形質に含まれる色素体が重要な役割を果たしている。この色素体には、子葉や貯蔵器官にあって貯蔵デンプンをもつアミロプラスト、トマトの果実やニンジンの根に含まれる橙(とう)色の有色体、ダイコンにあるクロロフィルを欠く白色体、緑の葉にあるクロロフィルをもつ葉緑体などがある。葉緑体の微細構造は種子植物、コケ植物、藻類でそれぞれ特徴があり、高等植物の葉緑体はラメラとグラナの分化が著しい。植物の細胞には、原形質とははっきりと仕切られた液胞がある。液胞はとくに成熟した細胞にあり、若い細胞や老化したものにはみられない。液胞の中の細胞液には無機イオン、多糖類、タンパク質、アミノ酸、さらに有毒植物の場合には毒の本体となるアルカロイドや配糖体が含まれる。植物の成長は、古い細胞に新しく分裂した細胞が積み重なることによるもので、胚(はい)から生じた幼植物でも、すでに細胞分裂が行われる部分は限定されている。細胞分裂を盛んに行うのはおもに茎や根の先端にある成長点、茎の維管束にある維管束形成層、樹皮の内側にあるコルク形成層などである。このような成長点や二次分裂組織が活動を続ける限り、植物は無限に成長し続ける。この点で植物は動物と大きく異なっている。
[金井弘夫]
植物の分類が学問的体系を整えたのは18世紀のリンネの時代からである。リンネは、植物の生殖器官の特徴、とくに雄しべの数によって種子植物の分類を試みたが、これは人為的な分類法であった。19世紀になってC・R・ダーウィンの進化論が発表されると、系統の概念に基づいて、単一の形質だけでなく、生活型、形態、遺伝、生態などの特徴を多面的、総合的にとらえた系統分類が行われるようになった。現在、日本では高等植物の分類体系はドイツのエングラーによるものが広く用いられているが、ほかにイギリスのハッチンソンJohn Hutchinson(1884―1972)、アメリカのベッシーCharles Edwin Bessey(1845―1915)、クロンキストArthur Cronquist(1919―1992)、ソ連のタクタジャンАрмен Леонович Тахталжян/Armen Leonovich Tahtalzhyan(1910―2009)らの分類体系も参考にされている。藻類の分類体系に貢献したのはパシャーAdolf Pascher(1881―1945)である。
高等植物の分類の基準となる形質は生殖器官、すなわち花であるが、下等植物のうち、とくに藻類の分類では、生殖細胞の鞭毛(べんもう)の構造、細胞壁の主要構成成分、同化色素、同化貯蔵物質などが重要な分類形質となる。以下に植物の分類を示す。
①第1門 藍藻(らんそう)植物門Cyanophyta 原核植物で、核物質は核膜に囲まれないで存在する。有性生殖を行わず、分裂で無性的に増殖する。単細胞体、糸状体などの体制をもつ。細胞壁はおもにムコペプチドからなる。同化色素はクロロフィルaのほかにタンパク色素のフィコビリンを含み、これが藻体を藍(あい)色にみせている。同化産物はグリコーゲンである。藍藻粒、油脂などを貯蔵物質とする。水生のものが多いが、地衣類と共生するものもある。現在わかっている種はユレモ、ネンジュモ、スイゼンジノリなど約1400種である。
②第2門 紅色(こうしょく)植物門Rhodophyta 体制は多細胞の糸状、紐(ひも)状、葉状を示す。細胞壁はセルロースのほかにマンナン、キシランもわずかに含む。同化色素はクロロフィルaとdのほかにフィコシアニンなどの紅色色素も含むので赤くみえる。同化産物は紅藻デンプンである。生殖細胞は鞭毛をもたないので、他の鞭毛をもつものより原始的段階にあるとみられる。多くは海産で、テングサ、アサクサノリ、カワモズクなど約4000種が記録されている。
③第3門 黄色(おうしょく)植物門Xanthophyta 不動性の単細胞、群体、糸状体などの形態をもつ。鞭毛は1本、2本、3本とさまざまである。同化色素はクロロフィルa、c、eのほかキサントフィル。同化産物は炭水化物の一種クリソラミナランなどである。細胞壁はセルロースで二重になるものが多い。ヒカリモ、ケイソウ、フウセンモなど約1万種が記録されている。
④第4門 橙色(とうしょく)植物門Pyrrophyta 単細胞で、運動性のものは不等長の鞭毛をもつ。無性的に二分裂法により増殖する。同化色素はクロロフィルaとc、カロチン、キサントフィルを含み、黄色を帯びる。同化産物はデンプン、油脂などである。海水に多く、異常増殖により赤潮現象をおこす。ヤコウチュウ、ムシモ、ツノモなど約1000種が知られている。
⑤第5門 褐色植物門Phaeophyta 多くは肉眼でわかる大きさの藻類で、見かけは根、茎、葉に分化するものがある。細胞壁はセルロースである。同化色素はクロロフィルaとc、フコキサンチン、カロチンである。貯蔵物質はラミナラン、油脂、マンニトールなどである。有性世代と無性世代の交代がはっきりしている生殖法を行う。海産種がほとんどで、とくに寒海域に分布するものに食用とされるものが多い。コンブ、ワカメ、ホンダワラなど約1500種が現生する。
⑥第6門 ミドリムシ植物門Euglenophyta 単細胞体で鞭毛をもつ。同化色素はクロロフィルaとb、カロチン、キサントフィルを含む。体の先端に光を感じる眼点がある。細胞壁をもたず、運動性があることなどは動物的である。二分裂法による無性生殖が知られている。有機物の多いたまり水に繁殖すると、全体が緑色となる「水の華(はな)」とよばれる現象をおこす。ミドリムシ、コラシウムなど約400種が知られている。
⑦第7門 緑藻(りょくそう)植物門Chlorophyta 藻体はさまざまな形をとり、単細胞のものから、仮根のようなもので着生する糸状体、葉状体などがある。多くは淡水産で、ときには陸上の岩石、土壌、樹皮上にも生育する。同化色素はクロロフィルaとb、カロチン、キサントフィルで、デンプンを貯蔵する。生殖法は有性世代と無性世代を交代するものが多い。葉緑体の構造、同化産物、細胞壁の主要構成成分などは陸上植物との類縁を示す。アオミドロ、ボルボックス、カワノリなど約5000種がある。
⑧第8門 車軸藻(しゃじくも)植物門Charophyta 外形はスギナに似て、肉眼で見える大きさの藻類である。葉緑体、同化産物など多くの点で緑藻植物に似るが、栄養体の成長法、生殖器官の構造は異なる。細胞壁はセルロースで、外層はクチクラからなり、さらに外には石灰質が沈着するので固く、化石として残りやすい。同化色素はクロロフィルaとb、カロチン、キサントフィルで、デンプンを貯蔵する。生殖は卵と精子による有性生殖を行う。造卵器は多細胞である。淡水産で、シャジクモ、シラタマモ、フラスモなど約200種ある。
⑨第9門 コケ植物門Bryophyta 蘚類(せんるい)と苔類(たいるい)に分けられる。蘚類は単相の配偶体をもち、茎、葉の分化がみられ、根は仮根で高等植物の根毛状である。ときには通道組織として道束をもつ。複相の胞子体は小形で光合成能力もほとんどなく、栄養は有性世代に寄生的に依存する。一方、苔類では背腹性のある配偶体は葉状を呈し、茎と葉の分化はない。葉緑体は小形でピレノイドをもたない点でも蘚類とは異なる。コケ植物は、緑藻植物とシダ植物などの陸上植物との中間的形質を示す植物である。蘚類にはミズゴケ、スギゴケなど、苔類にはゼニゴケ、ツノゴケなどがあり、その数は2万種を超えている。
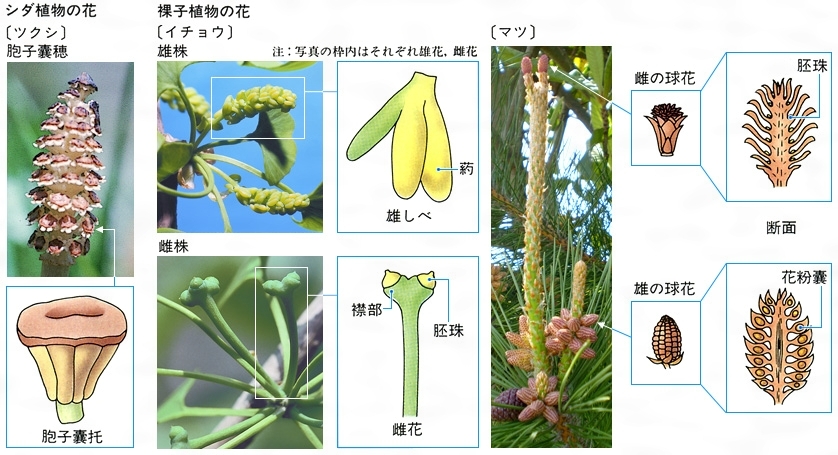
⑩第10門 シダ植物門Pteridophyta 陸上植物のうち、維管束をもつが種子の形成をみない段階の植物をいう。まれに水中に生育するものがある。古生代初期のころから陸上性のシダ植物の化石が発見されており、陸上植物の祖先的存在とみられる。日常見慣れているシダは複相の胞子体で、シダの生活史の大部分はこの胞子体で営まれている。胞子体は茎、葉、根が分化するが、根は不定根で、なかには退化したものもある。茎には木部と篩部(しぶ)をもつ維管束が発達し、普通の木部には仮道管がある。シダ植物の葉には大葉と小葉、輪葉のタイプがある。大葉は種子植物でもみられるもので、葉へ入る維管束が茎の中心柱から分かれるときに葉隙(ようげき)を生じる。ヒカゲノカズラなどにみられる小葉では、葉へ入る維管束は茎に葉隙を残さない。スギナやトクサにみられる輪葉は、節(せつ)に輪生する葉へそれぞれ1本の葉跡が茎の中心柱に向けて葉隙を残さずに入る。シダ植物の配偶体は、胞子が発芽して生ずる前葉体で、単相である。小さくて寿命の短い前葉体には多細胞の造卵器と造精器を生じる。精子には鞭毛がある。大葉をもつシダ植物は正月の飾りにされるウラジロ、食用にされるワラビなどがある。約1万種が知られている。
⑪第11門 種子植物Spermatophyta 種子をつくる維管束植物である。複相の胞子体がよく発達し、茎、葉、根が分化する。茎は維管束形成層による二次成長を行う。葉は形態的に多様で、単葉、複葉、ときには刺(とげ)や巻きひげにも変態する。根の先端には単細胞の根毛が発達し、吸収器官として効率のよい形態をとる。花はそれ自体、まとまった生殖器官を形成している。裸子植物の花は花被(かひ)がなく、鱗片(りんぺん)上に胚珠が裸出している。被子植物には目だった花被があり、多くは花弁と萼(がく)に区別できる。胚珠は子房に包まれる。裸子植物と被子植物は多くの点で異なるため、それぞれ被子植物亜門、裸子植物亜門に区別される。現生種でみると、裸子植物亜門は約800種、被子植物亜門は20万~30万種とされる。被子植物は、現在、陸上でもっとも優勢な植物群といえる。
[金井弘夫]
植物には単細胞体、多細胞体の両様があるが、多細胞体でも、その体制は動物に比べて簡単で、組織の分化も単純である。植物体では栄養器官と生殖器官を区別することができる。ミドリムシのような単細胞体や、ユレモなどの下等な多細胞体の植物では、生殖器官も単細胞で、とくに分化がみられない。このような植物では1個の細胞内によく発達した小器官が認められ、これが養分の同化、分泌、貯蔵などに働いている。これら植物の光合成色素体の微細構造は分類群の特徴となっている。一方、高等植物、とりわけ陸上植物では、それぞれ器官や組織が分化している。
[金井弘夫]
栄養器官は、地上部に伸びる茎、光合成を行う葉、養分を吸収し植物体を支える根からなっている。茎は頂端部で盛んに細胞分裂を行い、そこに葉を側生する。茎の内部構造では、木部と篩部からなる維管束、およびその内側中心部に髄、外側を囲む皮層と表皮が認められる。種子植物の維管束には形成層があり、二次的に肥大成長するものが多い。しかし、シダ植物にはこのような維管束形成層はないし、コケ植物では維管束のかわりに、茎の中心に伸長した細胞群からなる道束が認められる。茎から生じる枝は葉腋(ようえき)にできる芽の伸びたものであるが、ときには茎ではなく、直接根(サツマイモ)や葉(コモチマンネングサ)から生じる不定芽もある。コケ植物やシダ植物では枝は葉腋から分枝しないで、茎の頂端分裂組織で二又分枝するのが普通である。茎は地上部を支え、物質の運搬、貯蔵を行うが、ときには本来の形態と著しく違う形を示すことがある。ジャガイモの根茎、タマネギの鱗茎、ワラビの地下茎、ブドウの巻きひげ、ナギイカダの葉などは、いずれも茎の変態したものである。
[金井弘夫]
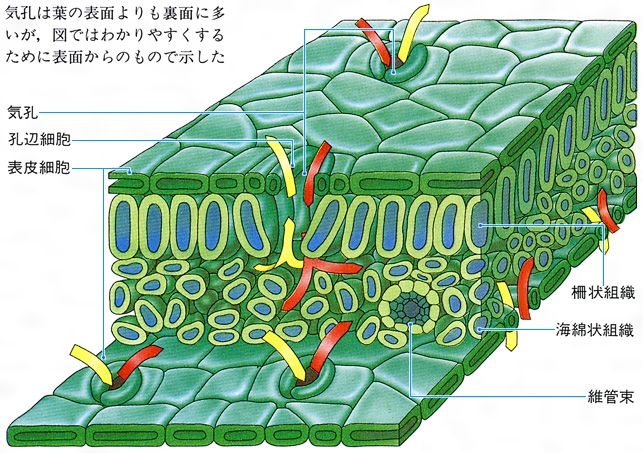
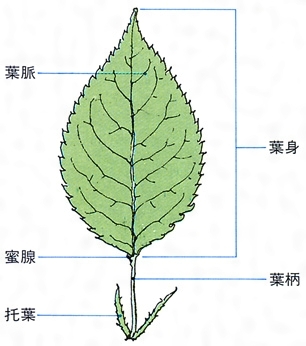
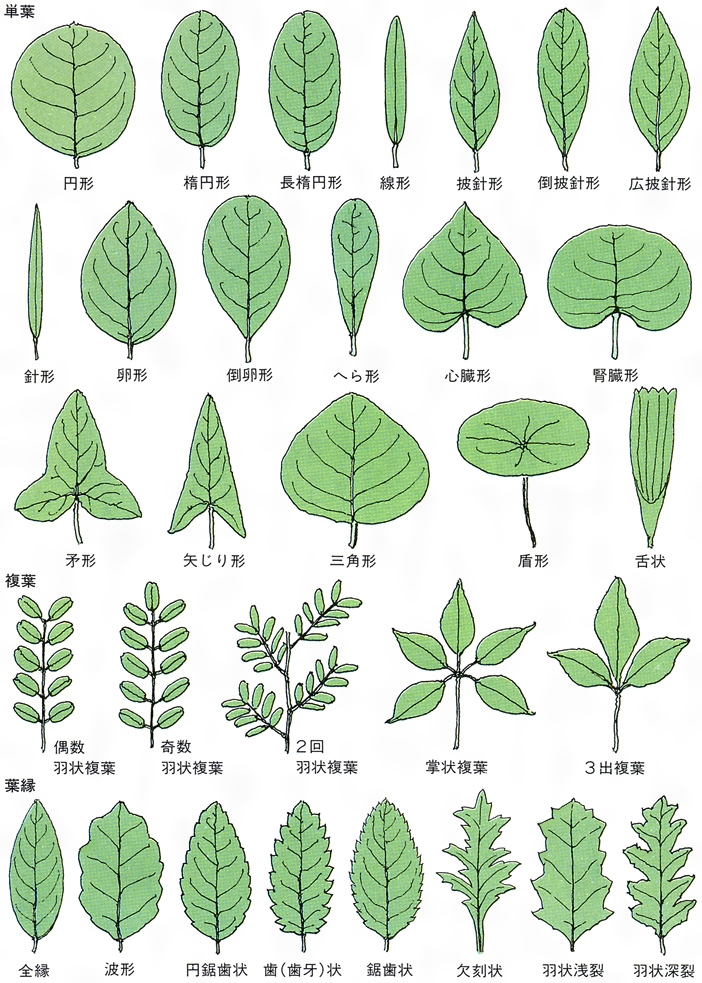
葉は茎頂近くから発生する。葉の原基が生じると周縁分裂組織の活動や介在成長によって平面的な葉がつくられる。葉の表面には気孔が分布し、ここでは二酸化炭素や水蒸気などのガス交換が行われる。表皮の内側をつくる葉肉は、葉緑体を含む柵(さく)状組織と細胞間隙の多い海綿状組織に分化しており、光合成の場となっている。維管束は茎から連続して葉に入り、水分や養分の運搬を行う。葉の外形は葉身、葉柄、托葉(たくよう)などの部分に区別されるが、葉柄や托葉がなく、かわりに葉鞘(ようしょう)となることもある。葉身は縁に鋸歯(きょし)をもつもの、小葉に分かれて複葉となるもの、ネギのように円筒状をした単面葉のもの、ハスのような楯(じゅん)状葉など、さまざまな外形がある。子葉は胚につくられる最初の葉で、普通葉と異なる形態を示し、冬芽を包む鱗片葉、花芽を覆う包葉などとともに異形葉とよばれる。花を構成する各部分は雄しべ、雌しべを含めてすべて葉の変態である。サボテンの針葉、ウツボカズラの嚢(のう)状葉、サンショウモの根状葉なども葉の変態したものである。
[金井弘夫]
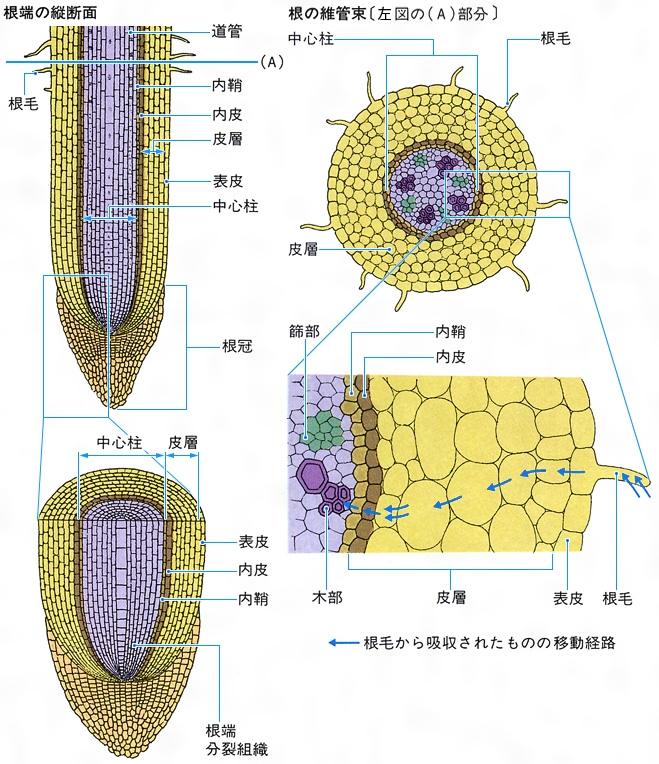
根は茎と同様に根端とよばれる頂端分裂組織によって伸長し、先端部は根冠とよばれる細胞群で保護されている。根の表皮には単細胞の根毛が分化し、水や無機塩類の吸収を行う。根には中心柱を囲む1層の細胞層(内皮)があり、吸収した物質の取捨選択を行うとされる。内皮と維管束の間の柔細胞の部分を内鞘といい、側根や根の形成層をつくる分裂組織である。維管束配列は茎とは異なり、木部と篩部が交互に並ぶ放射中心柱である。主根以外の茎や葉に生ずる根を不定根という。根から芽を生じることはあるが、直接葉を生じることはない。根は地中で植物体を支え、水や養分を吸収する役割をもつが、それ以外の役割をもつものも多い。サツマイモの貯蔵根、タコノキの気根、ヒルギの呼吸根、キヅタの付着根などがその例といえる。
[金井弘夫]
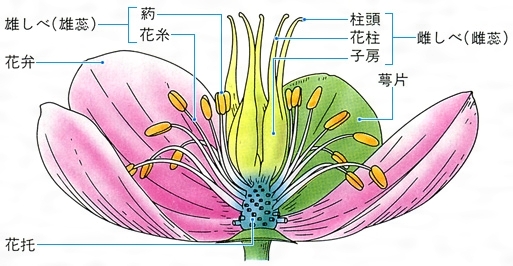
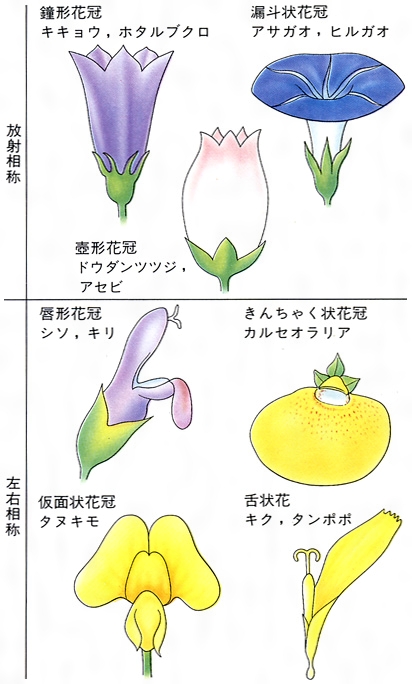
生殖器官をみると、種子植物では複雑に分化した形態を示し、花をつくる。シダ植物におけるヒカゲノカズラなどの胞子嚢は花と相同するという意見もあるが、一般にはこれを花とはみていない。種子植物の花は、胚珠(シダ植物の前葉体に相当する部分)を生じる有性生殖器官であり、胚珠を保護する心皮、花粉を生じる雄しべ、これらを覆い囲んでいる花弁や萼などによってその祖先をたどると、葉の変態としてとらえることができる。被子植物の花には単生花、花序をつくる花、左右相称花、放射相称花、有花被花、無花被花などさまざまな外部形態をもつものがあり、これは種を区別する重要な特徴である。また、両性花、雄しべか雌しべのいずれかを欠く単性花、子房の位置、雄しべの数と配列なども、分類するうえでの特徴となっている。このほか、果実、種子の形、分裂細胞にみられる染色体の数や形なども、高等植物の分類においては役だつ資料となる。
[金井弘夫]
植物は原則として動物のように移動できないため、生活は環境に依存することになる。この環境に適応するということから、植物には特有の生理現象がみられる。光合成、水や無機塩の吸収と移動、同化産物の貯蔵、窒素や炭水化物の代謝などがそれであり、植物はこれらの作用を行いつつ発芽、成長、分化、老化、死という生活環を完結している。植物の生活環の始まりは受精卵から成長した胚であり、種子の中で子葉、幼根、胚乳などを分化する。種子の段階では、成長は一時的に停止して休眠に入り、呼吸などの生理作用も不活発になる。この休眠は植物の環境に対する適応の一つで、樹木の芽、ジャガイモの塊茎、チューリップの鱗茎などでもみられる。休眠中の種子の発芽は吸水に始まり、まず幼根を下方へ伸ばす。このとき、水分条件のほかに温度、酸素、光などの諸条件の影響も大きい。発芽が始まると、貯蔵器官である胚乳、子葉に含まれる物質が分解され、成長のエネルギー源とされる。成長と分化はほとんど同時に行われる。分裂能力をもつ細胞はすべて分化する能力をもっており、芽と根の分化はホルモンのオーキシンとアデニン誘導体のバランスによって決められる。普通、オーキシンが多いと根が分化し、サイトカイニンが多いと芽が分化する。また、花芽の分化は日長の影響を受ける。植物には、日長が長くなると花芽を分化する長日植物(コムギ、ホウレンソウ、ダイコン)、日長が短くなると花芽を分化する短日植物(イネ、キク、アサガオ)のほか、日長に関係がない中性植物(タンポポ、トマト、キュウリ)などがある。植物体の各部分は成長とともにしだいに機能が低下し、やがて成長速度がゼロになる。これを老化といい、老化の時期は葉、茎、根などで異なっている。それぞれの時期が異なるのは、各器官が互いに影響しあうためである。たとえば、葉の老化は茎頂分裂組織の活性だけでなく、根の存在も関係している。つまり、根で生産されたサイトカイニンは葉へ運ばれ、葉の老化を防ぐ働きをするわけである。このように植物の成長、分化、老化はホルモンのバランスによって調節されている。
植物の成長、分化に必要な養分は無機物から合成される。この無機物から有機物を合成する仕組みを光合成という。光合成は植物だけにみられる生理作用で、細胞内の葉緑体は光エネルギーによって水を分解し、生じた水素と大気中の二酸化炭素を酵素反応で結合させて炭水化物を合成する。この炭水化物と窒素、リンなどの無機塩類は細胞内で結合されて、アミノ酸、タンパク質など、原形質に含まれる有機物に変えられる。このため、植物は光合成で生産された炭水化物以外にも窒素、リン、カリウム、マグネシウム、カルシウム、鉄などの元素を必要とし、これらの一部が不足しても成長は不完全となる。これらの無機塩類は、水に溶けた状態で根からイオンの形で吸収される。根における吸水の機構は、隣接する細胞との濃度差がおこす拡散流と呼吸エネルギーを使って行われる。根から吸収された水は、葉の気孔や木の皮目(ひもく)から水蒸気として蒸散する。水が蒸散すると、葉の葉肉組織の細胞の拡散圧が高まり、水はさらに道管を通じて移動していく。水に溶けた無機イオンもこの蒸散流によって運ばれる。植物が合成したり、根や茎に貯蔵した同化物質も水などとともに移動する。同化産物のうち、炭水化物はブドウ糖、ショ糖としておもに篩管(しかん)を通る。また、上方の葉で生成された物質は茎の若い成長部へ、果実をつけた枝の物質はすべてこの果実へ移動する。根へは茎の下部の葉で生産された物質が供給される。さらに植物が生成した有機窒素化合物は、グルタミン、アスパラギンなどとなって道管を蒸散流にのって移動する。この同化物質の移動においては、葉で生産されたものがそのままの状態で運ばれる場合と、ふたたび分解されて運ばれる場合とがある。
植物は光合成によって生産された養分を栄養源とし、これを分解して成長、分化のエネルギーとする。この生理作用をつかさどるのが呼吸であり、高等植物の行う効率的エネルギー生成は酸素呼吸である。植物の酸素呼吸は動物と共通の生理作用で、ブドウ糖を酸化して水と二酸化炭素に分解してATP(アデノシン三リン酸)を生産する。このATPは植物の成長、分化にエネルギーとして利用される。植物の成長、分化の生理機構のなかでは、さまざまな植物ホルモンが重要な役割を果たしている。たとえば、果実の成熟を促す植物ホルモンとしてエチレンなどがあるが、これらは植物形態の変化に生理的な影響を与えているといえる。
[金井弘夫]
植物の生育場所は種属によって決まっている。これは、植物の生育が、光、温度、水などの環境条件を必要とするためである。また、地形、地史、気候の変動、種子散布能力、栄養要求の差などによっても自生する植物の分布域が決まってくる。狭い地域にのみ分布する植物を固有種という。日本で約4000種の自生する植物のうち、約300種が固有種であり、スギは日本の代表的固有種の一つである。一方、広く世界中に分布している植物を広布種という。種子植物のうち、世界の陸地面積の半分にわたって広く分布する広布種は約30種である。もっとも広く分布する植物にアシがあり、南アメリカのアマゾン流域を除く世界各地に分布する。植物の分布の見方には二つの方法がある。一つは植物の種属の構成に基づく「フロラ分布」で、他の一つは植物の生育環境を重視した「生態分布」である。フロラ分布は植物分類学的、植物誌的に植物の分布を研究し、生態分布は植物生態的、植物地理的に分布を理解しようとするものである。
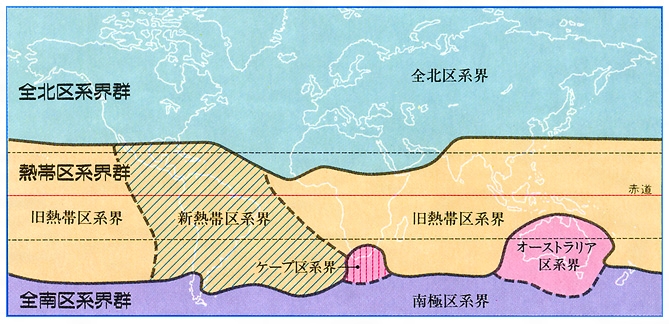
世界各地の植物の分布を種属ごとに比較すると、特色ある植物の分布域が認められる。これを植物区系といい、世界各地を大きく6区域に区分することができる。このような植物区系は生態的な気候条件よりもその地史に影響されるところが大きい。たとえば、同じ熱帯降雨林であっても、ニューギニア島と南アメリカのアマゾン流域ではフロラの構成要素が著しく異なるので、別の植物区系として扱われている。以下、植物区系について触れる。
①全北区系(全北区系界) 熱帯を除く北半球の大部分を含む。代表種にはマツ、ヤナギ、クリ、サクラ、カエデ、ユリがある。全北区系は気候や構成種によってさらに細分化され、日本は中国南部からヒマラヤまでつながる日華区系区に属する。
②旧熱帯区系(旧熱帯区系界) 旧大陸の熱帯、ハワイ付近の南太平洋地域を含む。代表種はコショウ、フタバガキ、タコノキ、バナナ、ヤシである。
③新熱帯区系(新熱帯区系界) メキシコ以南の新大陸全域。ただしパタゴニア地域は除く。代表種はサボテン、オオオニバス、パイナップル、カンナ、リュウゼツランなどである。
④オーストラリア区系(オーストラリア区系界) オーストラリアとタスマニア島を含む地域である。代表種はアカシア、ユーカリ、バンクシアなどである。
⑤ケープ区系(ケープ区系界) アフリカの南端のごく狭い範囲だが、アフリカ大陸の他の地域とは異なる特徴的フロラをもつ。代表種はエリカ、アロエ、マツバギクなどである。
⑥南極区系(全南区系界) 南アメリカ大陸の南端、南温帯太平洋や南インド洋の島、ニュージーランド、南極大陸を含む地域である。代表種はナンキョクブナである。
なお、生態学的にはこの六つの植物区系を、さらに三つの群にまとめることもある。すなわち、①をそのまま「全北区系界群」、②と③をまとめて「熱帯区系界群」、④~⑥をまとめて「全南区系界群」とする方法である。
日本の植物区系の区分については、まだ定説がない。その理由としては、日本の植物相が変化に富み、地史的にも分布のうえからも、さまざまな要素が重なって複雑な様相を示すことがあげられる。また生態的、気候的な分布のうえからみても、日本は山国であり、緯度の違いによる水平的植物分布と海抜高度の違いによる垂直的植物分布が入り組んでいることも分類を困難にしている理由の一つである。このため、ここでは、気候的植生と一致する分布域を示す種類を基準とした生態的区分を示すにとどめる。
①ウバメガシ群団 暖温帯に属する気候に分布し、九州、四国、紀伊半島の一部から亜熱帯の要素も加わる沖縄まで含まれる。代表種としてはスダジイ、タブノキ、シイノキがある。
②クリ群団 九州から関東地方にかけての低地、低山帯を含む地域に分布し、代表種としてはクリ、ツガ、マツ、コナラ、クヌギ、シデ類などの暖帯落葉樹が多く、温暖地の常緑樹林と冷温地のブナ林の中間にみられるものである。
③ブナ群団 日本中部の山岳地帯、東北地方に分布域をもち、日本の植物相のなかでも、もっとも特徴的なものである。日本に分布する固有種の大部分はこの地域に含まれる。代表種はブナ、ミズナラ、カエデ、トネリコ、ハルニレ、シナノキなどであり、固有種ではヤマグルマ、カツラ、フサザクラ、コブシ、シラネアオイなどがある。
④トドマツ群団 日本中部以北の亜高山帯、北海道に分布域をもち、亜高山針葉樹林帯をつくる。気候的には寒帯下部に属する。代表種にはドロノキ、ケショウヤナギ、ダケカンバ、ツルツゲ、オガラバナ、ツバメオモト、タカネナナカマド、ジンヨウイチヤクソウなどがある。
⑤ハイマツ群団 日本中部の高山帯と北海道の高山帯にみられ、寒帯に属する。ハイマツ、ガンコウラン、コケモモなどの低木のほかに、高山のお花畑を埋めるハクサンイチゲ、シナノキンバイ、クロユリ、コバイケイソウ、コマクサ、チョウノスケソウなどが生育している。
いずれにしても、このような植物の分布については、種の分化の問題を含んでいるため、今後の種属誌的研究の成果に期待する面が多い。
[金井弘夫]
人類の植物利用の歴史は古い。人間は有史以前からその生活を維持していくために植物を利用し続けて今日に至っている。現在でも有用植物の利用は広く各方面に及んでおり、とくに食用としての依存度は大きい。化学合成技術の発達は、人間生活における植物への依存度を軽減したが、それでもなお、植物の重要性は失われていない。人間の歴史の初期のころは、食糧を採集と狩猟によって得ていたが、やがて定まった土地に住居をつくり、火を用いる生活が行われるようになると、人間は採集した木の実や種子の食べ残しなどから、芽が伸び、実がなることを知った。やがて住居近くに種子を播(ま)いて、自然に芽が出るのを期待するようになる。これが栽培の始まりである。初期の栽培植物の特徴は、一つのものが多目的に利用されたということである。
たとえば、クワ科の一年草であるアサは、中央アジア原産で、ペルシアでは紀元前2000年には栽培されており、繊維植物、薬用植物、香辛植物とされ、良質の油もこれから得ていた。また、中国では、家の周りに野生するクワの果実は食用に、葉はカイコの餌(えさ)に、材は細工用に、また根からは黄色染料をというように多目的に利用してきた。熱帯地方の海岸に広く分布するココヤシも多目的に利用されるものの一つである。種子からとれるココナッツミルクは養分に富んだ飲み物とされるほか、胚乳の「コプラ」とよばれる部分から油をとり、果肉の繊維部分はロープ、マットなどに使われる。また、ココヤシの葉で屋根を葺(ふ)き、開花前の花序の軸を切って得られる樹液からやし糖、やし酒もつくられる。このように多目的に利用される植物も、初めは食用としての利用がなされ、その後しだいに多くの利用法がくふうされたわけである。
コムギはイネとともにユーラシア大陸の栽培植物としてはもっとも古いものであり、イラク東部からは前7000年ころの炭化したコムギがみつかっている。現在、イネやコムギは南極大陸以外のあらゆる所で栽培されている。また、トウモロコシとジャガイモはアメリカ大陸原産の代表的な栽培植物である。前5000年のメキシコ遺跡からは、もっとも古いトウモロコシの野生株がみつかっている。両植物は15世紀以降、世界各地に広まり、現在は食用、飼料として広く栽培されている。このほか、マメ類は一般にタンパク質に富み、種子の水分含量が少なく貯蔵性があることから、重要な食用作物となっている。栽培されるマメ類は、おもに温暖帯性の一年生のものである。
植物は薬用としても利用されるが、その歴史は古い。薬用植物にはアルカロイド、配糖体などの有毒成分を含むものが多いが、人間はその毒性を病気の治療に有効に活用してきた。古代エジプトにおいては、すでにケシから得られるアヘンの麻酔性が知られており、苦痛を和らげる薬として、これを利用していた。また、アイヌが「熊(くま)狩り」に矢毒として用いたトリカブトは猛毒をもつが、古代中国では、少量を用いれば神経痛の薬、強心剤、血圧降下剤として効果があることを知っていた。現在多くの医学的治療に用いられているステロイド系物質のコーチゾンは、リウマチ性関節炎の治療や、合成女性ホルモンとして避妊用ピルに用いられるが、最近ではヤマノイモ属やリュウゼツラン科の植物から大量に得ることが可能となった。
さらに植物は、工業用に盛んに利用されている。木材は建材、装飾具、器具などに多用されている。木材を酸欠状態でゆっくりと炭化させたのが木炭である。材を砕いて加熱し、これによって生じる蒸気を集めるとメタン、水素ガス、メチルアルコール(木精)、酢酸などが得られるが、これは木材を構成する多糖類のセルロースが分解するためである。松柏(しょうはく)類の材には松脂(まつやに)を含むが、これを蒸留するとテレビン油が得られる。野球の投手が滑り止めに用いるロジンはこの蒸留で残った成分である。これら木材の細片は木材パルプとして紙をつくるが、さらにこれを化学処理するとレーヨンやプラスチックとなる。木材パルプのセルロースから生産されるプラスチックをセルロイドという。セルロイドは燃えやすい欠点があるため、最近ではさらに化学処理を加えて不燃性のセルロースアセテートとし、写真フィルムなどに用いられている。人造皮革も植物からつくられる。これは、木材セルロースの硝酸塩とトウゴマの種子からとれるひまし油を結合させたものである。このほか、植物は、染料、飲料用、嗜好(しこう)用など、さまざまな分野に利用され、人間の生活に物質的豊かさを与えてくれている。最近では、森林の樹木から発生する気体成分が健康保持に役だつとして注目されている。
[金井弘夫]
『小野知夫著『基礎植物学』(1954・裳華房)』▽『井上浩著『植物学入門講座』全6巻(1980・加島書店)』▽『相見霊三・朝比奈正二郎他編『現代生物学大系』全14巻(1985・中山書店)』▽『佐竹義輔著『植物の分類・基礎と方法』(1964・第一法規)』▽『山岸高旺編『植物系統分類の基礎』(1974・北隆館)』▽『井上浩編『植物系統進化学』(1974・築地書館)』▽『河野昭一著『植物の進化生物学Ⅱ 種の分化と適応』(1974・三省堂)』▽『熊沢正夫著『植物器官学』(1979・裳華房)』▽『館岡亜緒著『植物の種分化と分類』(1983・養賢堂)』▽『原襄著『植物の形態』増訂版(1984・裳華房)』▽『沼田真編『生態の事典』(1976・東京堂出版)』▽『沼田真編『植物生態の観察と研究』(1978・東海大学出版会)』▽『桜井英博他著『植物生理学入門』(1980・培風館)』▽『清水碩著『植物生理学』(1980・裳華房)』▽『朝日新聞社編・刊『朝日百科 世界の植物』(1978)』
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponserd by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponserd by ![]()
 (へんぱん)たり、群獸
(へんぱん)たり、群獸
 (ひし)たり。
(ひし)たり。字通「植」の項目を見る。
出典 平凡社「普及版 字通」普及版 字通について 情報
Sponserd by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponserd by ![]()
…耐乾性drought resistanceが大きく,乾燥して土壌水分が乏しい土地に生育できる植物。低温・塩類の過多などで土壌からの吸水が困難な土地に生育する植物を含める場合がある。…
…いずれにせよ,エネルギー転換にかかわるエネルギーのシステムと,それの方向を指示し,自己保存,自己増殖を行う情報のシステムとが組み合わさったとき,初めて,生物が生じたことは確かである。
[生物の分類]
生物は動物と植物に大別されるが,これは外界からの物質やエネルギーのとりこみ方の違いに基づくものである。すなわち,主要なエネルギー源として太陽エネルギーを,体を構成する物質源としては無機物質をとりこむ方式で生きるものが植物であり,エネルギー源,物質源を有機物質に頼るのが動物であると考えてよい。…
…現在,地球上には約30万種の植物が知られている。それらのうち人類の生活になんらかの関わりがあり,食料,香辛料,衣料,糸や籠の材料,住居の建築や木工細工,燃料,薬,さらには美的な観賞対象としての園芸植物にいたるまで,さまざまな形で利用されている植物は数万種にのぼるであろう。…
※「植物」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponserd by ![]()
福岡県福岡市博多区の櫛田神社の夏祭り。壮麗な山笠で知られる。今日,山笠には飾り山笠と舁き山笠(かきやまがさ)の 2種類がある。明治時代に電線が架設されて以降,物語場面の人形などを飾りつけた高さ 15m...