デジタル大辞泉 「魚類」の意味・読み・例文・類語
ぎょ‐るい【魚類】
[類語]脊椎動物・無顎類・両生類・爬虫類・鳥類・哺乳類
Sponsored by ![]()
翻訳|fish
Sponsored by ![]()
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報 | 凡例
Sponsored by ![]()
体に背骨を備え、水中の運動のためにひれをもち、えらで水中の酸素を呼吸する水生動物群。分類学上は、脊索(せきさく)動物門、脊椎(せきつい)動物亜門、顎口(がっこう)上綱Gnathostomataに属する。一般には、顎骨(がくこつ)がある顎口上綱の魚類と、顎骨のない無顎上綱Agnathaのヤツメウナギ類やヌタウナギ類など(旧、円口類)も含め、「うお(魚)」または「さかな(魚)」と俗称され、便宜的にいっしょにして魚類として取り扱う。体形は紡錘形を中心に多様であり、大きさも最小7.9ミリメートルのスマトラ島産のコイ科の一種から最大20メートル近くにもなるジンベエザメまである。形態の顕著な特徴は、原則として体表が鱗(うろこ)で覆われて保護され、感覚器官系の側線が発達し、多くは水中で体の比重を効果的に調節するためのうきぶくろがある点である。
[落合 明・尼岡邦夫 2015年3月19日]
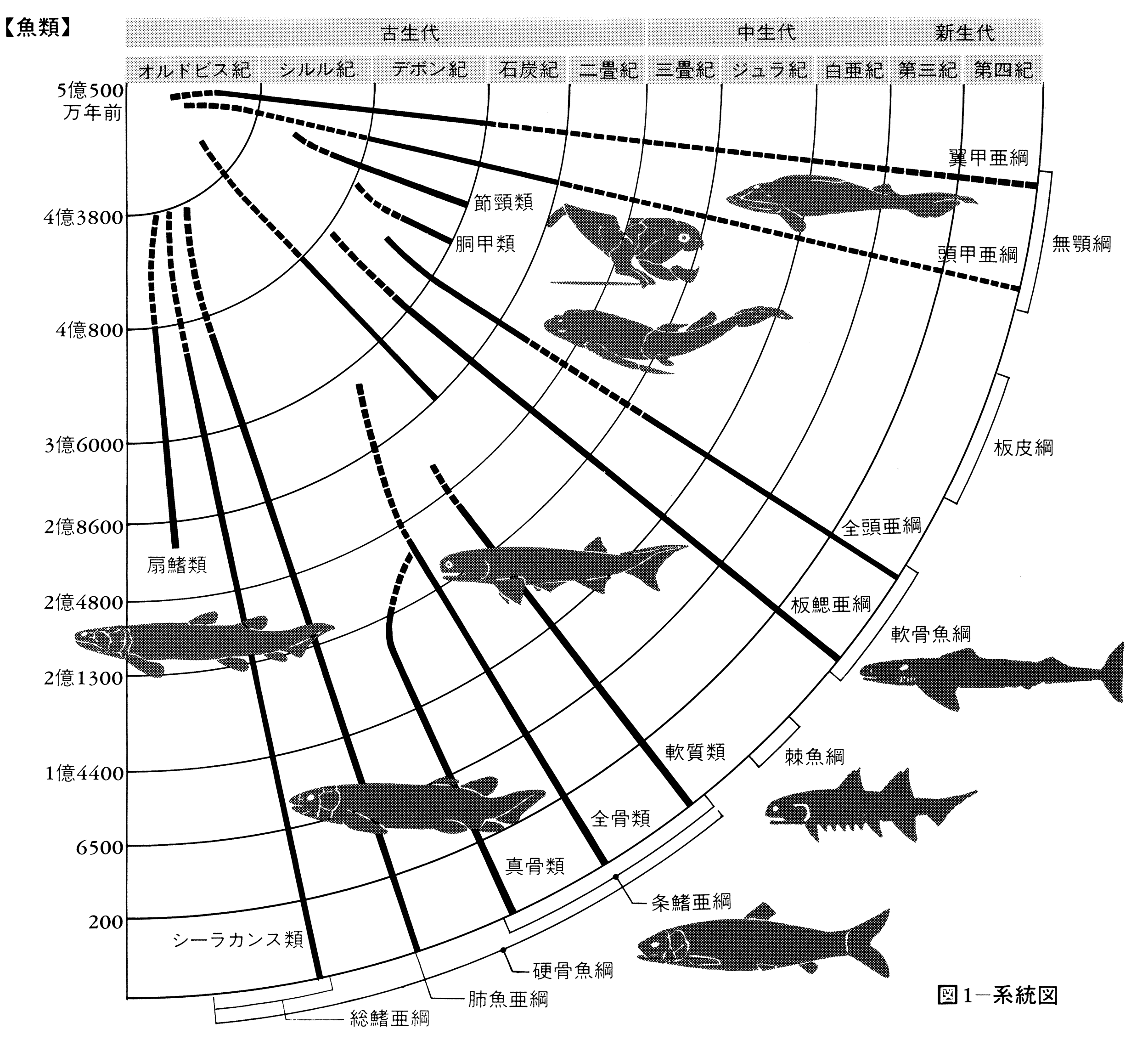
この類の起源は非常に古くて、少なくとも数億年以前に出現していた。古生代末期から現代に至る間に、魚類より遅れて出現した両生類や爬虫(はちゅう)類が、一度大いに栄えたのち、現在は勢力を失ってしまったのに比べ、魚類は、絶えず新しい環境に適応して進化し続けて現代に至っている。
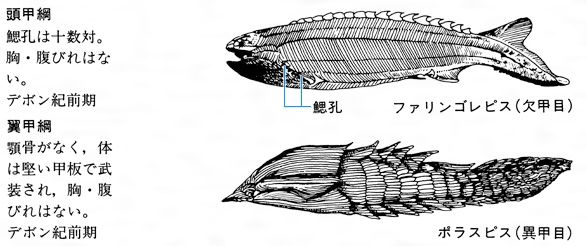
口にあごの骨を欠く無顎類では、体が縦扁(じゅうへん)し、歯のような鱗をもった歯鱗類(しりんるい)がもっとも古いという説もある。しかし、一般的には古生代オルドビス紀に出現した箱形の厚い甲板(こうばん)で包まれた翼甲類のアストラスピス、アランダスピス、プテラスピスなどが初期の魚類と考えられている。翼甲類は、古生代オルドビス紀からシルル紀にかけて徐々に勢力を強めて、次のデボン紀になると、いろいろな無顎類が出現した。古生代の無顎類は鰓孔(さいこう)が15対もあり、頭部がよく骨化した甲板で包まれた頭甲類や、一段と魚らしい形をした欠甲類がその代表である。しかし、シルル紀にあごのある魚類が出現し、デボン紀に急速に勢力を広めたために、無顎類は競争に敗れて、この紀末期にはほとんどが絶滅した。そのなかで、ヤツメウナギ類やヌタウナギ類は寄生あるいは腐肉食性の特殊な生活様式を獲得して、あごをもつ魚との生存競争に生き残った。古代魚の基本的な特徴を現代まで維持しながら世界中の海や淡水に生存し続けている。この類は甲板を欠き粘液性の皮膚で被われて外見上はウナギ型をしているが、えらが袋状で多く、5~15対もある。内部の骨格もすべて軟骨であり、胸びれや腹びれがない。
[落合 明・尼岡邦夫 2015年3月19日]
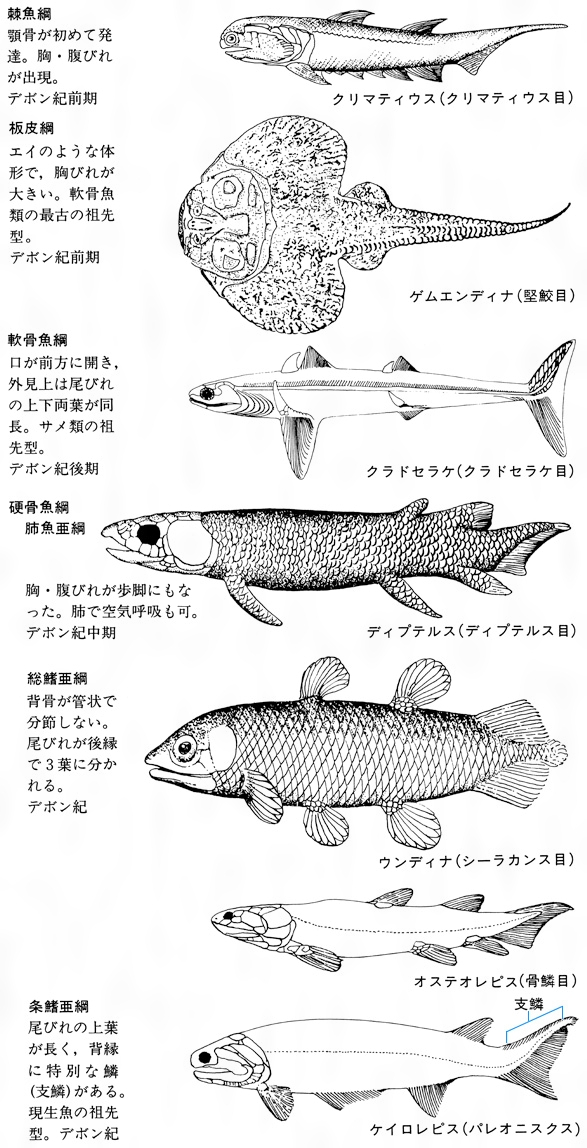
古生代シルル紀からデボン紀は、魚類の進化上特筆すべき時代であり、真のあごをもった原始的な顎口類が続々と出現して栄えた。えらを支えていた最前の鰓弓(さいきゅう)をあごの骨に変化させて、あごをもつようになり、このことによって摂餌(せつじ)機構が飛躍的に改良された。呼吸水と餌(えさ)をいっしょに吸い込んでいた方式から、呼吸水と食物の取り込みの機能を分離することができるようになった。その代表が最古の脊椎動物と考えられている棘魚類(きょくぎょるい)である。棘魚類では、尾びれを除くすべてのひれに棘(とげ)を備え、あごやえらを支えるための軟骨が出現し、胸びれや腹びれが形成された。菱(ひし)形の硬い鱗や頭蓋骨(とうがいこつ)などが部分的に硬骨化している点では、硬骨魚類の原始型に似ている。一方、上顎の骨、歯、えら、ひれなどがサメ類に似ているところから、軟骨魚類に近いとする説もある。また、板皮類は、体の前半部または大部分がじょうぶな骨板(こつばん)(外殻を覆う平板な骨)で包まれており、外形や内部の骨格がサメ類に似ているので、軟骨魚類へ進化したとする説と硬骨魚類へ進化したとする説の二つの仮説がある。約4億4000万年前の古生代シルル紀に初めて出現し、デボン紀に繁栄したこれらの原始的な顎口類は、約2億6000万年前の古生代ペルム紀(二畳紀)の終わりごろから3億年前の石炭紀に適応力を失って絶滅した。
[落合 明・尼岡邦夫 2015年3月19日]
約4億年前の古生代デボン紀前期に出現しており、いろいろな型の古代ザメ(クラドドント型)が淡水域で栄えた。その代表的なものがクラドセラケで、アメリカのエリー湖の地層から発見された。全長1.5メートルほどのほっそりとした体つきで、口が前に開き、胸びれや腹びれが体壁から変わった皮弁状で、棘のある背びれを2基もっていた。尾部の後端部は上方に曲がり、そこに大きな尾びれをもっていた。古代ザメは中生代にはしだいに滅亡し、それにかわって現在のサメに連なる中間型(ハイボドント型)が出現した。この類は種類数では古代ザメに及ばなかったが、あごが多少とも動かしやすいように変わり、胸びれや腹びれも遊泳に役だつよう進化し、摂餌とその行動は現代のサメ・エイ類のように多様性を備え、雄は交尾器をもっていたらしい。しかし、約1億年前の白亜紀中ごろには勢力を失ってしまった。一方、現生のサメ類は中生代のジュラ紀に大半のグループが出現した。現在のサメは各部の骨格がなお軟骨であるが、口が腹側へ移り、あごがいっそう動かしやすくなり、各ひれを支える骨も整った。現在の軟骨魚類はサメ類とエイ類が主流で、板鰓(ばんさい)類(亜綱)に属し、種類数で930種余りおり、そのうち20種余りは淡水にも生息している。なお、現存するサメ類でも、ラブカは古代ザメの形質を残し、またネコザメ類は中生代のサメの形質を残している。
軟骨魚類のもう一つの系列は、ギンザメ類に代表される全頭類(ぜんとうるい)(亜綱)である。この類は、現在、種類数が少なくて劣勢であるが、古生代デボン紀に起源を有し、石炭紀に進化の絶頂に達していた。鰓孔が1個で、第1背びれに起伏できる棘があるなど、硬骨魚類に似た形質を備えている。
[落合 明・尼岡邦夫 2015年3月19日]
古生代シルル紀中期に初めてあごを備えたクリマチスのような棘魚類が出現した。あごを獲得することで、積極的に獲物をとらえることができた。そのために運動能力が向上し、筋肉が発達し、それに伴って脳や神経系が発達した。硬骨魚類の仲間はそのようなものから分かれ、初期には古代魚の特徴を保持しながら、内部骨格が硬骨化しだした。その一部のものは地上生活者としての進化の道をたどった。総鰭類(そうきるい)(シーラカンス類)や肺魚類がそれである。この仲間はデボン紀から石炭紀またはペルム紀にかけて、地球上に勢力を拡大した。しかし、その後はほとんど絶滅してしまい、わずかにその一部が、外敵の少ない南半球で「生きた化石」として生き続けている。シーラカンスや肺魚は、背びれが2基で体の後方にあること、胸びれや腹びれが腹面にあって葉状に伸び、これで四足動物のように体を支えたり、歩行したりした。また、鼻孔が口腔(こうこう)や咽頭(いんとう)に開いて、うきぶくろが肺の働きをし、空気呼吸も可能であった。総鰭類のなかの骨鱗類(オステオレピス類)は、両生類の前駆となる形質を獲得したイクチオステガIchthyostegaのような初期の両生類へ進化するが、この類はペルム紀(二畳紀)に絶滅した。
硬骨魚類の進化の主流は、水中生活者、とくに海洋での生活に適した方向へと進み、魚類としてもっとも優れた体制や機能をもつようになった。このグループを条鰭類(じょうきるい)という。条鰭類は中生代、新生代を通じて骨格が硬骨化し、ひれが遊泳に適するように構造や位置を変え、うきぶくろが比重を調節するようになった。このグループは腕鰭(わんき)類、軟質類、新鰭類の3群に分かれている。腕鰭類(岐鰭(きき)類ともいう)はシルル紀後期に出現し、デボン紀から特殊な方向に進化したが、白亜紀にほとんどが絶滅した。現在アフリカの淡水域にポリプテルス類は生き残っているが、多くの特殊化した特徴を備えている。軟質類は古生代シルル紀後期に出現し、中生代前半にかけて栄えたが、中生代の終わりの白亜紀ごろにはほとんどのものは滅んでしまった。この類で現在まで生き残っているものはチョウザメ類だけである。もっとも原始的な軟質類は、背骨が分節しないこと、尾びれの上部が下部より長いこと、肺があることなどの特徴を備えている。
軟質類のパレオニスクスとよばれるグループから、新鰭類のなかでもっとも原始的な全骨類とよばれていたグループの新しい条鰭類が古生代ペルム紀に出現した。この類では、硬骨化が進むとともに鱗が薄くなりだして2層となり、肺がうきぶくろに変わった。全骨類は中生代の中ごろに栄えたが、中生代の終わりごろに地球上から姿を消してしまった。現在、アメリカ大陸にのみ生存しているアミアやガーパイクは、全骨類の生き残りである。
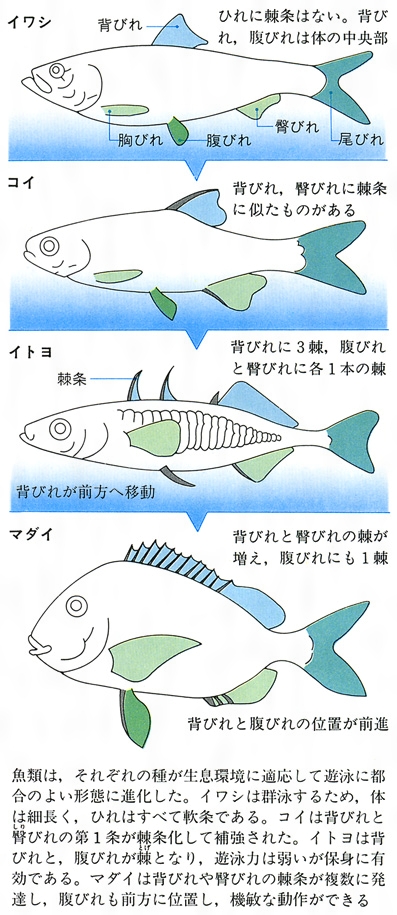
約2億2000万年前の中生代三畳紀後期に、全骨類から一段と進化した真骨類が出現した。この類は硬骨魚類として進化の頂点にあり、世界中の水域に広く分布する。種類数は2万6000種余り、魚類全体の約96%を占め、40目、448科、4278属のなかに置かれ、全脊椎動物のなかでもっとも多くの種が多様に分化している。真骨類は化石種のフォリドフォラスやレプトレピスとよばれるグループから進化したと考えられ、中生代末期(白亜紀)から新生代の中ごろ(中新世)までに、現存する種類の大部分が出現した。骨格がよく硬骨化したこと、浮力調節用の機能としてうきぶくろを備えるようになったこと、上下両顎の骨が可動性になったこと、鱗がいっそう薄くなり、円鱗または櫛鱗(しつりん)で覆瓦(ふくが)状に並ぶこと、尾びれは正尾またはその変形であること、脊椎骨、椎体はよく発達すること、頭の骨が簡略化すること、腸に螺旋(らせん)弁がなくなること、心臓球が退化し動脈球が発達していること、などが真骨類の特徴である。このような特徴の獲得によって、積極的に餌を求めてあらゆる環境に進出することが可能となり、種の多様性を生むことになった。この類では、イワシ類、サケ類などが原始的なグループであり、これらは体の構造がなお全骨類に似ている。しかし、進化に伴って、背びれや臀(しり)びれに棘が発達すること、腹びれの軟条数が減少し、前に1本の棘をもつこと、腹びれの位置は前進し、腰帯は肩帯に付着すること、胸びれが高位に移動すること、上顎は前上顎骨だけで縁どられ、主上顎骨は口を伸出させる働きだけに変化すること、うきぶくろは有気管から無気管に変わる傾向にあること、などの変化がみられる。それぞれの類はひれやうきぶくろ、骨格などに特有な構造を進化させている。スズキ類やカサゴ類では進化の頂点にある。
[落合 明・尼岡邦夫 2015年3月19日]
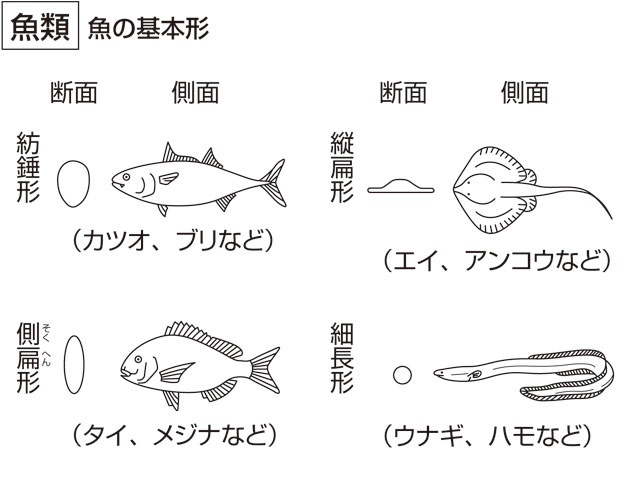
体形は多様であるが、多くは紡錘形で、遊泳中の水の抵抗が少ないように、中央部付近が太く、頭端や尾端へ向かってしだいに細くなる。しかし、タイ類、ハタ類、カレイ類などでは、左右に著しく扁平(へんぺい)になり、これを側扁形という。この体形は岩の割れ目や海底に体を接し、外敵からの食害を防ぐうえで効果的な体形である。一方、エイ類やコチ類のように背腹の方向に扁平になった体形を縦扁形といい、水底に定着して生活するのに適している。ウナギやハモなど、水底にいて穴の中に潜入するものは円筒形を呈する。そのほか、球形や楕円(だえん)形をしたものなど生息場所に適応してさまざまな形がみられる。
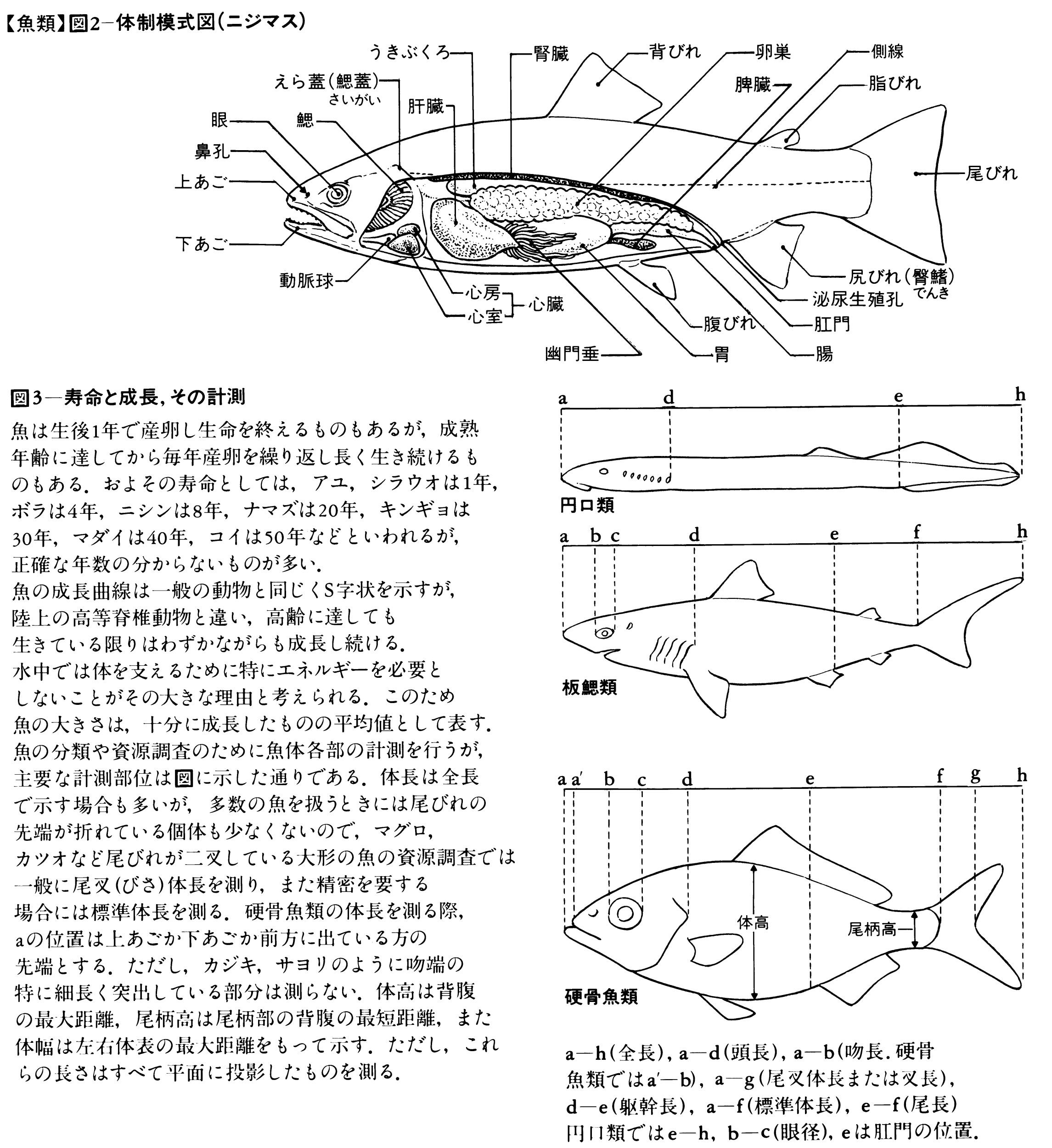
魚体の大きさは、次の三つの方法で表される。頭端から尾端までを全長total length、頭端から尾びれ後縁の切れ込みまでを尾叉長(びさちょう)または尾叉体長fork length、頭端から尾びれの付け根までを標準体長または単に体長standard length(SL)という。前二者は外部から簡単にわかるために漁業調査などで、後者は正確な長さを必要とする分類のときに用いることが多い。
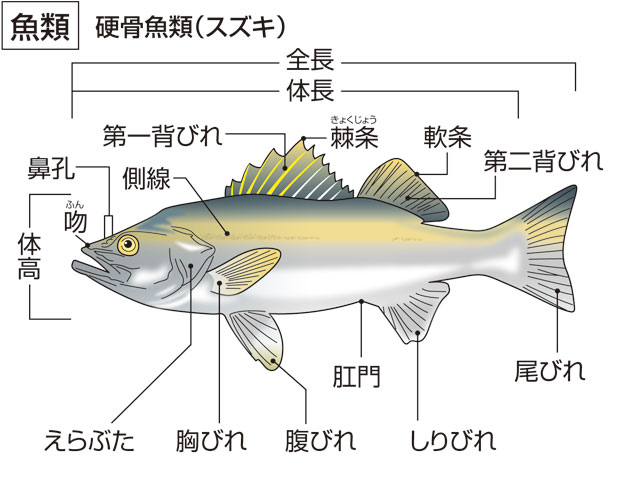
体は大別して頭部head、躯幹(くかん)(胴部)trunk、尾部caudalからなる。頭部は上顎の前端から最後の鰓孔または鰓蓋(さいがい)の後端部までをいい、肛門(こうもん)を境にして、それより前部を躯幹、後部を尾部という。頭部で目より前方を吻(ふん)snout、目より後背方を後頭部、目の下方を眼下部、えらを覆っている部分を鰓蓋部opercleという。躯幹部のうち胸びれの基部付近を肩部、その腹側面を胸部、さらに後方部を腹部とよぶ。尾部のうち背びれ、臀びれの後端付近から尾びれの付け根までを尾柄(びへい)caudal peduncleという。
[落合 明・尼岡邦夫 2015年3月19日]
体正中線上に位置する背びれ、尾びれ、臀びれを垂直鰭vertical finまたは無対鰭(むついき)unpaired finという。これらのひれは主として推進力を生み出し、体の左右安定を助ける働きをする。また、胸部と腹部に、それぞれ対(つい)をなした胸びれと腹びれがある。これらの対鰭paired finは、左右平衡や遊泳中の方向転換、遊泳停止などに役だつ。各ひれとも分節をした軟条rayや骨化した棘条spineで支えられており、これらを総称して鰭条fin rayという。各鰭条は薄い鰭膜によって連結されている。ひれの位置や構造は、魚の分類や進化を知るうえで重要である。とくに、腹びれの位置や鰭条数は有効であり、下等なグループでは後方にあって、多くの軟条からなり、棘条が未発達である。ひれは本来の運動器官としての機能以外に別の機能に進化した魚がみられる。たとえばアンコウ類では背びれ第1鰭条は誘引突起(ルアー)に、ホウボウ類では胸びれの下部の遊離軟条は歩行に、トビウオ類では胸びれは翼に、ハゼ類やクサウオ類では腹びれは吸盤に変化している。
[落合 明・尼岡邦夫 2015年3月19日]
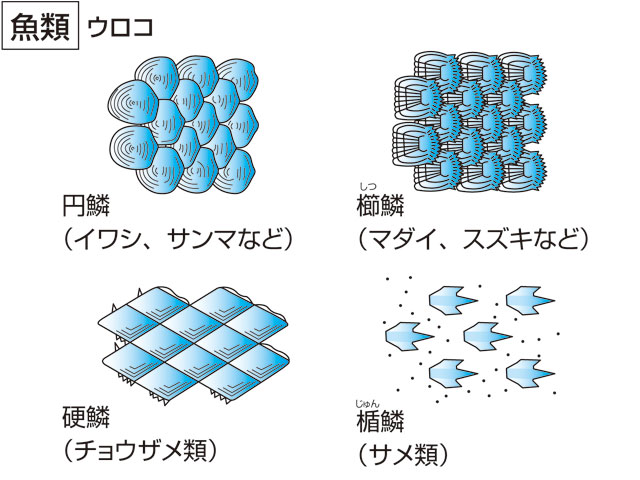
魚類の鱗は、皮膚の内層(真皮)にある細胞に起源がある。古生代の無顎類の体を覆っていた骨板から、鱗は2方向へ進化したといわれている。一つは軟骨魚類の楯鱗(じゅんりん)であり、もう一つは硬骨魚類にみられる鱗で、コズミン鱗(コスミン鱗ともいう)、硬鱗、円鱗および櫛鱗である。軟骨魚類にみられる楯鱗は、歯と同じ構造と発生をすることから皮歯ともいう。基底板から歯のように体表に突出した棘で、内側から髄、歯質およびエナメル層からなる。サメ類では体に密集して生えているが、エイ類では退化的である。ギンザメ類ではほとんどなくなり、側線に沿って埋まって残っているだけである。硬骨魚類にみられる鱗は基底部に板骨層(ばんこつそう)があり、その上が成分の異なる層で覆われている。コズミン鱗は厚くて、表面からエナメル層(あるいは硬歯質)、コズミン層、板骨層の3層からなる。おもに古生代の化石魚の総鰭類や肺魚類にみられる。現生の総鰭類のシーラカンスや肺魚類の鱗は薄くなっているが、コズミン鱗が退化したものと考えられている。硬鱗は表層を厚いガノイン層で覆われ、軟質類の化石魚、現生のポリプテルス、チョウザメ類、ガーパイクなどにみられる。ポリプテルスやガーパイクの鱗は瓦(かわら)の配列のように重ならず、隣り合う鱗は互いに連結している。普通の硬骨魚類では鱗は基底の繊維板層と表面の骨質層の2層からなり、薄くて瓦状に配列する。表面に棘のないものを円鱗、棘があり表面が粗雑なものを櫛鱗という。鱗は体を保護し、病原微生物の侵入を防ぎ、体の浸透圧調節をするのに役だつ。また、鱗の表面に環状に並んだ隆起線の粗密の配列によって、魚体の年齢を推定することができる。鱗の周囲から中央部の焦点に向かった放射状に走る溝を溝条(こうじょう)といい、その有無、数などは種によって異なる。体の中央部を水平に走る側線の上に1列に並んだ鱗を側線鱗lateral line scalesといい、その数は魚類の分類に重要である。
[落合 明・尼岡邦夫 2015年3月19日]
皮膚中に何種類もの色素胞があり、ホルモンや自律神経の作用を受けて、体色や斑紋(はんもん)が出現したり、変化したりする。黒色のものは黒(こく)色素胞(メラニンを含む)に、赤色は赤色素胞に、黄色は黄色素胞に由来する。また、黄色や赤色に関係したプテリンという色素胞もある。一方、グアニンやプリンなどを含んだ白色素胞(光彩細胞)があって、光の反射や屈折により、物理的にもいろいろな色が出される。なお、魚の縞(しま)模様は、頭部から尾部の方向を縦縞、背部から腹部の方向を横縞という。これは魚の頭を上に、尾部を下に立てたときの見え方による表現である。
[落合 明・尼岡邦夫 2015年3月19日]
体の各部やひれは骨格によって支えられる。骨格は脊索、軟骨、硬骨によって構成される。無顎類や軟骨魚類は前二者よりなり、硬骨魚類は三者からなるが、ほとんどが硬骨である。骨格系では頭骨、脊索および脊柱からなる中軸骨格と、胸びれや腹びれを支える肩帯と腰帯、背びれや臀びれの鰭条を支える担鰭骨(たんきこつ)からなる付属骨格に分かれる。さらに、頭骨は脳などを保護する神経頭蓋、いわゆる頭蓋骨と、両顎、舌弓(ぜつきゅう)、鰓弓を支持する内臓頭蓋に分かれる。さらにそれぞれは多くの骨のパーツで構成される。骨格の形や構造は化石となって長く残るうえ、種類によっても差異がみられるので、骨格は魚の進化や分類を研究する重要な資料となる。
[落合 明・尼岡邦夫 2015年3月19日]
筋肉は、口、えら、心臓や消化器官、ひれなどに発達するが、普通に魚肉とよばれるものは骨格筋であり、とくに、躯幹と尾部に規則的に配列する体側筋をさす。体側筋の伸縮によって推進力の多くが生み出される。筋肉は形態と生理的な特徴から赤色筋と白色筋に分かれる。赤色筋は筋繊維が細くて血管が多く走り、酸素を貯蔵する役目をするミオグロビンという色素が多い。マグロ、カツオなどの回遊魚にはとくに多く含まれる。また、体側の表層や中心部には暗赤色の筋肉があり、とくに血合肉(ちあいにく)(血合筋)とよばれる。赤色筋は多くのエネルギー源を含み、長距離の持続的な運動に耐えられる。一方、白色筋は筋繊維が太くて、ミオグロビンが少ないので、持続的な運動よりも瞬発的な力強い運動に向いている。一般的には海洋の表層に生息し、活発に遊泳している魚類では赤色筋がよく発達し、海底にすみ、動きの少ない魚類では赤色筋が少ない。また、魚類によっては絶えず動かしている胸びれは、瞬発的に動かす腹びれよりも赤色筋の割合が多い傾向にあることが知られている。
[落合 明・尼岡邦夫 2015年3月19日]
口腔から鰓腔、食道、胃、腸(後部を直腸という)、肛門までを消化管という。一方、捕食や消化・吸収作用を円滑にするため、肝臓、胆嚢(たんのう)、膵臓(すいぞう)などの消化腺(せん)と鰓耙(さいは)と歯がある。歯は上下両顎、口腔、咽頭、鰓腔などにある骨の上にあり、餌をとらえ、食べるために重要な働きをする。その形態、発達する場所などが食物によって大きく異なる。両顎のない無顎類では口腔と舌などに歯があるだけである。軟骨魚類のサメ類では3~5尖頭(せんとう)の歯をもつものが多いが、ホホジロザメやヨシキリザメなどでは鋭い歯に鋸歯縁(きょしえん)を備える。エイ類では両顎歯が敷石状に並ぶ。ギンザメ類は歯が癒合し歯板になっている。硬骨魚類の歯は食性によって著しく変化するが、次のように分けられている。牙(きば)状のものを犬歯状歯といい、動物を積極的にとらえて食べるタチウオ、ミズウオなどにみられる。錐(きり)状のものを円錐歯(えんすいし)といい、マイワシ、サケ類、タラ類など多くの種類にみられる。この歯の発達程度はさまざまで、多様な餌生物に適応している。微毛状のものを絨毛状歯(じゅうもうじょうし)といい、何列にも帯状に並んでいることが多い。ハゼ類に多くみられ、細かな餌生物をとらえて食べるのに適している。臼歯(きゅうし)は貝やカニなどの固い餌をかみ砕くイシダイ、マダイなどにみられる。門歯状歯はアオブダイ、フグ類などにみられ、岩礁などに付着している生物、サンゴ礁、植物性のものをかみ切るために適している。タツノオトシゴやヨウジウオ類には歯がない。歯は無顎類と軟骨・硬骨魚類で発生や構造が本質的に異なる。また、ヒトの歯と異なり、魚の歯は一生の間に必要に応じて何回でも生え替わる。
鰓耙は無顎類を除く魚類には、鰓弓の内側に2列に並ぶ。鰓耙は食物の濾過(ろか)器官である。軟骨魚類では一般に退化的であり、これを欠く種があるがプランクトン食のウバザメでは長くて密に生える。硬骨魚類では鰓耙は棒状、へら状、櫛(くし)状、歯状など魚種によってさまざまで、食性と密接に関係する。おもに細かな食物を食べるイワシ類などでは長く、数が多く、コノシロでは200本以上ある。大形の動物食のタチウオでは短くて歯状で、20本前後である。アンコウのようにまる飲み型では鰓耙がない。鰓耙の形態や数は魚類の分類に重要である。
胃は入り口の噴門(ふんもん)部、食物を収納する膨らんだ盲嚢(もうのう)部、腸につながる幽門(ゆうもん)部からなる。硬骨魚類では胃は種類によっていろいろな形がみられる。I型(フグ類)は盲嚢部がなく、まっすぐであるが、U型(ワカサギなど)は湾曲している。湾曲部にすこし盲嚢部ができたV型(サケ類など)、それが発達したY型(マイワシ、ウナギなど)からさらに発達したト型(マサバ、マダラなど)の5型に分かれる。胃は無顎類ではみられないが、軟骨魚類では基本的にはU型またはV型である。多くの硬骨魚類では、胃と腸の境目に盲管状の幽門垂があり、そこで餌を蓄えるとともに消化酵素を出して吸収作用も行う。幽門垂は種類によって大きさや数が異なる。イカナゴ類の小さい1本からマダラなどの300本以上のものまでさまざまであるが、コイ類やウナギではこれを欠いている。また、無顎類や軟骨魚類にはない。
腸は魚類では十二指腸、中腸、直腸の境界は明瞭(めいりょう)でない。腸は種類によって長さはさまざまである。また、成長段階によっても異なり、仔稚魚(しちぎょ)では直線的で短いが、成長するにつれて湾曲し、長くなる傾向にある。一般的に植物性の餌をとるものでは長く、腹腔内で複雑に回転している。軟骨魚類や下等な硬骨魚類の腸の内面には、吸収面を広くするため螺旋弁がある。軟骨魚類の腸の終末部に盲嚢の突起があり、直腸腺という。これには塩類の排出機能があるといわれている。
肝臓は消化腺としての胆汁を生成するが、栄養物の貯蔵場所であり、解毒作用も行う重要な器官で大きい。普通は左右の2葉からなる。胆汁が胆嚢に集められ、腸に送られる。胆嚢は緑色あるいは黄緑色で、卵形、紐(ひも)状などいろいろな形をしている。普通、肝臓と腸の間にあるが、肝臓に密着したり、その中に埋まったりしていることもある。
膵臓は軟骨魚類では腸の始部に1~2葉が認められるが、無顎類や多くの硬骨魚類では散在している。コイ、マダイ、ベラ、コチなどでは、肝臓内に入り込んでおり、肝膵臓とよばれる。
[落合 明・尼岡邦夫 2015年3月19日]
うきぶくろは、無顎類や軟骨魚類にはみられないが、多くの硬骨魚類では消化管と脊柱の間にあり、消化管の壁の一部が膨れたものである。細い管(気道)によって消化管と連結しているうきぶくろを有気管鰾(ゆうきかんひょう)、気道が消滅したものを無気管鰾という。イワシやコイなど下等なグループは有気管鰾であり、サバ、マダイ、ヒラメなど高等なグループは無気管鰾である。うきぶくろは、一般に卵形で1室であるが、2室または多くの小室からなるものもある。うきぶくろは普通、体の比重調節に用いるが、肺魚などでは毛細血管がよく発達して肺の役目をする。さらに、コイ類のように聴覚の補助に使用されるものもある。シマイサキ、ニベ類、タラ類などは、うきぶくろの外側に発達した筋肉の伸縮によって低周波の音を出して仲間どうし、また異性間の認知をする。マトウダイ類などではうきぶくろの内側の筋肉を伸縮させ、カサゴ類ではうきぶくろの前室と後室の間の隔膜を振動させて発音する。また、ホウボウ類はうきぶくろの外側と内側の筋肉を別々に振動させて2種類の音を出し、それぞれ求愛と警告に用いる。
[落合 明・尼岡邦夫 2015年3月19日]
心臓は胸部の腹側の囲心腔(いしんこう)の中にあり、静脈洞、1心房、1心室、および心臓球または動脈球からなる。軟骨魚類では心臓球、硬骨魚類では多くは心臓球が退縮し、動脈球が発達する。静脈洞と心房の間はくびれて、血液の逆流を防ぐ洞房弁があり、心房と心室の間もくびれて房室弁があり、心臓球の中に数列の弁がある。血液は、静脈洞から心房、心室、心臓球などを通って腹大動脈に入り、えらでガス交換をしたあと全身に供給される。体各部の末端で動脈血が酸素を放出し、老廃物を受け取った静脈血はしだいに集められて静脈洞へ運ばれる。したがって、心臓の中は静脈血だけが通る。ポンプの働きをする心室は著しく肥厚する。血液には赤血球、白血球、血小板などがある。血液は脾臓(ひぞう)、腎臓(じんぞう)、腸の粘膜下組織、肝臓などでつくられる。リンパ系もある程度整っていて、リンパ液が体内を循環する。
[落合 明・尼岡邦夫 2015年3月19日]
えらが主要な呼吸器官であり、ときにほかの器官を補助的に用いる。無顎類では多くの鰓嚢(さいのう)が鰓管の左右に対をなして並び、各鰓嚢の内面に多くの鰓弁がある。この類のヤツメウナギの仲間は各鰓嚢が小管によって外の鰓孔に通じる。この類では鰓孔が多く、7~15対ある。ヌタウナギの仲間は各小管が途中で合流して1個の鰓孔が開く。一般の魚では、5~7対の鰓弓の後縁に多数の糸状の鰓弁が前後2列に並ぶ。各鰓弁の表面には、直立した無数の二次鰓弁が1、2列に棚状に並び、ここに毛細血管がよく発達する。軟骨魚類では最後を除く各鰓弓に2列の鰓弁がある。鰓弁の間にある鰓隔膜が伸長して体表に達する。そのために鰓孔は5~7対ある。前後に分かれた鰓弁のことを片鰓とよぶ。多くの硬骨魚類では5対の鰓弓のうち前の4対に鰓弁がある。鰓隔膜は退縮して体表まで達しないので、1鰓孔として体表に開く。前後に分かれていない鰓弁を全鰓とよぶ。軟骨魚類のギンザメ類は鰓隔膜が退縮しているので1鰓孔である。
呼吸器官ではないが、鰓弓の前縁には突起物があり、それを鰓耙とよぶ。鰓耙は水を濾過して餌を漉(こ)し取る(前述の「消化器官」の章を参照)。
魚によっては補助的な呼吸器官がある。カムルチーには鰓腔上部にひだ状の粘膜よりなる上鰓器官が、キノボリウオには第1鰓弓の上皮から変化した迷路状器官があり、それによってガス交換をする。また、肺魚類ではうきぶくろが空気呼吸をつかさどる。ドジョウは空気を吸い込んで、腸の上皮で空気呼吸をする。皮膚でも多少の呼吸作用がなされる。
[落合 明・尼岡邦夫 2015年3月19日]
腎臓とえらで老廃物や余分な塩類を排出する。無顎類では中腎が、軟骨魚類や一部の下等な硬骨魚類では、中腎の発生に先だって前腎管が縦に二分裂し、その一つが輸尿管となる。サメ・エイ類の雄ではこの管は精巣とつながり、輸精管として働くので、生殖器官と密接な関係がある。硬骨魚類では腎臓は前端の多少肥大した頭腎と細長い中腎からなり、頭腎には尿細管がなく、排出は中腎でのみなされる。中腎はイワシ類のように左右が接合し、境界が明瞭でないもの、コイ類のように前半部で分離し、後半部で接合するもの、多くのスズキ類やカサゴ類のように後部だけが接合するもの、フグ類やアンコウ類のように完全に分離するものなど形状は多様である。輸尿管は左右1本ずつあり、それぞれ腎臓組織を縦走し、腎臓の後端で左右がまとまり1本となり腎臓で生成された尿を体外に排出する。輸尿管は無顎類では泌尿生殖洞に、軟骨魚類では総排出腔に、多くの硬骨魚類では泌尿孔または泌尿生殖孔へ開く。輸尿管の後端部が膨らみ膀胱(ぼうこう)を形成する種もある。なお、えらは呼吸作用のほか排出器官としても非常に重要である。
[落合 明・尼岡邦夫 2015年3月19日]
ホルモン物質を生産し、分泌する作用は、脳の前下方の視床下部に突出する脳下垂体で、前葉、中葉、および神経葉からなる。前葉では成長ホルモン、甲状腺刺激ホルモン、生殖腺刺激ホルモン、副腎刺激ホルモン、プロラクチンなどが分泌される。中葉からは黒色素胞刺激ホルモン、そして神経葉からは血圧上昇ホルモンなどが分泌される。成長ホルモンは成長を促進し、刺激ホルモンは甲状腺、生殖腺、副腎皮質などに働き、ホルモン活動を統制する。プロラクチンは浸透圧調節機能に関係する。
甲状腺は舌の基部の組織の中にある袋状の細胞の集まりで、形態や場所はさまざまである。甲状腺からヨードを含むホルモンを合成し、血液中に送り出し、変態、物質代謝、浸透圧調節、グアニン沈着、回遊促進などと密接な関係がある。
副腎は頭腎にあり、間腎腺とクロム親和細胞群からそれぞれ異なるホルモンが生産される。間腎腺はコルチゾール、コルチゾンなど、糖質、ナトリウムイオンやカリウムイオンの代謝に関係したホルモンを出す。クロム親和細胞群からはアドレナリンとノルアドレナリンが分泌され、血糖、血圧上昇、利尿などの作用に関与する。
ランゲルハンス島(膵島)は膵臓の中に埋没しているが、硬骨魚類のなかには独立した小体として胆嚢あるいは幽門垂付近にあるものもある。インスリン、グルカゴンというホルモンを出す。前者は血糖値を低くし、後者は高くする作用がある。精巣からはテストステロン、卵巣からはエストロンなどの性ホルモンが出され、ともに生殖腺を成熟させて、精子や卵形成を促進する。そのほかに硬骨魚類にだけみられる尾部下垂体が脊髄の末端腹面にあり、ナトリウムイオンの調節、利尿、血圧上昇に関与している物質を分泌する。
[落合 明・尼岡邦夫 2015年3月19日]
無顎類では生殖輸管(輸卵管、輸精管)は発達しないので、成熟卵、精子は直接体腔へ出て、生殖孔から体外へ放出される。サメ・エイ類と真骨魚類では生殖輸管の発生過程や形態に大きな違いがある。サメ・エイ類では輸卵管は卵巣に直接結び付いておらず、前端は卵の受け口として体腔へ開いている。体腔へ排卵された成熟卵はここから左右の輸卵管に入る。輸卵管の前部には卵の殻をつくる卵殻腺があって、その成分を分泌する。卵生種では大きく、胎生種では小さい。交尾で輸卵管に入った精子はここで成熟卵を待って、受精する。胎生魚では輸卵管は子宮の役割をする。輸卵管の後端は総排出腔に開く。雄では精巣でつくられた精子は輸精管を経て中腎管へ入り、輸尿管を経て総排出腔へ送られ、交尾で雌の輸卵管へ送られる。交尾器の基部にはサイフォンサックsiphon sacがあって、ポンプの働きで精液は交尾器の溝を経て、子宮内へ注入される。真骨魚類で、嚢状の卵巣をもつ種では、輸卵管は卵巣壁が延びて管状になったもので、前端は卵巣腔に開き、後端は生殖孔として肛門に開く。雄では精巣から成熟した精子が輸精管に入り、生殖孔から精液として放出される。サケ類では輸卵管は退化し、成熟卵は腹腔へ出て、肛門直後の生殖孔から産出される。雄では精液は腹腔へ流出し、生殖孔から放出される。
[尼岡邦夫 2015年3月19日]
魚類の大部分は卵生であるが、少数のものは卵胎生または胎生によって繁殖する。卵は硬骨魚類やヤツメウナギ類では小さくて直径1ミリメートル前後にすぎない。1尾がはらむ卵の数は、普通数千粒から数百万粒、なかには数千万粒以上のものもある。一方、ヌタウナギ類や軟骨魚類の卵は大きく、堅い卵殻に包まれている。ヌタウナギ類では20~44個ほどの卵を産む。軟骨魚類は体内受精により卵または胎仔(たいし)を産む。ネコザメ科、トラザメ科、ジンベエザメ科、ガンギエイ科などは卵生であるが、ほかの多くの軟骨魚類は雌の体内でよく発育した胎仔を出産する。栄養は卵黄だけに依存する種から、雌からなんらかの栄養が供給されるものまでいろいろなタイプがみられる。胎仔数は種類や体長によって変化するが、10尾以下から80尾以上のものまである。妊娠期間は多くは10か月ほどである。硬骨魚類では、メバル、カサゴ、ムラソイなどのカサゴ類、カダヤシ、ヨツメウオ、ウミタナゴ類、ある種のサヨリ類、ゲンゲ類、イタチウオ類、アシロ類、シーラカンスなどが胎仔を産む。カサゴ類では孵化(ふか)後まもない仔魚(しぎょ)の状態で産み出されるが、多くの種は体内で栄養の補給を受け、稚魚になって出産される。栄養の補給の方法は種類によって異なる。妊娠期間は普通数か月、長いものでは1年またはそれ以上に及ぶ。
受精は卵生魚では水中で、胎仔を出産する魚類では卵巣か輸卵管(子宮)の中でなされる。しかし、卵生の軟骨魚類では受精は交尾によって雌の体内で行われ、受精後まもなく水中へ産み出される。硬骨魚類の多くは、卵の一極(動物極)に径3、4マイクロメートル程度の細い孔(あな)(卵門)があり、ここから1個の精子が卵内に入って卵核と合体する。ただし、チョウザメ類は複数の卵門があるが、自然状態では1個の精子だけが卵内に入る。ヤツメウナギ類や軟骨魚類では卵膜に特別な孔がなく、精子から出されるライシンで卵膜の一部を溶かして小孔をつくる。軟骨魚類では1個の卵に多数の精子が入り込むが、そのうちの1個だけが卵核と合体し、そのほかの精子は卵黄の多い部分へ移動して、胚(はい)の形成には関与しない。また、ごく限られた魚類では雌性発生をする。ギンブナや熱帯魚のポエシリア属Poeciliaやポエシロプシス属Poecilopsisのある種類では真の受精がなされず、ほかの種類の精子の刺激によって発生が進み、卵の核だけで胚となる。したがって精子は遺伝に関与しない。このような魚類では、雌だけで、母親と同じ遺伝形質をもった子孫を何代にもわたって産む。
受精した卵は卵割を始める。ヤツメウナギ類、肺魚、チョウザメ類、アミア、鱗骨魚などは、全割holoblastic cleavageといわれるもので、卵割は卵の全体でなされる。しかし、ヌタウナギ類、軟骨魚類、多くの硬骨魚類では、部分割meroblastic cleavageといって動物極の部分に胚盤がつくられ、この部分だけが分割する。硬骨魚類では孵化に先だって、胚体のある部位で孵化腺がつくられ、そこから分泌されるタンパク質分解酵素は卵膜表面の薄い膜を残して、硬タンパク質の内層を化学的に分解させる。これに加えて胚体の口や鰓蓋の開閉などによる機械的な運動によって残った卵膜が破壊される。ナヌカザメ属の1種Cephaloscyllium ventriosumなど卵生の軟骨魚類では、胚体の背面に2縦列の歯状突起が生じ、孵化後に退化する。これは孵化のときに歯車のように動いて、堅い卵殻を破る働きをすると考えられている。受精から孵化までの日数は、種類によって著しく異なり、早いものでは1日以内、遅いものでは2か月以上にもなるが、一般に数日から1週間ぐらいである。卵生魚では卵は水中で発生・孵化するので、水温、塩分濃度、溶存酸素量などの影響を受ける。そのうちもっとも強い影響を受けるのは水温で、高すぎても低すぎても正常な発生を妨げる。孵化の適温は北方種と南方種では異なるし、同海域でも魚種によって違っている。ニシン、ワカサギ、タラ類、カレイ類は数℃前後、マダイ、ヒラメ、アイナメなどは15℃前後、サバ類、マアジ、ブリなどは18℃前後、コイやフナは20℃、カツオ・マグロ類、熱帯魚では22℃以上である。一般的に、適温の範囲内では温度が高いほど孵化するまでの時間が短くなる。また、卵の発生には一定量の温度が必要とされ、完了までに必要とする時間と平均有効温度の積はほとんど一定である。これを積算温度total effective temperatureとよび、「D°」で表される。たとえばサケの場合、受精から孵化までの平均有効温度は8.3℃で孵化での日数は58日であるので有効積算温度は481.4D°となる。
卵発生に適した塩分濃度はその種の産卵水域によって異なるが、水温のような強い影響を与えない。適塩分濃度を外れた場合の孵化率は低く、孵化しても仔魚はすぐに死ぬことが知られている。溶存酸素量は低下すると、発生が遅れたり、奇形が産まれたり、孵化率が低下したりする。
人工的に大量生産した幼生を、増養殖の種苗として利用するようになり、アユやサケ・マス類、マダイ、ヒラメ、クロダイ、トラフグ、マアジ、クロマグロなど相当数の種の種苗が生産されている。このためホルモンの投与、日照時間や水温調節などによって成熟を促進したり、人工受精や水槽内での自然産卵などに関する技術が開発されたりしている。ニホンウナギのように、種の保存が厳しく求められ、種苗生産のさらなる研究開発が望まれている種もある。
[落合 明・尼岡邦夫 2015年3月19日]
生殖行動は魚種によって多様性に富んでいる。体内受精をする魚類は雌雄が対になり交尾をする。体外受精をする魚類は雌雄が対になるものから集団をつくるものまでさまざまである。普通、分離浮性卵を生む魚類は海面近くで生殖行動をする。マイワシでは日没後に雌雄が群れで海表面に集まり、産卵・放精をする。マダイでは数尾の雄が雌を追って海面に追い上げて、そこで産卵・放精をする。ベラやブダイでは沖に向かう潮流がおこる時間にあわせて産卵が行われるが、卵を分散させることや食べられることを避けるためであると考えられている。
沈性卵や付着卵を産む魚類は雄雌で産卵行動をとる種が多い。サケでは雌がひれで川底を掘って産卵床をつくり、雌雄が対になって産卵・放精をし、雌が卵の上に砂利をかぶせる。
ハゼ類は海底の穴や貝殻などに産卵巣を設ける。イトヨは河川、池、湖の底に水草などで産卵巣をつくる。スズメダイでは雄が産卵期に縄張りをもち、岩の表面を歯で掃除して、産卵床をつくる。雄は求愛行動をして雌を誘い込み、産卵・放精が行われた後に雄が卵塊に水流を送り、卵を保護したり、外敵から守ったりする。タナゴ類では雌が産卵管をカラスガイなどの出水管に挿入して鰓腔に産卵し、雄は放精し、受精させる。テンジクダイ科のテンジクダイやネンブツダイの雄は卵を口内にくわえて口内保育mouth broodingをする。カワスズメ科(シクリッド類)では、いろいろな口内保育のタイプがみられる。雌が口内保育をする種がもっとも多いが、雌から雄に途中で交代する種がいる。なかには托卵(たくらん)されたナマズ類の卵をいっしょに口内保育する種がいる。口内保育はいずれも卵や子供を敵から守るためであり、口内保育をしないで見張りをする種より卵や子供の数が少ない。
[尼岡邦夫 2015年3月19日]
孵化後の仔魚は、ただちに、またはしばらくして餌をとり、前期仔魚prelarva、後期仔魚postlarva、稚魚juvenile、若魚(わかうお)young(もしくはadolescent)、未成魚immatureの各期を経て成魚adult、老魚senescentとなる。前期仔魚期は孵化から卵黄が吸収されるまでの期間である。腹部に卵黄を入れた袋があり、体は透明で、背・腹正中線上に1枚の薄い膜状のひれがある。卵黄を吸収してから各ひれが完成して、各ひれの鰭条が定数になる、背骨などの内部骨格が骨化するまでを後期仔魚期という。この間は主としてプランクトンを食べて変態するものが多い。ウナギやアナゴなどは細長い葉状を呈するので、とくに葉形幼生(レプトセファルスleptocephalus)という。カレイ・ヒラメ類では、この期の終わりごろに一つの目が他側に移動し、体の構造が左右不相称となる。海水魚では多くの種類で頭部や体部に骨質突起が一時的に出現する。体を浮遊させるのに役だてるためといわれている。特異な突起をもつ幼魚には特別の名前が与えられている。イットウダイ類のリンキクチスrhynchichthys幼生、アマシイラのヒストリチネラhystricinella幼生、チョウチョウウオ類のトリクチスtholichthys幼生などである。後期仔魚から稚魚になる間に著しい形態変化がおこる。このことを変態metamorphosisとよぶ。稚魚期には体形がほぼその種類の特徴を現し、体色も成魚に似てくる。この時期には消化器官が完成して本来の餌を食べるようになり、成長や運動能力が著しく促進される。若魚期には形態的に種の特徴が完備し、行動が活発になるが、二次性徴はまだ現れていない。未成魚期は体つきが成魚に近いが、性的には未成熟で、生殖能力は不十分である。その後、生殖腺が発達して成魚期になる。生活の機能や生殖能力が衰え、体つきが変わり、老魚期になる。
サケ・マス類の子供は特別の名前でよばれる。孵化後の大きな卵黄を備えたものをアレビンalevin、卵黄を吸収して摂餌活動を始めるものをフライfry、回遊生活前のパーマークを備えたものをパーparr、体が銀色(銀毛)になり、回遊生活に入ったものをスモルトsmolt、回遊生活を1年ほど過ごしたものをウィトリングwhitlingとよぶ。
身近にいる魚で成長段階によって呼び名が変わるものを出世魚という。ボラ、スズキ、ブリなどがその代表である。稚魚、若魚、未成魚、成魚といった各段階に対応して、生息場所や生態、漁法、経済的価値などが変わるため、呼び分けられるようになった。呼び名は地方によって多少の差がある。
成魚になる年齢は魚種によって著しく異なる。多くのものでは生後2年から数年であるが、アユやメダカなどは1年以内、ウグイ、イワシ類など小形の魚類は1年、チョウザメやオヒョウ類は長く、10年前後である。アユ、ウナギ、遡河(そか)性のサケ・マス類などは一生に1回産卵して死亡するが、多くの魚類では一生に何回か産卵する。魚の寿命は短いもので1年以内のものがあるが、多くは数年から10年である。しかし、コイやマダイなどは15年以上、チョウザメ類は100年余りも生きる。
[落合 明・尼岡邦夫 2015年3月19日]
種類により主食とする餌は著しく異なり、その構成には種的特徴がみられ、それを食性という。もっとも多いのはプランクトンと動物肉である。淡水魚ではゲンゴロウブナ、ニゴロブナ、レンギョ(コクレン、ハクレン)、内湾や入り江などではコノシロ、イワシ類、キビナゴ、イカナゴ、磯魚(いそうお)ではテンジクダイ類、ハタンポ類、ニザダイ類、沖合いや外洋ではサンマ、トビウオ類、ウバザメ、ジンベエザメ、深海魚ではハダカイワシ類などが一生プランクトンを捕食する。プランクトン食魚では、鰓耙が長くて数が多く、これで水とともに飲み込んだ餌を濾過する。普通、食性は肉食性carnivorous、植食性herbivorous、および雑食性omnivorousと分けられる。肉食性の魚類は、昆虫、魚類、甲殻類、軟体動物、環形動物、棘皮(きょくひ)動物などを主食または混食する。淡水域のカムルチー、海洋でのマグロ類、カジキ類、サメ類、ハモ、ヒラメなどは魚食魚の代表であり、食物連鎖ではそれぞれの生態系で最高の段階にある。肉食魚は一般に口が大きく、歯がよく発達しており、鰓耙は少なく退化的であり、腸も短い。
藻類や水草、陸草などを主食にして成長する魚は植食性で、これを草食魚という。淡水では、岩石に付着する藻類を主食とするアユやアジメドジョウ、ある種のティラピア、陸草や水草を主食とするソウギョなどがそれである。海水魚でも、ある種のニザダイ、ブダイ、アイゴ、メジナなどは冬に藻類を主食にする。動物と植物を区別せずに混食するものは雑食性である。淡水では、昆虫と藻類を食べるウグイ、オイカワ、ドジョウ、コイ、キンブナなど、海では、魚類、甲殻類、多毛類、海藻類を混食するある種のサヨリ類、ベラ類、フグ類などが雑食魚である。そのほかに変わった食性をもったものがみられる。サンゴ礁にすむチョウチョウウオ類やスズメダイ類の仲間はサンゴのポリプから分泌される粘液を食べる。アフリカのタンガニカ湖にすむシクリッド類はほかの魚の鱗を食べる。ホンソメワケベラ、メジナの仲間などは、ほかの魚の体表の寄生虫を食べる。
養魚では、プランクトン食のサバヒーやティラピアなどは、施肥などにより人工的にプランクトンを繁殖させて飼育することもある。これに対して、雑食魚や肉食魚の養殖には、餌料(じりょう)代が経営に大きな割合を占めるので、植物性の割合を高くしたり、人工的に合成した餌料で成果をあげたりしている。また、釣り餌はそれぞれの魚類の好みに応じて選択され、肉食魚では嗅覚(きゅうかく)や視覚をよく刺激するような環形動物、エビ類、貝類、小魚などを用いる。
[落合 明・尼岡邦夫 2015年3月19日]
魚類でも性は遺伝的な支配を受けるが、二次的に環境や生理的な条件で転換することがある。自然の状態で一生の間に雄から雌、雌から雄へと性転換する。魚類の性染色体は一般に雌がホモの場合はXX‐XY型(ときにXX‐XO型)、雌がヘテロではWZ‐ZZ型またはWY‐YY型である。しかし、性が発現するときには常染色体上の性支配因子によっても影響され、これが性染色体の性支配因子より優勢になることがある。したがって、魚の性の発現はほかの高等動物より複雑である。
魚類は雌雄異体であるが、なかには卵巣組織と精巣組織を有する両性生殖腺を有し、雌雄同体hermaphroditismとなるものがある。ある種のカワヤツメ類は未成熟期に44%が雌雄同体であり、成熟するとすべて雄になる。真骨魚類ではアオメエソ類、ミズウオ類、ハタ類、タイ類、ベラ類、チョウチョウウオ類、スズメダイ類などかなり多くの種にこの現象がみられる。メキシコ湾のハタの仲間のセラヌスSerranus subligariusという磯魚やメダカ目のある種では、一個体で卵巣や精巣が同時に成熟し、他家受精のほかに、ときには自家受精する。雌雄同体の魚は性転換を行うものが多い。初めに精巣が発達して雄として機能し、のちに精巣が退化して卵巣が発達して卵を産むものがある。これを雄性先熟protandryといい、ヨコエソ、オニハダカ、クロダイ、ヘダイ、クマノミ類、多くのコチ類などがその代表である。クマノミ類は1尾の大きな雌が中心となり、すこし小型の成熟雄と数尾の未成熟魚がイソギンチャクと社会構造を形成し、生活する。雌が死んだり、いなくなったりすると、そのなかの1尾の雄が性転換して雌になる。雄性先熟とは逆に、初めに卵巣が成熟し、のちに精巣が発達する雌性先熟protogynyの魚には、ソードテール、タウナギ、ハタ類、ハナダイ類、キダイ、ベラ類、チョウチョウウオ類がある。性転換に伴って体色が雌から雄に変化し、1尾の雄が縄張りをつくり、数尾の雌と未成魚によるグループで、雌が雄に性転換することをコントロールしている。ベラ類のホンソメワケベラでは1尾の雄が数尾の雌と数尾の未成魚で1グループをつくる。雄が死ぬと、もっとも大きな雌が性転換して雄になり、このグループを支配する。雄の死後30分で雄的な威嚇行動を、1日後に求愛行動を示し、2週間ほど経つと生殖活動が可能となる。一般に、運動性が大きく、群れをつくって回遊をするものには雌雄同体現象が認められない。狭い水域にすみ、環境要因や餌物(えもの)などが不安定な淡水魚、熱帯や亜熱帯域で社会的なグループをつくる熱帯魚、磯魚、深海魚に性転換が多くみられる。
魚の性は人工的に転換させることができる。性ホルモンを幼生期に餌とともに2、3か月連続して与えると、遺伝型と異なった性に変えられる。雌化にはエストロゲンを、雄化にはアンドロゲンを用いる。実験的にはメダカ、ドジョウ、グッピー、ティラピア、サケ・マス類などで成功している。ウナギやドジョウなどのように雌雄によって成長が異なる場合には、成長のよい性だけを養殖したほうが経済成果が大きい。また、ティラピアでは繁殖力が非常に旺盛(おうせい)であり、雌雄を混合して養殖すると過密になって小形化し、全体の収量が減少してしまう。それで雌を雄にし、成長のよい雄だけを養殖するようくふうされている。
[落合 明・尼岡邦夫 2015年3月19日]
魚類は、光を主として目で受容するが、両眼の間の頭頂にある上生体(松果体)からも光を感ずる。魚類の目はレンズが球形であり、レンズの下方に付着する筋肉の働きで、レンズを前方または後方に移動させて像を網膜に結ぶように遠近調節する。視力は明るい所でも0.5以下と弱いが、動く物体を感知する能力はヒトと同様か、またはその倍もある。受容する光の波長は400~750ナノメートルで、もっとも敏感に感ずる波長は、淡水魚では平均して620ナノメートル、海水魚では500~550ナノメートルである。一般的に色覚は淡水魚や岸辺の魚では発達しているが、深海魚や外洋の表層魚では退化して色の区別ができないものもある。上生体には網膜と同じような光を受容する細胞があり、日照時間を感知する能力がある。暗黒の洞穴の中には目がなくなった魚がいる。深海では、光が少ないところでは、マトウダイ類は光を受容するために目が大きくなり、ソコオクメウオ類、クジラウオ類などでは目が小さくなっている。なかにはチョウチンハダカのように目が退化してなくなっているものもある。深海魚にはボウエンギョやデメニギスのように望遠鏡のような筒状の目、ヨツメニギスやテンガンムネエソのように上を向いている目など特殊化した目をもったものがいる。
[落合 明・尼岡邦夫 2015年3月19日]
嗅覚は底生魚や夜行性の魚類でよく発達している。鼻腔の中ににおいを感受する細胞が集まった嗅房(きゅうぼう)がある。普通、前の鼻孔から入った水は嗅房を通って後鼻孔から体外へ流れ出る。嗅房で水に含まれる化学的成分が嗅細胞を刺激し、これが嗅神経や嗅球を経て中枢部の端脳に伝えられる。嗅覚は餌を探すほか、サケ・マス類は降海直前にすり込まれた生まれた川(母川(ぼせん))の水のにおいを記憶していて産卵のために生まれた河川に戻ってくる。ナマズなどでは、同種間で出し合うにおい物質によって仲間どうしが群れをつくる。コイ類やナマズ類では外傷のある魚体から出される恐怖物質にあうと、群れを崩して跳びはねるなど異常な行動をして外敵からの難を避ける。遡上中のサケ・マス類に上流から哺乳(ほにゅう)類の皮膚のにおいが流れると遡上が一時止まる。岩の割れ目に潜んでいるウグイに動物模様の布きれを近づけても反応しないが、イタチの皮では驚いて飛び出してくる。視覚よりも嗅覚に敏感に反応する。イソギンポ類やナマズ類、グッピー、アユなどは、産卵期に異性から出されるにおい物質(性フェロモン)に誘引され雌雄が集まり産卵する。キンギョでは排卵された卵をもつ雌を追尾し生殖孔近くをつつくが、鼻孔に栓をしたり、嗅神経を切ったりすると排卵雌とそうでない雌を区別できない。
[落合 明・尼岡邦夫 2015年3月19日]
甘味、塩味、苦味、酸味などは味蕾(みらい)で受容される。多くの種類では特殊なアミノ酸や核酸関連物によく反応し、10-10モル程度の低い濃度にも反応する。味蕾は普通、洋ナシのような形をしていて、表皮中にあり、先端は表面に出ている。その基底部は下層の真皮層から表皮層へ突出している棒状の台に接している。味蕾は感覚細胞、支持細胞および基底細胞からなり、各味蕾は顔面神経あるいは迷走神経の末梢(まっしょう)につながっているが、味覚の中枢は延髄にある。味蕾には口腔、鰓腔、咽頭、食道などにある味蕾と口唇、体表、ひれなどの表皮に分布する皮膚味蕾に分けられる。味蕾の分布は摂餌生態と密接に関係する。吸い込み型の摂餌をするコイ、フナ、キンギョなどでは口蓋部に密に分布している。コイ、ドジョウ、ナマズ、ヒメジ、マダラなどのひげには味蕾があり、水底の餌を探すのに利用している。ホウボウ類の胸びれの下部の遊離鰭条には感覚細胞が分布し、海底に潜む餌生物を探し出す。
[落合 明・尼岡邦夫 2015年3月19日]
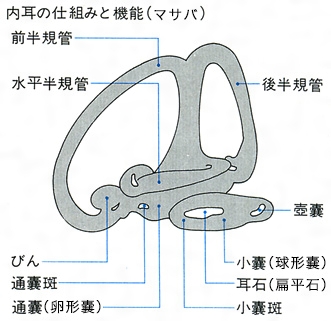
音の受容器には内耳と側線があり、内耳は聴覚と平衡感覚に関与し、側線は機械的な刺激や化学的刺激を感受する。耳は内耳だけで、頭蓋骨の耳殻の中に左右1対ある。外耳や中耳はなく、内耳は外部から見ることはできない。内耳の形態は種類によって著しく異なる。内耳は上部と下部の2部に分かれる。上部には通嚢(卵形嚢ともいう)と前、後、水平の三半規管からなり、下部は小嚢(球形嚢)と壺嚢(つぼのう)からなる。三半規管の一端は膨らみ、瓶ampullaを形成する。この内面には感覚上皮があり、多くの有毛細胞をもつ。通嚢、小嚢および壺嚢の内壁には平衡斑とよばれる感覚上皮があり、総称して耳石器とよぶ。各嚢の平衡斑の表面に耳石が接している。通嚢の耳石を礫石(れきせき)、小嚢のものを扁平石、壺嚢のものを星状石といい、一般に扁平石がもっとも大きく、単に耳石というときにはこれをさす。耳石はおもに炭酸カルシウムで形成される。硬骨魚類では、扁平石の形は魚種によって異なり、矢じり形、楕円形などさまざまで種の特徴になることもある。一般的に、遊泳魚では小さく、マサバでは2ミリメートルであるが、底生魚では大きい傾向にあり、マダラでは10ミリメートルほどである。また、平衡石は炭酸カルシウムの沈着の周期性からくる輪状帯を利用して年齢・日齢査定に利用される。サメ・エイ類には硬い耳石がないが、各嚢には粒状の炭酸カルシウムを含む平衡砂がある。各嚢や三半規管の内腔はリンパ液で満たされ、わずかな振動で液が流動し、有毛細胞が刺激を受けて、音の強さ、周波数、方向などの信号を分析している。音の受容にはおもに通嚢と壺嚢が関係している。また、体の姿勢制御には三半規管、耳石器が関係しているが、多くの魚では通嚢も働いている。可聴範囲は魚種によって異なるが、10ヘルツから3000ヘルツの範囲であるといわれている。うきぶくろは、音波を受容したとき、膨れたり収縮したりするので、うきぶくろが中耳のような役目をする。とくに、過去に骨鰾類(こっぴょうるい)ともいわれていたように、コイ科やナマズ類では、うきぶくろと内耳が骨片で連結し、哺乳類の内耳のような構造になっている。このため聴覚は敏感で1万6000ヘルツまで受容する。
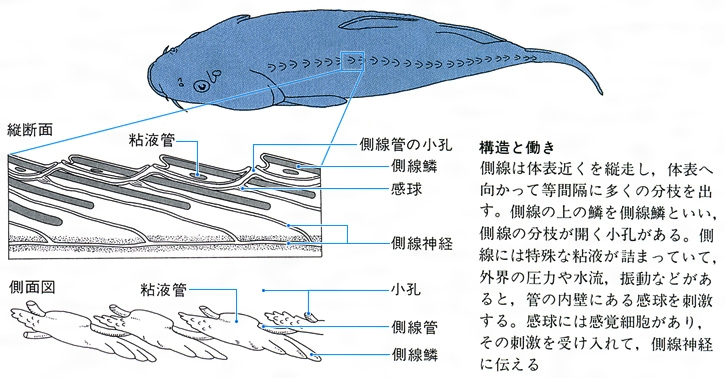
側線は体の側面と頭部にあり、体表に点在する孔器と側線管内に並ぶ管器とに分かれ、いずれも感丘が受容器になっている。感丘は有毛細胞と感丘の表面を覆う寒天質のクプラをもった感覚細胞が分布したもので、基本的な構造は耳と同一である。仲間や餌生物、捕食者などの動きから生じる水量の変化を感知することができる。
[落合 明・尼岡邦夫 2015年3月19日]
魚類のなかには鰾(うきぶくろ)、上鰓規管、腸などで補助的に空気呼吸するものもあるが、ほとんどの魚類は水中でえら呼吸をする。空中に比べて水中では利用できる酸素が少なく、また、水中に拡散する速度がきわめて遅いために空気呼吸に比べて著しく悪い環境にある。たとえば水温15℃の海水で6ミリリットル、淡水で7ミリリットル以下である。しかしこれを克服するために効率のよいえらを備え、また、絶えず水流をおこして新鮮な水が流れるような装置をもっている。魚類のえらは、周囲の水中の酸素量の80%ぐらいまで吸収する。多くの空中または水中動物では、呼吸器の酸素の利用率は20~30%程度であるので、魚類のえらは呼吸器官として非常に優れている。
えらには普通5対(ときには6~7対)の鰓弓があり、前4対に鰓弁がある。鰓弓の外側に前後2列に並ぶ櫛状の無数の鰓弁を備える。サメ・エイ類では前後の鰓弁の間に鰓隔膜がよく発達し、前と後の鰓弁を分離している(片鰓)。硬骨魚類では鰓隔膜が退縮し、前後の鰓弁は分離しない(全鰓)。無顎類には鰓弓はなく、鰓弁は各鰓嚢のなかに並ぶ。魚類の各鰓弁は両側面にさらに半円形、三角形などいろいろな形の多数の二次鰓弁が並ぶ。二次鰓弁は薄く、その上皮細胞は薄い単層で、その下に毛細血管が網目状に並び、水と血液との距離を少なくしている。この構造がガス交換の効率をよくしている。さらに鰓弁は数百から数千もあり、鰓弁の長さ1ミリメートル当り数個~30個余りの二次鰓弁がある。したがって二次鰓弁の総面積は体表の数倍にも達する。一般に鰓総面積はカツオ、マグロなどの遊泳性魚類では広く、カレイ、アンコウなどの底生性魚類では狭い。また、一般に活発に泳ぐ魚ほど、小さく、密に並ぶ二次鰓弁を備えた鰓弁を多数もっている。二次鰓弁の表面の厚さは魚種によって異なるが、表層を活発に泳ぐカツオ、マサバ、ビンナガではきわめて薄く0.2~3.6マイクロメートルであるが、あまり遊泳しないカレイなどの底生魚では0.9~44.3マイクロメートルと厚い傾向にある。さらに、呼吸効率を高めているものに、二次鰓弁の毛細血管の血液の流れが水流に対向する仕組みがある。口から入った水は各二次鰓弁の周りを内から外に向かって流れる。一方、鰓弁に入った動脈から二次鰓弁内の毛細血管に流れ込んだ血液は、水流と対向して流れる。このような対向流システムはガス交換の能率を大きくしている。
えらに新鮮な水を取り込むシステムとして、口腔の加圧ポンプと鰓腔内の吸引ポンプがあり、うまく連動して働く。それには口唇の上下にある口腔弁や鰓蓋の後縁にある鰓蓋弁が重要な役割を果たす。口と鰓孔が腹面に開く底性のエイ類では頭部背面に大きな呼吸孔spiracleがあり、呼吸水を取り込むことができる。
酸素不足症で死亡する原因に、環境、水の汚染、毒物や病害虫によるえら組織の異状化、ヘモグロビン不足による貧血などがある。養魚では水中の酸素不足をおこさないくふうが施されている。ウナギ、コイなどを止水状態で飼育するとき、良質の植物プランクトンを繁殖させ、その光合成の結果出された酸素を利用するようにしている。コイを流水状態で飼育して、多くの酸素を供給すると、1平方メートル当り200キログラムの生産があがる。この量は止水の場合の200~400倍にあたる。
[落合 明・尼岡邦夫 2015年3月19日]
うきぶくろは無顎類と軟骨魚類にはないが、多くの硬骨魚類には体腔の脊柱と消化管の間にある。うきぶくろの起源は消化管の腹面から膨らんだ対状の呼吸器官であったが、真骨魚類では腹面から側面や背面に移動し、内部にガスを満たし、機能は浮力の調節器官に変化した。うきぶくろのない魚類では体の比重はおよそ1.1に近いが、うきぶくろのある淡水魚ではうきぶくろの容積が体のおよそ5.7%以上であるのに対し、海水魚では5.7%以下でそれぞれ比重がおよそ1になり、つり合う状態になる。
うきぶくろの中のガスは、酸素、窒素、二酸化炭素、希ガスなどからなる。それらの組成は、普通、空気と著しく異なっている。酸素は淡水魚では10%前後、海水魚では多く、とくに深海魚では80~90%もある。窒素は酸素とは反対に、淡水魚で80%以上、深海魚で10%余りである。二酸化炭素は多くても数%にすぎない。海では10メートル深くなるごとに1気圧ずつ増えるので、うきぶくろの中のガスの分圧も深海魚では大きい。
マイワシ、コイ、ウナギなどで、うきぶくろが食道に通じている魚(有気管鰾)では、ガスは口から吸い込んでうきぶくろの中に入れる。スズキ、マダイ、マダラなどではうきぶくろが独立したもの(無気管鰾)では、特別なポンプの仕組みで、うきぶくろへ連なった動脈血との間でガスの取り入れや放出がなされる。うきぶくろの一部に、異常に発達した無数の動脈と静脈の毛細血管が並行して走っている。この部位を奇網rete mirabileといい、ここでも動脈血と静脈血が対向的に走り、動脈血内のガスの分圧が効果的に高められる。奇網の先端にあるガス腺gas glandでは、乳酸などによって血液中の水素イオン濃度(pH)が変わり、ガスの親和性や溶解度合いが減少し、血液からうきぶくろの中へガスが出される。うきぶくろのガスを減らす場合には、うきぶくろに通じた特殊な管を通じて体外に出されることもあるが、うきぶくろの後部に発達した卵円体oval bodyの毛細血管へ集められる仕組みになっている。一般的にいえば、水深200メートルより浅いところにすむ魚ではうきぶくろをもつ種がもっとも多いが、200~1000メートルでは少なくなり、1000メートルより深いところではなくなる。うきぶくろのない深海魚は体側筋が退縮し、水分と脂質の含有量が増し、タンパク質と灰分が少なくなり、体は軽くなっている。深所と浅所の間を鉛直回遊するハダカイワシ類などは体の水分は多く、体比重は1.04で、わずかに重いが、うきぶくろを退縮させ、ガスを脂質で置き換えることで、水圧による影響を少なくして浮力調節ができる。
[落合 明・尼岡邦夫 2015年3月19日]
魚類は水中にいるため、絶えず体の浸透圧を調節する必要がある。淡水魚の体の浸透圧は周りの水よりはるかに高いので、絶えずえらから水分が多量に入り込む。それで多量の希薄な尿を体外に排出する。その量は体重1キログラム当り1日に48~60ミリリットルほどである。一方、海水魚は浸透圧にして体液の3倍も高い海水中に生息する。体内に水分を保持する必要上、積極的に海水を口やえらから取り入れ、排尿量を少なくしている。その尿量は1キログラム当り1日に0.4~5.0ミリリットルである。つまり、体液の浸透圧を一定に保つために淡水魚では水ぶくれにならないように水を飲まないようにし、排尿量を増やす。海水魚では脱水にならないように多量の海水を飲み、排尿量を減らす。窒素を含んだ物質は、最終的にアンモニア、尿素、トリメチルアミン・オキサイドなどになって排出される。アンモニアは硬骨魚類に多くて、80%以上がえらから出され、尿中に排出される量は20%以下である。尿素は軟骨魚類で多く産出され、血液中にかなり高濃度に含まれる。軟骨魚類ではえらや腎臓から尿素が失われないようにし、これで血液の浸透圧を高めている。トリメチルアミン・オキサイドは海水魚とくに軟骨魚類に多く、血液の浸透圧を高めるのに役だっている。また、脳下垂体前葉部から分泌されるホルモンのプロラクチンや副腎皮質から分泌されるコルチゾールは浸透圧調節に重要な役割を果たしていることがわかってきている。
[落合 明・尼岡邦夫 2015年3月19日]
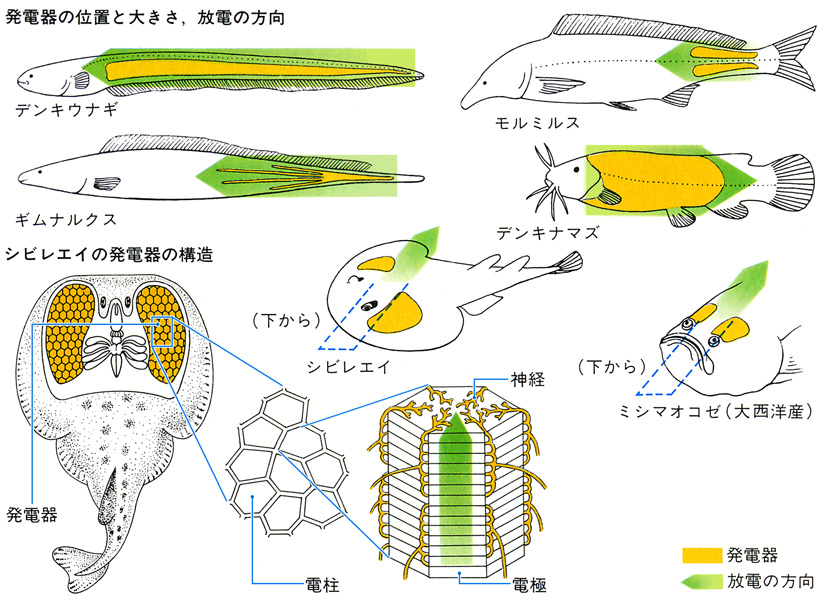
生物電気をおこして、これで周囲を探索したり、餌の動物や外敵を感電させたりする発電器をもつ魚がある。この発電器は、ある部位の筋肉から変化したものである。発電器の位置や形態は種によって異なる。基本的には、コロイドを満たした電函(でんかん)の中に発電能力のある薄い1枚の電板があり、この箱が多数重なって電気柱をつくり、多くの電気柱の集合体で発電器がつくられる。すでに数億年も昔に発電する魚が出現していたが、現在ではシビレエイ、デンキナマズ、デンキウナギなど10科に属するデンキウオ(電気魚)が知られている。シビレエイでは70~80ボルト、デンキナマズでは400~450ボルト、もっとも強い電気を出すことができるデンキウナギでは300~866ボルトで、平均350ボルトの電気によって小形の餌物を感電死させる。
[落合 明・尼岡邦夫 2015年3月19日]
餌物を集めたり、外敵から身を守ったり、仲間や異性をみつけたりするために発光する魚がある。発光器の細胞内でおこる化学反応によって光る自力発光型と、共生する発光バクテリアの発光で光る共生発光型がある。自力発光器は、神経作用で明滅する発光細胞が体表または体内の定まった位置にある。発光器には発光細胞のほか、レンズ、反射層、色素層などがあって、光を効果的に外方に向ける仕組みになっている。ハダカイワシ科、ヨコエソ科、ムネエソ科などの深海魚やカスミザメ属、カラスザメ属などの深海ザメがこの型の発光器をもつ。共生発光器は、口、食道、直腸、肛門付近にある。体表または消化管に開く1本の小管があり、海水中の発光細菌はこの小管から発光器の中へ入り込む。ヒイラギ類では食道の周りに、ソコダラ類では腹中線上、チョウチンアンコウ類ではエスカ(擬餌(ぎじ)状体)の中にこの型の発光器がある。発光魚は深海で生活するものに多い。照明により餌物を集めたり、仲間または異性を認知したり、発光によって自分の体の輪郭を消して外敵から見えなくしたりする。種類によっては発光液を体外に出して姿をくらませる。発光器の形態や配列様式は種によって異なり、重要な分類形質になる。
[落合 明・尼岡邦夫 2015年3月19日]
ある種の器官をすり合わせたり振動させたりして特有な音を出し、縄張り(テリトリー)を知らせたり、異性間の交信または外敵を威嚇したりする魚がかなり多い。カワハギ類やフグ類などは上下両顎の歯をすり合わせて鋭い摩擦音を出す。うきぶくろには特別な筋肉が付着し、その収縮や伸長でリズミカルな振動音を出すうえ、うきぶくろは音を増幅させる作用がある。うきぶくろの内側に筋肉を有する型にはマトウダイ類などが、外側に強い筋肉をもつものにハタンポ類、ニベ類、タラ類などがある。ホウボウ類には両型をもつ種がいて、交信と威嚇の2種類の音を発することができる。
[落合 明・尼岡邦夫 2015年3月19日]
毒魚には毒のある棘で刺して毒作用をおこさせるもの、皮膚から水中に毒を分泌して防御するもの、筋肉や臓器に毒があり、食べることで中毒をおこさせるものがある。ゴンズイ、オコゼ類、アイゴ類、アカエイ類などには、ひれの棘の周りに毒腺があり、これに刺されると毒液が流入されて激しく痛む。これらの魚には棘の基部に毒液をつくる毒腺がある。ヌノサラシ、コバンハゼ、ミナミウシノシタ、ハコフグなどは皮膚の下にグラミスチンとよばれる毒またはそれに類似した毒物質を生産する細胞があり、皮膚にある多数の孔から分泌する。フグの毒はテトロドトキシンとよばれ、魚類のもつ毒のなかではもっとも毒性が強い。トラフグ、マフグ、ヒガンフグなどの卵巣や肝臓、皮膚に多く、ドクサバフグには筋肉にも含まれる。しかし、養殖のフグでは肝臓でも毒性が低い。フグ類は驚いたときに体を丸く膨らませるが、このときに周囲の海水中へ、テトロドトキシンを出す。琉球(りゅうきゅう)諸島にいるツムギハゼにもテトロドトキシンがあり、とくに皮膚に多く含まれる。サンゴ礁などにいるカマス、アジ、ハタ、ブダイなどの種類の筋肉や内臓を食べると中毒をおこして死亡することがある。これをシガトキシンという。この毒はこれらの魚が食べる餌に由来する。
[落合 明・尼岡邦夫 2015年3月19日]
魚類は、脊椎動物の総種数の半数よりわずかに多く、約2万8000種の有効種が知られ、熱帯から北極または南極まで広く分布し、また、チベットの海抜5200メートル以上の高度からプエルトリコ海溝の水深8370メートルの超深海まで生息している。河川や湖沼には全体の43%に相当する約1万2000種がすんでいる。とくにアジア、アフリカ、中央アメリカ、南アメリカなどの熱帯域はコイ科、ハゼ科、カワスズメ科などの淡水魚が豊富である。いずれの大陸とも高緯度には種類が少なく、北極にはイワナ、カワカマス、サッカー、サケスズキ、南極にはある種のノトセニア類がいるだけである。また、水温は-2.34℃の氷床下から42.5℃の高温のソーダ湖にまで生息がみられる。
海では沿岸域にもっとも多く、約9000種が生息する。太平洋西部からインド洋にわたる熱帯の沿岸域は、環境が安定しているため世界でもっとも種類が多い。とくにサンゴ礁域は周年にわたって水温が高く、複雑な地形が魚類に格好の生息場所となり、チョウチョウウオ科、スズメダイ科、ベラ科、ハタ科などに多くの種がみられる。沿岸魚のなかにはイワシ類、アジ類、サバ類、ブリ類、タイ類など水産業上重要な魚が多い。水深200メートルより沖の外洋は、塩分が高く、これに適応したウバザメ、オナガザメ、カツオ・マグロ類、マンボウなど250種余りがいる。各種はそれぞれ小さいプランクトンから大きい動物までの各サイズの餌を利用している。外洋の水深200メートルから数千メートルのさまざまな深度(中深層、漸深層、深海層、超深海層)の深海には、ツノザメ類、アンコウ類、ハダカイワシ類、タラ類、キンメダイ類、イタチウオ類などがすむ。このような魚類の地理的または垂直的な分布は、水温や塩分のほか、地形、海流、水圧などに支配される。
魚類の多くは、餌場、産卵場、越冬場などを一定の順序で季節的に移動する。これを回遊migrationといい、その規模や方向は種類によって異なる。回遊には幼期回遊、索餌回遊(さくじかいゆう)、越冬回遊、産卵回遊、降河回遊、遡河回遊、両側(りょうそく)回遊、季節回遊などがある。イワシやマアジ、マサバなどは、沿岸を流れる海流または支流内を南北回遊する。高温期に高緯度海域へ餌を追って移動し、索餌回遊をする。降温期には低緯度へ向かって越冬回遊または産卵回遊をする。外洋の表層にいるサンマやカツオ・マグロ類などは回遊の規模が大きく、二つ以上の海流を横切って東西または南北方向に回遊する。とくに、クロマグロやビンナガの若魚や未成魚は、日本の周辺から北アメリカ西海岸の間を2回以上も横断し、成魚になると南の海域へ南下して産卵する。
一生の間に淡水と海との両方を回遊(両側回遊amphidromous migration)して、成育し産卵するものがある。ボラ、スズキ、ナガエバ、シマイサキなどは、幼生期に海から川へ入る。これと反対にアユやある種のハゼ類は、幼生期に海へ入って成育し、稚魚期から川へ帰って成長し成熟する。ウナギは降河(降下)回遊catadromous migrationといって、一生の大部分を淡水で生活し、成熟すると川を下って外海の中層で産卵する。ワカサギ、シラウオ、マルタ、ある種のヤツメウナギ類、チョウザメ類、サケ・マス類などは遡河(遡上)回遊anadromous migrationといって、海で成長し、成熟前に川を遡上して産卵する。サケ・マス類では降河した川に帰る母川回帰性がある。この習性は、降河した川の水のにおいをよく記憶すること、太陽の位置と体内時計により方向を知る太陽コンパスの能力があるためと考えられている。
[落合 明・尼岡邦夫 2015年3月19日]
水界での一次生物生産者は水生植物であり、海では植物プランクトンと藻類である。これらは、プランクトンや藻類を主食とする魚類によって消費される。プランクトン捕食魚や藻食魚は小形の肉食魚に、小形の肉食魚は大形の肉食魚に捕食される。このような食物連鎖の実態は、水域の環境によって著しく異なるが、一つの段階を経るごとに魚類の生産効率は10%余り低下する。一般的に一次生産量は、水温や塩分などが異なった二つの海流の接触域や垂直混合の多い水域で高い。そのような水域は太平洋や大西洋の北部、南半球の西風海流、ペルー海流やベンゲラ海流域などである。魚類生産が豊かな海域は、北半球の温暖域から冷温域の岸より300~500キロメートル内の沿岸域である。
好漁場は、北海、ニューファンドランドを中心とした大西洋の西部、日本近海から北洋、南アメリカ西岸などに形成される。北アメリカやヨーロッパの西岸域、オーストラリアの東岸なども漁場としてよく開発されている。世界における魚類の総漁獲量は1948年に1700万トン、1967年に5500万トン、1970年代では6500万トン、1985年には8600万トン、1989年には1億トンを超えるまで年々増加した。1991年に9700万トン、1997年には9000万トンを切ったが、2000年代では9000万トン前後を維持している(国連食糧農業機関『世界漁業・養殖業白書(日本語要約版)』による)。日本の漁獲量も1960年(昭和35)は450万トンであったが、遠洋漁業や沖合漁業の伸びによって1972年には1000万トンの大台にのった。200海里時代に入ってからも、遠洋漁業の減少を沖合漁業の拡大によって取り返し、しばらくはこの大台を維持していた。しかし、1990年代に入ってからは沖合漁業の漁獲量の低下により減少傾向にあり、1991年(平成3)に957万トン、1997年には598万トンと下がり、2000年代に入ってからは500万トン台を維持していたが、2009年(平成21)以降は400万トン台に、そして2011年には東日本大震災の影響で400万トンを切るまでに落ち込んだ。しかし2012年には473万トンに回復している(農林水産省「漁業・養殖業生産統計」による)。
養魚の歴史は古いが、コイのほかに、アジア、アフリカなどの熱帯域ではティラピア、サバヒー、ボラ、日本ではアユ、ウナギ、アメリカではナマズ、欧米各地ではマス類の養殖が盛んである。浅海魚の養殖の歴史は浅いが、日本では1960年ごろからブリ(ハマチ)養殖が本格化してから、マダイ、ヒラメ、トラフグ、マアジ、シマアジなどの海面における生け簀養殖(すようしょく)が盛んとなり、生産量が増加した。内水面を含めると1980年では約109万トン、1988年に143万トンとピークに達したが、1990年では137万トン、2000年では129万トン、2005年には125万トン、2010年には115万トンと下降を続けた。2011年には東日本大震災の影響で90.7万トン(岩手県、宮城県、福島県を含まない)、2012年には107万トンに低下し、2013年では103万トンにとどまっている。しかしこれは2013年の漁業の総生産量(479万トン)の21.5%を占めている(農林水産省「漁業・養殖業生産統計」による)。クロマグロはいままで幼魚を捕獲して養殖していたが、2002年に人工孵化からの完全養殖に成功した。その後、専用の配合飼料が開発され、2014年には80トンを生産するなど、産業化が進行中である。
[落合 明・尼岡邦夫 2015年3月19日]
魚類は観賞や遊漁の対象として、また動物性タンパク源として古くから各地で利用された。観賞魚としてキンギョやニシキゴイ(イロゴイ)、熱帯魚がある。
遊漁は健全なスポーツとして、年々盛んになり、遊漁の人口は日本だけでも2000万人に達するといわれている。しかし、その種類や規模が年々拡大する傾向にあるため、環境汚染や漁業者との間でしばしば紛糾しており、とくに遊漁者の資源保護や環境保全に対する自発的な協力が切望されている。
魚肉の動物性タンパク源としての価値は年々重要さを増し、畜肉の不足を補うほかに、栄養的見地から保健食料としても見直されている。魚肉や卵巣などは生鮮で利用されるほか、冷凍・塩蔵品、乾製品、練り製品、薫製品、缶詰などに加工される。
嗜好(しこう)や流通上の便利さから加工品が増加している。とくに練り製品は海外では健康食品として人気がある。塩蔵品としては塩ざけ、塩さば、塩いか、すじこ、イクラ、からすみなど、乾製品としては身欠きにしん、数の子、田作り、かつお節、そうだ節、棒だら、くさやの干物などが代表的であり、古くからそれぞれの地方名産として伝えられている。缶詰としてサケ、カニ、サバなどの水煮、マグロ、イワシなどの油漬け、サバ、イワシ、サンマ、貝類などの大和(やまと)煮、サンマ、ニシンなどのトマトソース漬けなどがある。練り製品は、古くから白身の魚によってつくられ、かまぼこ、ちくわ、はんぺんがおもであった。なお、中国では古来、淡水魚でつくられている。1960年代にスケトウダラを原料とした冷凍すり身が開発され、練り製品が大量生産されるようになった。しかし現在は国内の原料の価格の上昇によってほとんどは海外から輸入されている。また、練り製品はイワシ、サバ、グルクマなど赤身の魚から製造する技術も開発された。1955年ごろからカツオ・マグロ類、カジキ類などの赤身魚肉のフィッシュソーセージなどが製造されだし、その量は急増している。他方、魚油、魚膠(ぎょこう)、フィッシュミール(魚粉)、フィッシュソリュブル、アイシングラス、肝油などがつくられている。
[落合 明・尼岡邦夫 2015年3月19日]
人類にとって魚類は、陸上動物と並ぶ貴重な動物性タンパク源であり、沿海、湖沼、河川の流域地方においては、早くから採集狩猟と並行して漁労活動が大きな比重を占めてきた。魚類は野生動物に比べて、生息環境が水中に限定されていて比較的捕獲が容易なこと、また、回遊と産卵の季節的周期と場所に変動が少ないため、一般の採集狩猟民に比べると漁労民は定住性が高い。生食のほか焼いたり煮たりする調理法が一般的であるが、調理保存法としては乾燥、薫製、日本のなれずしのように塩を用いた発酵貯蔵やさらに発酵させて調味料として用いる魚醤(ぎょしょう)の類も東アジアから東南アジアに広くみられる。肉ばかりでなく、皮や骨、ひれ、卵、内臓も食用としてさまざまに利用されているほか、極北の民族の間では皮をなめして衣類や小物入れなどに用いたり、また魚の骨や歯を骨角器や道具に用いたりする例も多かった。そのほか、魚粉として畑の肥料や家畜の飼料として用いられたり、金魚や熱帯魚のように愛玩(あいがん)用にも親しまれたりしている。
魚類への依存度や文化的な意味づけ、関心は、社会によってさまざまであるが、なかでも大群をなして季節的に回遊する特定の魚に深く依存している社会は、魚と人間との調和のとれた共存関係がその世界観に反映されており、神話や儀礼などの主要なテーマとなっている。たとえば、北アメリカの北西海岸先住民の諸部族においては、サケの大群は、海のかなたに住む人々がサケに姿を変え、毎年きまった季節に幸をもたらすため訪れると考えられており、そこでは毎年、いちばん最初に川をさかのぼってきたサケを特別に手厚くもてなして、再会を祝うとともに、豊漁を祈る「初鮭の儀礼(はつさけのぎれい)」が行われる。北シベリアの河川流域に住むエベンキ人でも、サケ類は、祖先の霊魂がふたたびこの世に生まれ変わるため、川の源にある「魂の池」からサケに姿を変えて戻ってきたものとみなす。かつては日本でも東北地方を中心にサケにまつわる伝承や儀礼が広くみられ、またサメなどの特定の魚が漁民に豊漁をもたらすとされる信仰もみられた。あるいは特定の魚が海をつかさどる竜王の使いとみなされたり、吉祥(きちじょう)とみなされたりする。また魚の霊を慰めるために魚霊塔を建てることも広くみられた。このような神話、信仰、儀礼により特別な意味づけをされた魚は、単なる食料としてではなく、人間と共存しあう伴侶(はんりょ)として位置づけられており、その利用や消費にも一定の秩序がみられる。しかし、今日では、流通手段や加工保存技術の発達とともに魚を単なる食糧資源として収奪の対象とみなす経済学的、技術論的傾向が著しく、このような文化的意味づけは急速に失われつつある。
[伊藤亜人]
『阿部宗明著『原色魚類検索図鑑』(1963・北隆館)』▽『宮地伝三郎・川那部浩哉・水野信彦著『原色日本淡水魚類図鑑』(1976・保育社)』▽『田村保編『魚類生理学概論』(1977・恒星社厚生閣)』▽『松原喜代松・落合明・岩井保著『魚類学』上下(1979・恒星社厚生閣)』▽『M・マイルズ著、岩井保・細谷和海訳『古生代の魚類』(1981・恒星社厚生閣)』▽『日本魚類学会編『日本産魚名大辞典』(1981・三省堂)』▽『益田一・尼岡邦夫・荒賀忠一・上野輝彌・吉野哲夫編『日本産魚類大図鑑』(1984・東海大学出版会)』▽『岩井保著『新水産学全集4 水産脊椎動物Ⅱ 魚類』(1985・恒星社厚生閣)』▽『益田一・小林安雅編『日本産魚類生態大図鑑』(1994・東海大学出版会)』▽『有元貴文・難波憲二編『魚の行動生理学と漁法』(1995・恒星社厚生閣)』▽『伊藤勝敏著『図説・魚たちの世界へ』(1999・河出書房新社)』▽『桑村哲生著『子育てする魚たち 性役割の起源を探る』(2007・海游舎)』▽『中坊徹次編『日本産魚類検索』第3版(2013・東海大学出版会)』▽『Joseph S. NelsonFishes of the World Fourth Edition(2006, John Wiley & Sons, Inc.)』
化石魚復原図(無顎上綱)
化石魚復原図(顎口上綱)

アランダスピス・プリオノトレピス

ケイロレピス・トレイリ
魚類の進化を表すひれの形質変化
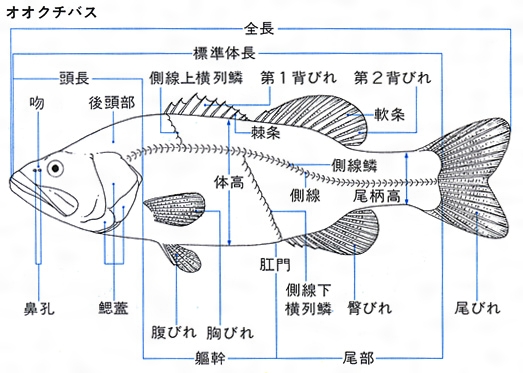
魚類の外部形質
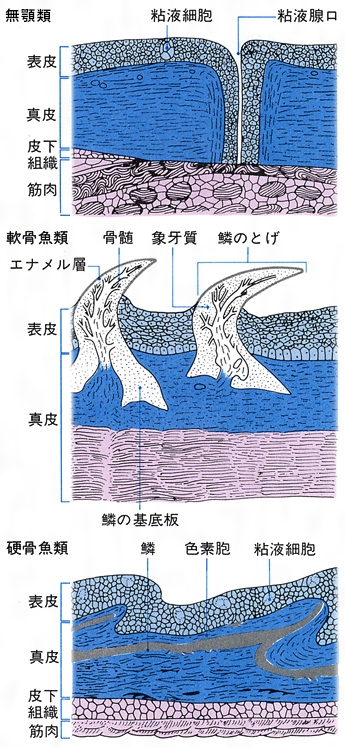
魚類の皮膚(断面模式図)
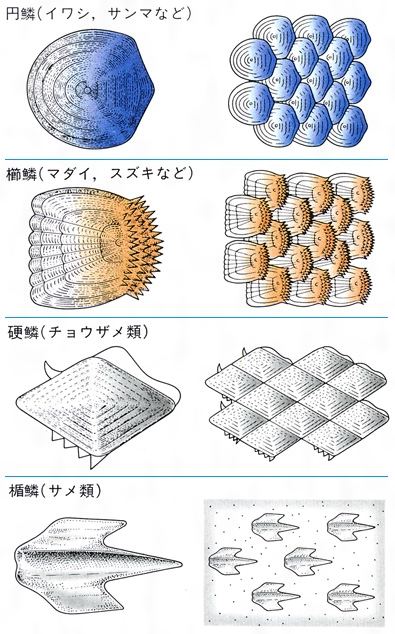
魚類の鱗のおもな種類と付き方
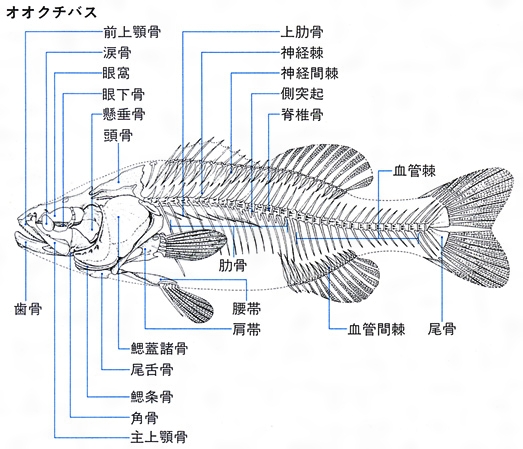
魚類の骨格
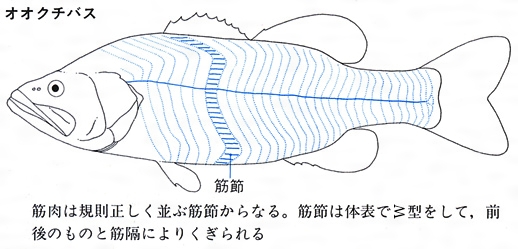
魚類の筋肉の配列
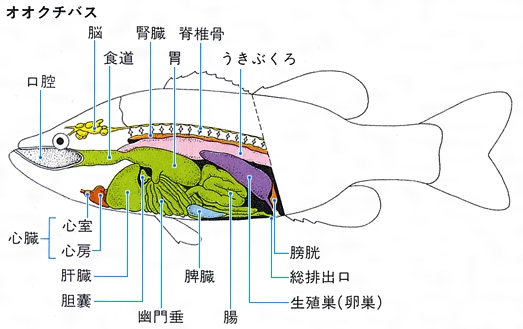
魚類の内部形質

うきぶくろの構造と働き
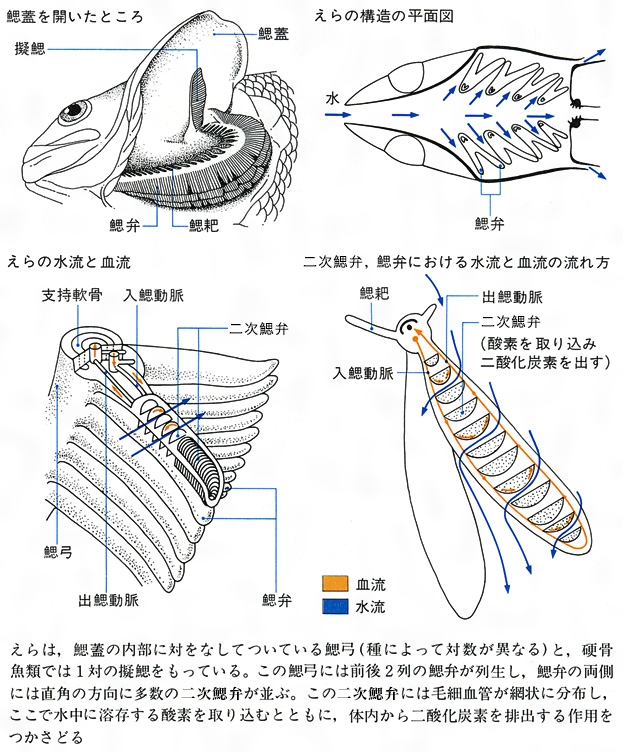
えらの構造と働き
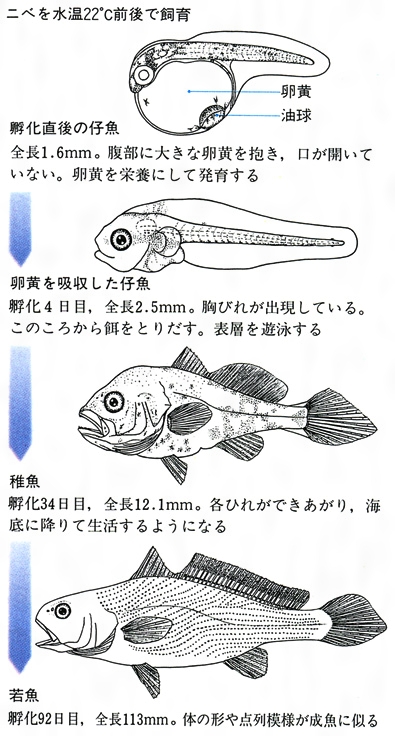
魚類の初期発育(ニベの例)

魚類の感覚器官(視覚)

魚類の感覚器官(嗅覚)
魚類の感覚器官(聴覚)
魚類の感覚器官(側線)
電気魚のおもな種類と発電器

タナゴの産卵(生態)

サザナミヤッコの幼魚(生態)

サザナミヤッコの成魚(生態)

ホンソメワケベラとクエの共生(生態)

ウツボの捕食(生態)

カサゴの捕食(生態)

カワハギの索餌(生態)

アナゴの砂潜り(生態)

ベラの休息(生態)

ムツゴロウの跳躍(生態)

ホウボウの歩行(生態)

ヘコアユの擬態(生態)
出典 小学館 日本大百科全書(ニッポニカ)日本大百科全書(ニッポニカ)について 情報 | 凡例
Sponsored by ![]()
魚類は古生代の前期に最初の脊椎動物として地球に姿を現した動物群で,現在では種類数も多く,生息範囲も地球表面積の71%を占める海洋から陸水域にわたりすこぶる広い。古代エジプトの遺跡壁面には魚が頻繁に描かれ,また養魚法の古典《養魚経》は春秋時代に陶朱公(越の范蠡(はんれい))によって著されたと伝えられている。海に囲まれ水に恵まれた日本でも魚を食用としてきた歴史が古いことは,いたるところの貝塚の発掘品からも,また《風土記》《延喜式》など古文書の記録からも明らかである。これらのことから,魚と人間とのかかわりあいがいかに古くからあったかがうかがわれる。
近年,いずれの国も魚類の食糧資源としての重要性を改めて認識し,漁業資源の科学的な国際管理の必要性を痛感するに至って,さまざまな水域について国際漁業条約が締結されるようになった。長い年月をかけて論議が続きほぼ合意に達した国際海洋法の趣意にも,海洋のスペース利用,海底資源の開発などと並び将来の食糧不足に備えて漁業資源を確保したいとの切実な願いがこめられている。また,生物学の面では,脊椎動物の最下位にある魚類が系統学的な観点から注目されるようになり,生態学,生理学などさまざまな立場からの研究が活発になってきた。なお,民俗については〈魚(さかな)〉の項目を参照されたい。
執筆者:日比谷 京
魚類は水中で生活し,主としてえらで呼吸を行い,ひれをもっている脊椎動物である。分類学的には脊椎動物門を魚類上綱と四肢動物上綱に分け,魚類上綱Piscesはさらに無顎(むがく)綱(無顎類)Agnatha,棘魚(きよくぎよ)綱(棘魚類)Acanthodii,板皮(ばんぴ)綱(板皮類)Placodermi,軟骨魚綱(軟骨魚類)Chondrichthyes,硬骨魚綱(硬骨魚類)Osteichthyesに分けることが多い。
地球の歴史において,もっとも古い魚類の化石はカンブリア紀後期の地層から発見されている。北アメリカ大陸のワイオミング州の5億1千万年前のアナトレピスAnatolepisがそれである。無顎綱の1種で,上下両あごがなく,現生のヤツメウナギなどを含む円口類の祖先にあたる。無顎類はシルル紀,デボン紀に栄え,石炭紀の終りまでにほとんど絶滅してしまった。無顎類のえらを支える骨のうち,前方のものが変化してあごの骨となり,有顎の棘魚綱がシルル紀に出現する。棘魚綱はひれに強いとげをもっており,板皮綱,軟骨魚綱,硬骨魚綱の共通の祖先に近いと考えられる。研究者によっては棘魚綱を硬骨魚綱の祖先とし,板皮綱を軟骨魚綱の祖先と考えて魚類の分類体系を考える場合もある。板皮綱はシルル紀に出現し,デボン紀から石炭紀にかけて繁栄したが,石炭紀の初めに絶滅した一群の魚類である。頭部と胸部は固い骨質の装甲に覆われ,あごにはほんとうの歯がなかった。体長数mに達するほど大きくなったことがわかっている。
軟骨魚綱はデボン紀に出現し,多様化して現在に至っている。骨格系が軟骨でできているのが特徴である。うろこは小さな楯鱗(じゆんりん)で,うろこが変化してできた歯をもっている。軟骨魚綱は大きく板鰓(ばんさい)亜綱(板鰓類)と全頭亜綱(全頭類)に分けられる。板鰓亜綱にはサメやエイの類が含まれ,全頭亜綱にはギンザメの類が含まれる。板鰓類の歯は歯冠が硬いエナメル質で覆われているが,全頭類の歯にエナメル質はないことがわかっている。軟骨魚類は体内受精を行い,雄は腹びれが変化した交尾器を備えている。この交尾器は軟骨魚類の祖先といわれる古生代の板皮類の化石にも発見されている。現在,地球上に生息している軟骨魚類の大部分を占める板鰓類のうち,もっとも原始的なものは鰓裂(さいれつ)が6対ないし7対あるカグラザメ目とラブカ目である。両者とも深海に適応して生きのびているが,中新世の時代までは比較的浅い海にも生息していたことがわかっている。カグラザメ目は2属,ラブカ目は1属1種で,いずれも世界中に広く分布している。ラブカは口が頭部の先端に開口している。歯は3本のとげのような突起からなる。ネコザメ目は鰓裂が5対のサメで,中生代に栄えたヒボーダス類の直接の祖先と考える説と,そうでなく現代的なサメ類の分岐した一群であると考える説がある。前方の歯は3尖頭(せんとう),または1尖頭であるが,あごの両側の側歯は頂上が丸い臼歯(きゆうし)状になっており,サザエなど硬い殻をもった貝を砕いて食べる。ネズミザメ目は種類も多く,次のツノザメ目,エイ目とともに現在もっとも繁栄しているサメ類である。ネズミザメ科のホオジロザメ,アオザメ,メジロザメ科のイタチザメなどは歯が大きく鋭いうえにどうもうで,人間を襲った事例も多く恐れられている。ツノザメ目は背びれにとげがあり歯は小さい。深海性のツラナガコビトザメのように20cmほどで成熟する小さなサメもこの目に含まれている。エイ目は胸びれが拡大して大きな体盤を形成し,これを上下に動かして水中を泳ぐ。歯は敷石状で貝類などを好んで食べる。鰓裂はふつう5対で体の腹面に開く。6対の鰓裂をもつものは最近発見された2種のみである。
硬骨魚綱はシルル紀の後期に出現し,種類も多く現在もっとも繁栄している魚類である。骨格系は主として硬骨からできているが,原始的なものでは軟骨が多く,また高等なものでも二次的に軟骨部分の多いものもある。硬骨魚綱はさらに条鰭(じようき)亜綱(条鰭類),総鰭亜綱(総鰭類),肺魚亜綱に分けられる。条鰭類は現生魚類の95%以上を占めるほどになり繁栄しているが,すでに古生代のシルル紀に出現している。原始的なものでは外骨格が発達し,内骨格は軟骨が多かったが,高等なものほど内骨格ががんじょうな硬骨となり,運動機能と口部の骨格が時代とともに効率がよくなる方向に進化してきた。総鰭類はシーラカンスやエウステノプテロンを含み,肺魚類とともに両生類の直接の祖先として議論の的となっている。現生の硬骨魚綱のうちもっとも繁栄している条鰭亜綱は,原始的な軟質上目(チョウザメ類)から全骨上目(アミアやガーパイク)を経て真骨上目(ニシンなど魚類の90%以上を占める)へと進化した。進化の傾向として内部骨格の化骨による強化と,外部を覆ううろこや骨板の軽装化があげられる。真骨上目の分類体系は最近急激な変化を遂げていて定説なるものがないが,進化の程度により,低位群,中位群,高位群に分けられる。原始的な低位群は大きくオステオグロッサム目,ニシン目,カライワシ目,ウナギ目,ネズミギス目,コイ目,サケ目に分けられる。ウナギ目はレプトセファルス幼期をもつということでカライワシ目と近縁と考えられている。サケ目から中位群のハダカイワシ目が進化し,さらにタラ目,アンコウ目,メダカ目,ダツ目,フリソデウオ目,キンメダイ目,マトウダイ目などが生じた。このうちキンメダイ目のような魚類から上位群のスズキ目が進化し,さらにスズキ目からカサゴ目,カレイ目,フグ目,タウナギ目などが分岐して生じたと考えられている。このような真骨上目の進化は摂餌機構と運動機能の効率を高める方向で進むと同時に,表層生活,中層遊泳,底生生活,深海・急流・湖水での生活,などいろいろな生活環境への適応現象によって多様化している。より進化の進んだ上位群の魚では,主上顎骨は口の縁辺から退き歯がなくなる。前上顎骨が長く大きくなり,この骨に上あごの歯が生え,口は伸出できるようになる。腹びれは体の中央下の位置から前方に移動して胸びれの近くに移り,胸びれと連動し,すばやく行動するようになる。背びれ,しりびれ,胸びれ,腹びれに棘条が生じ,ひれを強化する。また骨格は癒合して,進化の進んだものほど骨の種類の数が少なくなる。眼窩蝶形骨(がんかちようけいこつ),中烏口骨(ちゆううこうこつ)などが失われ,肉間骨は少なくなり,タチウオなどを除いて一般に脊椎骨数も減少する傾向が見られる。
魚類の体はえらぶたの後端と肛門位を境にして,頭部,体幹部,尾部の3部に分けられる。基本的な体型は,水中での運動に適した紡錘形で,マグロ,カツオなど外洋性回遊魚の形はその典型的な例である。遊泳力の劣る沿岸域,淡水域の魚や底生性の魚には側扁形(マダイ,チョウチョウウオ,ヒラメ),縦扁形(アカエイ,アンコウ),延長形(ヤツメウナギ,ウナギ,アカヤガラ)などの体型をもつものが多い。
皮膚は表皮と真皮の2層からなる。表皮には無数の粘液細胞があり,つねに粘液を分泌して体表を保護している。多くの魚の皮膚にうろこがあるが,円口類,シビレエイ,タチウオなどにはない。うろこには楯鱗,硬鱗,骨鱗などの種類があり,それぞれ構造が異なる。楯鱗はサメ・エイ類のうろこで,厚い象牙質の外側が硬いエナメル層で包まれ,内側には神経や血管の入りこんだ髄があって,歯と同じ構造をしている。チョウザメ類の硬鱗は厚い板骨層の表面に真珠光沢を呈するガノイン層があり,敷石状に並ぶ。硬骨魚類の骨鱗はコラーゲン質の繊維薄板が多数重なった表面を骨質層が覆ったもので,すこぶる強靱であり,真皮内に屋根瓦状に配列している。その遊離縁に櫛(くし)の歯状の突起のあるものを櫛鱗(しつりん)/(せつりん),ないものを円鱗と呼ぶ。骨鱗の表面には成長に伴って隆起線ができるが,冬など成長の衰えたときにできた隆起線の間隔は狭いので,これによって魚の年齢を推定することができる。骨鱗の周辺および真皮層の深部には黒いメラニン,黄または赤いカロチノイドを含む色素細胞やグアニン結晶を含む虹胞(こうほう)が分布し,それらのさまざまな組合せによって魚種特有の体色,斑紋が示される。
ひれには陸生脊椎動物の前肢に当たる1対の胸びれ,後肢に当たる1対の腹びれおよび皮膚のしわとして生じた背びれ,しりびれ,尾びれがある。ウナギ,フグには腹びれがなく,ウツボには胸びれも腹びれもない。多くのサメ類,アジ,ハゼなどには背びれが2基,タラには背びれが3基,しりびれが2基ある。また,サケ,アユには背びれの後方に脂びれ,サンマ,マグロ,サワラなどには背びれとしりびれの後方に何個かの小離鰭(しようりき)がある。コバンザメの第1背びれが吸盤状になり,ホウボウの胸びれの下部が歩脚肢状になっているなど,ひれが変形して特別な機能を果たしている例も少なくない。
骨格系は,頭骨と脊椎骨とからなる中軸骨格と,肩帯,腰帯などひれを支持する骨よりなる付属骨格とに分けられる。円口類には軟骨性の中軸骨格だけしかなく,板鰓類の骨格も軟骨性である。硬骨魚類の骨格は一部の軟骨を除いて骨化が進んでおり,進化した魚ほどその程度は著しい傾向がある。体側の筋肉は前後に並んだ多数の筋節からなり,原始的な体節構造を示す。筋節の数は魚種によって異なる。体側筋の左右表層には血赤色の表層血合筋(ちあいきん)が縦走し,さらに外洋性回遊魚には脊椎骨の左右両側に接して真正血合筋が縦走している。頭部には眼球,下あご,えらぶたなどを動かす筋肉,体部にはそれぞれのひれを動かす筋肉がある。筋肉の色は白色から赤色まで魚種によりさまざまであるが,遊泳力の優れた魚では筋肉色素(ミオグロビン)が多いため赤みが強い。ただし,サケ類の筋肉の赤色はカロチノイドの存在による。
口には一般に上下両あごがあるが,円口類では両あごを欠き,口は他の動物に吸着し肉や血液を吸いとるのに適応して吸盤状をなし,歯も角質で発達していない。多くの魚は上下両あごに円錐歯を備えるが,同じ円錐でも遊泳力の優れたカツオ,サワラでは小さく,遊泳力の劣ったアンコウ,タチウオなどでは大きく,数も多い。フグでは物をかみ切るのに適した門歯状を呈し,硬い殻をもった貝や甲殻類を食べるイシダイ,マダイなどでは臼歯が発達し,ネコザメ,ホシザメなどは敷石状の歯をもつ。口蓋部に歯をもつものも多いが,コイ科の魚では口腔の奥に咽頭(いんとう)歯があり,餌のそしゃくに役だっている。また,ウバザメ,イワシ,コノシロ,サンマ,ボラなど水中のプランクトンやデトリタスをこしとって食べる魚では,鰓弓(さいきゆう)の内側に列生した突起(鰓耙(さいは))が発達する。
魚には唾液腺はないが,口腔粘膜には粘液細胞が多い。胃は基本的には噴門部,盲囊部,幽門部の3部に分かれるが,単純な構造の胃をもつものも多く,コイのように胃のないものもある。コノシロ,ボラの幽門部は平滑筋層が肥厚して硬く,そろばん玉と呼ばれる。胃では塩酸とペプシノーゲンとが分泌され,タンパク質の予備消化が行われる。腸の始部には魚類特有の房状の幽門垂をもつものが多い。腸の長さ,巻き方は魚種によって特徴がある。板鰓類,肺魚類,チョウザメ類では腸の後部にらせん弁があり,腸の内腔がらせん階段状をなして吸収面積を広くしている。肝臓は胃に接して位置する大きな腺で,膵臓は板鰓類では独立した器官であるが,円口類,硬骨魚類では散在性で,コイ,クロダイ,ヒラメなどでは膵臓組織が肝臓内に侵入して肝膵臓をなしている。肝臓からは胆汁を,幽門垂,膵臓,腸腺からは各種の消化酵素を分泌しているが,それらの活性は魚の食性や発育段階によっても異なる。栄養の吸収は幽門垂,腸で行われる。
魚類のおもな呼吸器官はえらである。円口類では6~15対のふくろ状の鰓囊(さいのう)をなす。ヤツメウナギでは7対の鰓囊がそれぞれ体側に外鰓孔(がいさいこう)を開くので一見8対の眼があるように見える。板鰓類は4.5~6.5対のえらをもち体側に各5~7個の外鰓孔を開き,硬骨魚類は4対のえらをもち左右各1個の外鰓孔を開く。えらは軟骨,または骨質の鰓弓に2列の鰓弁が並んだもので,鰓弁には微細な二次鰓弁(鰓薄板)が櫛の歯状に並んでいる。その薄板には無数の毛細血管が平行に走っているので,口から吸いこまれた水がその表面を流れる間に,水と血液との間でガス交換が行われる。水の流れに対して血液の流れが逆の方向になっているので,いわゆる対向流系をなし,ガス交換の能率はきわめてよい。呼吸数は1分間のえらぶたの開閉数で測られるが,魚種,体の大きさ,水中溶存酸素量,魚の運動量などによって変動する。えらのほか,肺魚はうきぶくろ,トウギョなどは口腔内の迷器,ドジョウは腸,ウナギなどは皮膚でも呼吸を行う。
心臓は1心房1心室よりなり,自律性の拍動を示すが,迷走神経による抑制的支配を受ける。心臓から拍出された血液は動脈球(板鰓類では心臓球)から腹大動脈を経てただちにえらに入る。ここでガス交換を行って動脈血となると頸動脈(けいどうみやく)を経て頭部の,また背大動脈を経て体幹部,尾部の末梢血管に向けて流れる。毛細血管から静脈に流入した血液はそれぞれ腎臓,肝臓などを通って前主静脈,後主静脈,肝静脈から静脈洞に合流して心房に戻る。血液総量は体重100g当り5~7ml。赤血球は有核,楕円形でその数は血液1ml中に(0.15~3.9)×106個,血色素(ヘモグロビン)量は血液100ml中に2~21g。
体内でタンパク質,核酸などの分解によってできたアンモニアは毒性が強いので,高等脊椎動物では尿素に変えられて尿中に出されるが,魚類ではアンモニアの大部分はそのままえらから水中に排出される。含窒素性老廃物としては,そのほか尿素,トリメチラミンオキサイド,クレアチン,クレアチニンなどの形でも排出される。
水中にすむ魚類にとって,体液の浸透圧をつねに一定に保つことは生命を保つうえで基本的に重要なことである。淡水魚は,体液の浸透圧が環境水のそれより高いので,えらや粘膜からの水の浸入を防ぎ,塩類の欠乏を補う機構を備えており,腎臓からは多量の低張尿を排出する。海産魚は,体液の浸透圧が周囲の水より低いので,海水を飲んで水分を取りこみ,過剰の塩類はえらの塩類細胞を通じ,また尿中にも排出する。ただし尿量は少ない。このような機構は淡水魚ではプロラクチン,海産魚ではコルチゾールその他のホルモンによって調節されている。なお,海産の板鰓類はかなりの量の尿素を体液中に保持して浸透圧を高めるために役だてている。
(1)神経 脳は終脳,間脳,中脳,後脳,髄脳の5部よりなるが,その配列はほぼ直線的で,終脳は発達せず,嗅覚(きゆうかく)だけに関係する嗅脳の段階にとどまっている。中脳は視覚の中枢をなし,背側面が高まり,視葉と呼ばれる。交感神経幹は脊柱の腹側に左右対をなして走り,副交感神経は迷走神経によって代表される。(2)視覚 眼にまぶた,涙腺はない。レンズは球形で,レンズ筋によりレンズと網膜との距離を変えて多少の遠近調節はできるが,遠方までは見えない。一般に眼は頭の両側にあるので単眼視野は広いが,両眼視野は狭い。色彩感覚は魚種により差が著しく,ボラ,コイなどは優れ,ネコザメ,クロダイなどは劣っている。間脳の背面に突出している上生体(じようせいたい)(松果体(しようかたい))も光を感ずることが知られている。(3)聴覚 外耳,中耳はなく,内耳だけがある。内耳は通囊と小囊とよりなり,前者には三半規管が付属するが,後者からは蝸牛(かぎゆう)管は発達せず,壺(こ)と呼ばれる小胞が生ずるだけである。コイなどはうきぶくろが鼓膜の役をし,内耳にその振動を伝える特別の骨伝導路(ウェーバー器官)をもつので聴覚が鋭敏である。魚類の可聴域は20~2万Hzとされているが,感度のよいのは100~1000Hzの範囲で,低い音は体側側線器で受容する。また,音源方向を知覚することもできる。(4)味覚 味蕾(みらい)は口腔,えら,ひげ,皮膚などに広く分布し,さまざまの味覚刺激を識別できることは学習実験や電気生理学的研究によって明らかにされている。(5)嗅覚 鼻孔は円口類では頭部正中線上に1個,板鰓類では頭部腹面に左右1対ある。硬骨魚では眼の前方に左右2個ずつ開き前後に並ぶが,中はトンネル状に通じて鼻腔をつくる。魚種により鼻孔が左右1個ずつのものもある。サメ,ウナギなど肉食性,あるいは夜行性の魚は一般に嗅覚が鋭く,餌をさがすのもこの感覚によることが多い。また,サケが産卵のため自分の生まれた川に帰るのは稚魚期に記憶された川水のにおいによるものとされている。(6)その他の感覚 頭部や体側の側線器官は温度変化の刺激,電気刺激,機械的刺激(水流,水圧),塩類(1価の陽イオン)の化学的刺激などを受容する。
おもに食べる餌によって,プランクトン食性,底生生物食性,魚食性,藻食性,デトリタス食性,雑食性などに分けられる。食性は魚のすんでいる場所,遊泳力などにもよるが,捕食器官とも密接な関係を示し,プランクトン食魚(マイワシ,サンマ)は遊泳時口を大きく開き,えらには水中のプランクトンをこしとるのに適した多数の細長い鰓耙を備える。底生生物食魚(マダイ,トラフグ)および藻食魚(アイゴ,ブダイ)は門歯状の歯をもち,とくに硬い殻をもつ餌を捕食する魚(イシダイ,ネコザメ)は臼歯,または敷石状の歯を備える。魚食魚(アオザメ,タチウオ,アンコウ)は鋭い歯をもち,デトリタス食魚(ボラ,タツノオトシゴ,サギフエ)は口が小さく,デトリタス(生物体の分解砕片)を吸いこむのに適した管状の口をもつものも多い。しかし,ヒイラギが夏はおもにプランクトンを,秋には底生生物を,冬にはヨコエビ類をというように,餌生物の季節による量的消長に応じて食性を変えるものもあり,また,アユが稚魚期にはプランクトン,幼魚期には水生昆虫,成魚期には付着ケイ藻,ラン藻を食べるというように発育段階により,あるいは生息場所により食性を変える場合もあり,魚の食性は必ずしも固定的なものではない。
魚類は淡水魚と海産魚とに分けられるが,なかには一生の間に川と海との間を移動するもの(カワヤツメ,サケ,アユ,ウナギ,イトヨ)もあり,また塩分変化の著しい河口の汽水域にすむ魚種も少なくない。川の上流部は急流をなし水温が低く,冷水性のイワナ,ヤマメ,カジカなど,中流部はオイカワ,アマゴなど,下流域は流れが緩やかで水温もやや高く,ウグイ,モツゴ,ナマズ,ドンコなど種類が多い。湖沼は緯度,高度により,また栄養塩類の多少により魚類の種類や量に変化が多いが,一般に湖岸部には小動物や水草を食べる魚が多く,湖心部にはプランクトン食性の魚が多い。
海は内湾と外海,外海はさらに大陸棚上の浅海とそれより沖の大洋とに分けられる。内湾や浅海は陸からの栄養塩類の補給が多く,光が海底までとどいて植物の光合成が盛んなため,餌生物が豊富で魚の種類も量も多い。とくに内湾や陸地に接した水域は海藻が繁茂して底生生物も多いので,魚のもっともよい餌場であり繁殖場でもある。大洋の魚の多くは表層ないし中層を広い範囲にわたって行動する回遊魚で,漁業上の重要魚が多い。それらのほとんどは魚食魚で,魚種それぞれに適した水温を求め,とくに餌となる魚の群れを追いながら大群をなして移動する場合が多い。また,回遊には親魚が産卵場を目ざし,仔魚が成育に適した場所を求めて行うものなどがある。マグロ,カツオなど活発な回遊魚では,平常の遊泳時には主として血合筋が用いられ,普通筋は餌をとらえるときとか敵に襲われたときとか短時間の急激な泳ぎにだけ用いられる。
浅海も大洋も暖流域と寒流域とで魚類組成は異なり,マイワシ,トビウオ,ボラ,キハダマグロ,カツオ,ブリ,ベラなどは暖流性,ニシン,サケ,ハタハタ,タラ,オヒョウなどは寒流性である。また,ガンギエイ,マエソ,ギス,ホウボウ,ヒラメ,アンコウなどの底生魚は季節により深浅回遊をするものが多い。なお,生物学的な意味での深海魚は数千mの深海にすむ魚(チョウチンアンコウなど)をいう。
発光魚は比較的深い海にすむものが多い。発光器は体の腹側にある場合が多く,発光細胞が集まっているもの(ハダカイワシ,ヨコエソ),発光物質を分泌する発光腺をもつもの(キンメモドキ,ツマグロイシモチ),発光細菌が共生しているもの(マツカサウオ,ホタルジャコ)の3型がある。シビレエイやデンキウナギなどの発電器官は体側筋で,原理的には電池を直列につないだ構造になっていて,神経刺激により放電する。電圧は数Vから数百V程度。発音する魚も少なくないが,イシモチ,ホウボウなどは筋肉の収縮でうきぶくろを振動させ,また,モツゴ,ブリは歯を,ギギはひれの棘条をきしらせて音を出す。発光と発音は同種内での相互認識に役だち,とくに生殖時にその意味が大きい。発電は外敵に対する威嚇防衛と餌となる動物を倒すために役だつ。
魚にも敵から身を守るためのさまざまな適応現象が見られる。ハコフグ,マツカサウオなどは体を硬い甲鱗で包み,ハリセンボンは体表に無数の長いとげをもつ。アカエイ,ゴンズイ,ハオコゼなどはひれの棘条の基部に毒腺を備える。また,カレイ類は海底の砂と,ヨウジウオ,イザリウオは岩礁や海藻とまぎらわしい体色を呈する。イワシ,サバなどは背面が青く,腹面が白ないし銀色で,上下いずれからも発見されにくい。ウナギ,ナマズは夜行性で昼間は物陰に潜み,カクレウオはナマコの腸内に潜り,エボシダイはカツオノエボシの毒腺をもつ触手の間に隠れるなど,同様の例は非常に多い。また,自分の体を大きく見せることは敵を威圧する効果をもつが,フグが水を吸って腹を膨らませるのはそのよい例であり,イワシ,テンジクダイなどが大群をつくるのも同様な意味をもつものといわれる。
魚の性比は,マルブナのように雄がきわめて少ないというのはごく特殊な例で,一般にはほぼ1:1である。ただ,クロダイは2年魚まではすべて雄で,その後4年魚になるまでにおよそ半数が雌に変わり,結局は性比が1:1になる。ときに性転換を示す魚としてはソードテール,ベラ類,キダイ,イネゴチ,ヨコエソなどが知られている。第二次性徴は魚の種類によってさまざまであるが,一般的に性的成熟に伴って明らかになる場合が多い。体の大きさは,ウナギ,グッピーなどでは雌が大きく,マダイ,モツゴなどでは雄が大きい。カンダイの雄は前頭部が突出し,サケの雄は上あごがかぎ鼻状に曲がる。オイカワの雄はしりびれの前部が著しく長くなり,ドジョウの雄の胸びれは先端がとがる。キュウセンの雄(俗にアオベラ)の体色は青みが強く,雌(アカベラ)は赤みが強いが,ベニザケの雌,タナゴ,ウグイなどの雄が産卵期に示す美しい体色は婚姻色という。コイ科の魚,アユ,メダカなどの雄は頭部やひれに白点状の突起を生じ,追星(おいぼし)と呼ばれる。また,タナゴ類の雌は生殖孔の部分から長い産卵管をのばす。
大部分の魚は卵生であるが,ホシザメ,アカエイ,カダヤシ,メバル,カサゴ,ウミタナゴなどは卵胎生である。卵生の場合は雌の産卵とほぼ同時に雄が放精して,水中で受精が起こる。産卵数は親魚の大きさによっても異なるが,サケ4000,カワヤツメ7万,ニシン8万,コイ50万,ヒラメ50万,マダイ600万,マアナゴ800万などの数字がある。卵は性状から浮性卵,沈性卵(サケ,マス),付着卵の3種に分けられる。浮性卵は1個ずつ分離して水に浮いているもの(イワシ,ソウギョ,アナゴ,アジ,マグロ,ヒラメ)とゼリー状の物質の中に多数の卵が包まれて浮いているもの(アンコウ,イザリウオ)とあり,付着卵には卵の表面に粘着物質をもつもの(ニシン,コイ,フグ),付着膜をもつもの(アユ,ワカサギ),付着糸をもつもの(メダカ,サンマ,トビウオ,ハゼ)などがある。また,特殊なものとしては,親の口腔の中(テンジクダイ,ティラピアなど,マウスブリーダーと呼ばれる),雄の育児囊の中(タツノオトシゴ),貝のえら(タナゴ類),親のつくった巣の中(トゲウオ)などで保護される卵もある。
浮性卵は直径1mm内外,1日程度で孵化(ふか)するものが多いが,付着卵の孵化日数はやや長く,サケの卵は直径6~7mmで,水温8℃では孵化に60日間を要する。浮性卵,付着卵から孵化したばかりの仔魚(しぎよ)は体が透明で,腹の下には卵黄囊をつけている。これを吸収しつくすころから運動も活発になり餌をとるようになる。やがて稚魚期になると体型も整い,体色,斑紋も明らかになる。この間に変態を示す魚種もあり,例えばウナギのヤナギの葉状の仔魚(レプトセファラス)が体は透き通っているが親と同形の稚魚(シラスウナギ)になる。カレイ類では頭部の両側にあった眼の一方が反対側に移動する。次いで稚魚期から幼魚期になると,体型,体色などの特徴が成魚とほぼ同様になる。それまで藻場に群れていたものがこの時期に沖合へ出ていくなど,すみ場を変えるものが多い。
無機資源と違い,生物資源は再生産を繰り返す点に特色がある。しかし,魚の産卵数は多いが,ニシンで調べられた例でも,育つ稚魚数は親魚1尾当り最高4.3尾,最低0.05尾,平均1.12尾である。このように大きな減少は害敵による食害,環境条件の悪化,病気などによる。いずれにせよ,天然での魚類の増加率は決して高いものでなく,漁業により自然増分以上をとれば資源量は急激に減少する。また,プランクトン,藻類,小動物が十分に繁殖できないような環境状態は,餌の不足によって有用魚類の資源量に影響を及ぼす。魚類資源の維持増産のためには国内はもちろん,国際的にも真剣な努力が重ねられている。
魚は種類が多くいたるところで入手も容易なため,昔から食用とされ,一般には生鮮魚として家庭で調理のうえ食膳に供される。魚肉のタンパク質,脂肪は栄養化学的にも良質であり,魚種それぞれの〈しゅん〉と呼ばれる時期にはとくに美味のものが多い。その反面,鮮度低下,腐敗が起こりやすく,品質が季節によって一定せず,大漁,不漁により供給量の変動が大きいなどの欠点もある。そこで,保存性を与え,風味を加えるため,さまざまの加工法がくふうされている。冷蔵,冷凍,乾燥,塩蔵,缶詰,薫製,練製(かまぼこ,ちくわなど),煮熟(つくだ煮など),その他がそれである。魚肉のほか,内臓(塩辛),卵巣(かずのこ,筋子,キャビア,うるか,からすみ,たらこ),ひれ(フカのひれ),軟骨(氷頭(ひず)),腎臓(めふん)なども珍重される。そのほか,ツノザメ,イワシなどの魚油は食用,化学原料用に,魚粉は家畜,養殖魚の飼料用に,魚かすは肥料用に,内臓は医薬製造用(肝油,インシュリン)になり,用途は広い。
執筆者:日比谷 京
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
Sponsored by ![]()
Sponsored by ![]()
出典 株式会社平凡社百科事典マイペディアについて 情報
Sponsored by ![]()
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
Sponsored by ![]()
fishes
脊椎動物の中で,指をもたず,鰭
執筆者:籔本 美孝
出典 平凡社「最新 地学事典」最新 地学事典について 情報
Sponsored by ![]()
…目はカメラに似たレンズ眼である。他の脊索動物は微小な浮遊生物を鰓裂でろ過して食べるろ過食であるが,あごとひれを獲得した魚類では,より大きな動物を追跡して捕食できるようになり,四足類では消化器官がとくに発達して食性が多様化した。循環系には少なくとも3室からなる心臓ができて血流を速くし,ヘモグロビンをもつ赤血球の獲得とあいまって,大量の酸素を組織に運ぶことが可能になり,行動や脳の働きが活発になった。…
…(b)軟骨または硬骨の脊柱と頭骨ができて脳を包み眼はレンズ眼で,ヘモグロビンを含んだ赤血球ができて酸素を運ぶ能率が高まるもの 〈脊椎動物門〉がこれに属する。これには次のような進化の段階を異にする現生の綱があり,これらは魚類と四足動物の2上綱に整理される。なお魚類と両生類は,胚に羊膜ができないので一括して無羊膜類と呼ばれることがあり,ふつう体外受精である。…
※「魚類」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
Sponsored by ![]()
夏の暑さに体が慣れること。数日から数十日間で起こる短期暑熱順化と、数年または数世代にかけて起こる長期暑熱順化とがある。→寒冷順化[補説]近年では、冷房設備の普及にともない短期暑熱順化が起こりにくくなっ...